Virusinės ligos atsirado senovėje, tačiau virologija kaip mokslas pradėjo vystytis XIX amžiaus pabaigoje.
1892 m. Rusų mokslininkas botanikas DI Ivanovsky, tyrinėdamas tabako lapų mozaikinę ligą, nustatė, kad šią ligą sukelia mažiausi mikroorganizmai, praeinantys per smulkiai išgręžtus bakterijų filtrus. Šie mikroorganizmai vadinami filtruojamais virusais (iš lotynų viruso - nuodai). Vėliau buvo įrodyta, kad yra kitų mikroorganizmų, praeinančių per bakterijų filtrus, todėl filtruoti virusai pradėti vadinti tiesiog virusais.
Apskritai, viriono forma ir voko buvimas ar nebuvimas mažai ką pasako apie tai, kurias ligas gali sukelti virusai ar kurias rūšis jie gali užkrėsti, tačiau tai vis tiek yra naudingas būdas pradėti virusų klasifikaciją. 5 paveikslas Virusai gali būti sudėtingos formos arba palyginti paprasti.
Kuris iš šių teiginių apie viruso struktūrą yra teisingas? Viruso šerdyje yra genomas arba bendras genetinis viruso turinys. Virusų genomai paprastai yra maži, palyginti su bakterijomis ar eukariotais, kuriuose yra tik genai, koduojantys baltymus, kurių virusas negali gauti iš ląstelės-šeimininko. Ši genetinė medžiaga gali būti viengrandė arba dviguba. Jis taip pat gali būti tiesinis arba apskritas. Nors daugumoje virusų yra vienas nukleorūgšties segmentas, kiti turi genomus, sudarytus iš kelių segmentų.
Didelį indėlį tiriant virusus įnešė sovietiniai virusologai: M.A.Morozovas, N.F.Gamaleya, L.A.Zilberis, M.P.Chumakovas, A.A.Smorodincevas, V.M.Ždanovas ir kiti.
Virusai yra neląstelinė forma gyvosios materijos egzistavimas. Jie yra labai maži. Pagal V. M. Ždanovo perkeltinę išraišką „jų dydį, palyginti su vidutinių bakterijų dydžiu, galima palyginti su pelės dydžiu dramblio atžvilgiu“. Virusus pamatyti pavyko tik išradus elektroninį mikroskopą.
Virusai gali būti laikomi privalomais viduląsteliniai parazitai... Virusas turi prisijungti prie gyvos ląstelės, ją nuryti, gaminti baltymus ir nukopijuoti genomą, rasti būdą, kaip pabėgti iš ląstelės, kad virusas galėtų užkrėsti kitas ląsteles ir galiausiai kitus žmones. Virusai gali užkrėsti tik tam tikrų rūšių šeimininkus ir tik tam tikras ląsteles toje šeimininkėje. Šio specifiškumo molekulinis pagrindas yra tas, kad ant ląstelės-šeimininkės paviršiaus turi būti randama specifinė paviršiaus molekulė, žinoma kaip viruso receptorius, kad virusas galėtų prisijungti.
Šiuo metu virusams tirti naudojama daugybė metodų: cheminių, fizikinių, molekulinių biologinių, imunobiologinių ir genetinių.
Visi virusai skirstomi į žmones, gyvūnus, vabzdžius, bakterijas ir augalus.
Virusai turi įvairiausių formų ir biologinių savybių, tačiau visi jie turi bendrų struktūrinių bruožų. Subrendusios viruso dalelės vadinamos virionais.
Be to, metaboliniai skirtumai, pastebėti skirtingų ląstelių tipuose, remiantis diferencine genų ekspresija, yra tikėtinas veiksnys, dėl kurio ląstelės gali naudoti virusą replikacijai. Ląstelė turi sudaryti virusui reikalingas medžiagas, tokias kaip fermentai, kuriuose pačiame viruso genome nėra genų arba virusas negali daugintis naudodamas šią ląstelę.
Virusas turi „perimti“ ląstelę replikacijai. Viruso replikacijos ciklas gali sukelti dramatiškus biocheminius ir struktūrinius ląstelės-šeimininko pokyčius, kurie gali pakenkti ląstelėms. Šie pokyčiai, vadinami citopatiniais efektais, gali pakeisti ląstelės funkciją ar net sunaikinti ląstelę. Kai kurios užkrėstos ląstelės, pavyzdžiui, užkrėstos peršalimo virusu, žūva lizės ar apoptozės metu, tuo pačiu metu išlaisvindamos visus palikuonių virionus. Simptomai virusinės ligos yra imuninio atsako į virusą, kuris bando kontroliuoti ir pašalinti virusą iš organizmo, taip pat dėl \u200b\u200bviruso sukeltų ląstelių pažeidimų, rezultatas.
Skirtingai nuo kitų mikroorganizmų, turinčių tiek DNR, tiek RNR, virione yra tik viena iš nukleorūgščių - arba DNR, arba RNR.
Virusų nukleorūgštis gali būti viengrandė ir dvigrandė. Beveik visų virusų, turinčių RNR, genome yra vienos grandinės RNR, o tų, kuriuose yra DNR, yra dvigubos grandinės. Pagal dviejų tipų genetinę medžiagą virusai skirstomi į turinčius RNR ir DNR. DNR turinčios šeimos apima 5 šeimas, RNR turinčios - 10 šeimų.
Pradedant procesą, ląstelė nėra lizuojama ar žudoma iš karto. Dauguma produktyvių virusinių infekcijų vykdo panašius viruso replikacijos ciklo etapus: perėjimas, prasiskverbimas, neuždengimas, dauginimasis, surinkimas ir išleidimas. Virusas prisijungia prie specifinės receptoriaus vietos ląstelės-šeimininkės membranoje per prisijungimo baltymus kapsidėje arba baltymus, įdėtus į jo apvalkalą. Prisirišimas yra specifinis, ir paprastai virusas prisijungia tik prie vienos ar kelių rūšių ląstelių ir tik tam tikrų tipų ląstelių iš tų rūšių su atitinkamais receptoriais.
* (Čia pateikiami duomenys tik apie kai kuriuos žmonėms patogeniškus virusus.)
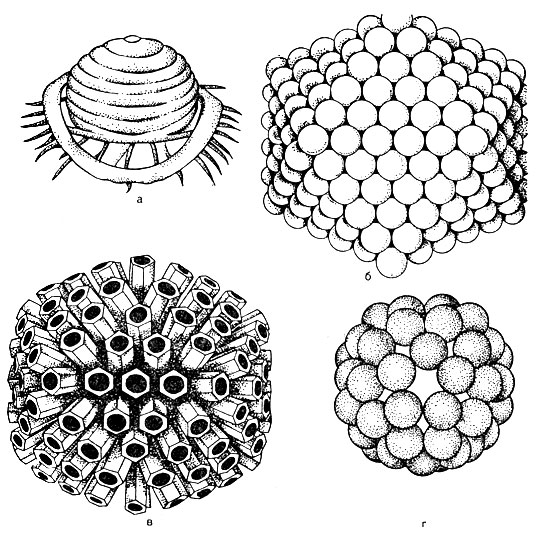
Viriono struktūra... Viriono centre yra nukleino rūgštis, kurią supa kapsidas (iš graikų kanca - dėžutė). Kapsidą sudaro baltymų subvienetai, vadinami kapsomeromis. Subrendęs virusas yra cheminės struktūros nukleokapsidas. Kapsomerų skaičius ir jų supakavimo būdas (52 pav.) Yra griežtai pastovūs kiekvienos rūšies virusui. Pavyzdžiui, poliomielito viruse yra 32 kapsomerai, o adenovirusuose - 252 kapsomerai. Kapsomerai gali būti sukrauti daugiakampio pavidalu su vienodais simetriškais kraštais - stačiakampio formos (pavyzdžiui, adenovirusas). Spiralinis (sferinis) stilius būdingas gripo virusams. Gali būti tam tikros rūšies simetrija, kai nukleorūgštis yra šaltinio formos, aplink kurią klojamos kapsomeros, tokiu atveju virusas yra lazdelės formos - virusas, sukeliantis tabako lapų ligą.
Skirtingai nuo gyvūnų virusų, bakteriofago nukleorūgštis į šeimininko ląstelę įleidžiama nuoga, o kapsidas paliekamas už ląstelės ribų. Augalų ir gyvūnų virusai gali patekti į ląsteles per endocitozę, kurioje ląstelių membrana supa ir apgaubia visą virusą. Kai kurie apgaubti virusai patenka į ląstelę, kai viruso apvalkalas susilieja tiesiogiai su ląstelės membrana. Po to viruso kapsidas suyra ląstelės viduje ir išsiskiria viruso nukleino rūgštis, kuri tampa prieinama replikacijai ir transkripcijai.
Replikacijos mechanizmas priklauso nuo viruso genomo. Žinoma, šiam modeliui yra išimčių. Jei ląstelė-šeimininkė nepateikia fermentų, reikalingų viruso replikacijai, virusų genai teikia informaciją apie tiesioginę trūkstamų baltymų sintezę.
Fagas turi sudėtingą simetrijos tipą: galva yra stačiakampio formos, o procesas yra lazdelės formos (spermos formos) (žr. 21, 22 pav.).
Taigi, priklausomai nuo pakavimo būdo, virusai skirstomi į kuboidines, sferines, lazdelės formos ir spermatozoidines formas.
Paskutinis viruso replikacijos etapas yra naujų virionų išsiskyrimas į šeimininką, kur jie sugeba užkrėsti kaimynines ląsteles ir pakartoti replikacijos ciklą. Kai kurie virusai išsiskiria mirus ląstelei šeimininkei, o kiti virusai gali palikti užkrėstas ląsteles, prasiskverbdami pro membraną, tiesiogiai neužmušdami ląstelės.
6 paveikslas Užkrėsti gripo virusu, glikoproteinai prisijungia prie šeimininko epitelio ląstelės. Dėl to virusas absorbuojamas. 7 paveikslas Virusai sukelia įvairias žmonių ligas, pradedant nesunkiomis ligomis ir baigiant sunkiomis ligomis.
Kai kurie sudėtingesnės struktūros virusai turi apvalkalą, vadinamą peplos. Jis susidaro virusui palikus ląstelę šeimininką. Šiuo atveju viruso kapsidą gaubia ląstelės-šeimininkės citoplazminės membranos vidinis paviršius ir susidaro vienas ar keli superkapsido apvalkalo sluoksniai. Tokį apvalkalą turi tik kai kurie virusai, pavyzdžiui, pasiutligės, herpeso, encefalito virusai. Šiame apvalkale yra fosfolipidų, kuriuos skaido eteris. Taigi, veikiant eteriui, galima atskirti virusą su peplosu nuo viruso, turinčio „nuogą kapsidą“.
Vakciną galima paruošti naudojant susilpnintus gyvus virusus, sunaikintus virusus arba molekulinius viruso vienetus. Apskritai dėl gyvų virusų padidėja imunitetas, tačiau jie gali sukelti ligą tam tikru dažniu. Nužudytos virusinės vakcinos ir subvieneto virusai negali sukelti ligų, tačiau jų imunitetas paprastai būna ne toks efektyvus arba ilgalaikis.
Susilpnintos gyvų virusų vakcinos yra sukurtos laboratorijoje, kad sukeltų mažai simptomų recipientams, tuo pačiu užtikrinant imunitetą nuo būsimų infekcijų. Poliomielitas buvo viena iš ligų, žyminčių vakcinų naudojimo etapą. Poliomielito vakcinos sėkmė atvėrė kelią įprastam vaikų skiepijimui nuo tymų, kiaulytės, raudonukės, vėjaraupių ir kitų ligų. Gyvos vakcinos paprastai gaminamos susilpninant laukinio tipo virusą, auginant jį laboratorijoje audiniuose arba esant kitokiai temperatūrai nei virusas yra įpratęs šeimininkui.
Kai kuriuose virusuose iš išorinio lipidinio sluoksnio sluoksnio kyšo erškėčių formos kapsomeros (šios erškėčiai yra bukai). Šie virusai vadinami peplomerais (pvz., Gripo virusas, žr. 52 pav.).
Viruso nukleorūgštis yra paveldimų savybių nešėja, o kapsidas ir išorinis apvalkalas atlieka apsaugines funkcijas, tarsi apsaugotų nukleorūgštį. Be to, jie palengvina viruso patekimą į ląstelę.
Pavyzdžiui, virusas gali būti auginamas ląstelėse in vitro, paukščių ar gyvų gyvūnų embrionuose. Prisitaikymas prie šių naujų ląstelių ar temperatūros sukelia mutacijas viruso genomuose, leidžiant jiems geriau augti laboratorijoje, tuo pačiu trukdant jų gebėjimui sukelti ligą, kai jie vėl patenka į šeimininko būklę. Taigi šie susilpninti virusai vis dar sukelia infekciją, tačiau jie nelabai auga, todėl laikui bėgant imuninė reakcija gali vystytis, kad būtų išvengta rimtų ligų.
Gyvų vakcinų, kurios paprastai yra veiksmingesnės už nužudytas vakcinas, vartojimo rizika yra maža, tačiau reikšminga rizika, kad šie virusai grįš į savo ligas sukeliančią formą atvirkštinėmis mutacijomis. Atvirkštinės mutacijos atsiranda, kai vakcina mutuoja šeimininke, kad ji perskaitytų šeimininką ir vėl galėtų sukelti ligą, kurią vėliau gali išplisti kiti žmonės, patekę į epidemiją.
Virusų dydžiai... Virusai matuojami nanometrais. Jų dydis svyruoja plačiame diapazone nuo 15-20 iki 350-400 nm.
Virusų matavimo metodai: 1) filtravimas per bakterijų filtrus, kurių porų dydis yra žinomas; 2) ultracentrifugavimas - dideli virusai nusėda greičiau; 3) virusų fotografavimas elektroniniu mikroskopu.
Gripo atveju paviršiaus molekulių genų mutacija padeda virusui išvengti apsauginio imuniteto, kurį jis galėjo gauti praėjusį gripo sezoną, todėl žmonėms būtina skiepytis kiekvienais metais. Kiti virusai, pavyzdžiui, sukeliantys vaikų ligas, tymus, kiaulytę ir raudonukę, mutuoja taip mažai, kad ta pati vakcina naudojama metai iš metų.
Vakcinos ir antivirusiniai vaistai gydymui
Kai kuriais atvejais vakcinomis galima gydyti aktyvią virusinę infekciją. Pasiutligės atveju - mirtina neurologinė liga, perduodama pasiutlige užkrėstų virusų seilėse, ligos progresavimas nuo gyvūno įkandimo iki patekimo į centrinę nervų sistemą gali būti dvi savaites ar ilgiau. Tai yra pakankamai laiko, kad būtų galima paskiepyti asmenį, įtariamą įsisiautėjusio gyvūno įkandimu, o sustiprėjusio vakcinacijos imuninio atsako pakanka, kad virusas nepatektų į nervinį audinį.
Cheminė virusų sudėtis... DNR ir RNR virusų kiekis ir kiekis nevienodi. DNR molekulinė masė svyruoja nuo 1 · 10 6 iki 1,6 · 10 8, o RNR - nuo 2 · 10 6 iki 9,0 · 10 6.
Baltymų virionuose yra nedaug, jie susideda iš 16-20 aminorūgščių. Be kapsidinių baltymų, yra ir vidinių baltymų, susijusių su nukleino rūgštimi. Baltymai lemia virusų antigenines savybes, taip pat dėl \u200b\u200btankios polipeptidinių grandinių pakuotės apsaugo virusą nuo ląstelių-šeimininkų fermentų poveikio.
Tokiu būdu išvengiama mirtinų neurologinių ligos pasekmių ir žmogus turi atsigauti tik po užkrėsto įkandimo. Šis metodas taip pat naudojamas gydant Ebolą - vieną greičiausių ir mirtiniausių virusų, kuriais užkrėsti žmonės, nors dažniausiai tai užkrėsta ribotas populiacijas. Ebola taip pat yra pagrindinė gorilų mirties priežastis. Naudojant naujai sukurtas vakcinas, kurios sustiprina imuninį atsaką, tikimasi, kad paveiktų žmonių imuninė sistema geriau kontroliuos virusą, galėdama sumažinti mirtingumą.
Lipidai ir angliavandeniai yra išoriniame sudėtingų virionų apvalkale. Ląstelių-šeimininkų membrana yra lipidų ir angliavandenių šaltinis. Kai kuriuos virusus sudarantys polisacharidai lemia jų gebėjimą sukelti eritrocitų agliutinaciją.
Viruso fermentai... Virusai neturi savo metabolizmo, todėl jiems nereikia metabolinių fermentų. Tačiau nustatyta, kad kai kurie virusai turi fermentų, kurie palengvina jų įsiskverbimą į ląstelę-šeimininkę. Pavyzdžiui, sergant A gripo virusu, buvo nustatyta neuraminidazė, kuri atskaldo neuramininę rūgštį, esančią gyvūnų ląstelių membranose (eritrocituose ir kt.). Fagai turi lizocimą, kuris sunaikina ląstelės membraną, fosfatazę ir kt.
Kitas būdas gydyti virusines infekcijas yra antivirusinių vaistų vartojimas. Šie vaistai dažnai turi ribotas galimybes išgydyti virusinę ligą, tačiau buvo naudojami kontroliuojant ir mažinant įvairių virusinių ligų simptomus. Daugumai virusų šie vaistai slopina virusą, blokuodami vieno ar daugiau jo baltymų veikimą. Svarbu, kad tikslinius baltymus užkoduotų virusiniai genai ir kad šių molekulių nebūtų sveikoje ląstelėje šeimininkėje.
Taigi viruso augimas yra slopinamas nepažeidžiant šeimininko. Yra daug antivirusiniai vaistaiprieinamos infekcijoms gydyti, kai kurios būdingos tam tikram virusui, o kitos gali paveikti kelis virusus. Antivirusiniai vaistai yra skirti genitalijų herpesui ir gripui gydyti. Sergant lytinių organų pūsleline, tokie vaistai kaip acikloviras gali sumažinti aktyvios virusinės ligos epizodų, kai pacientams atsiranda odos ląstelių virusiniai pažeidimai, skaičių ir trukmę.
Virusinių antigenų nustatymas... Virusinius antigenus užkrėstose šeimininkų ląstelėse galima aptikti naudojant imunofluorescencijos metodus. Preparatai, turintys ląstelių užkrėstas virusuyra gydomi specifiniais imuniniais liuminescenciniais serumais. Žiūrint fluorescuojančiu mikroskopu, būdingas spindesys pastebimas tose vietose, kur kaupiasi viruso dalelės. Viruso tipą lemia specifinio liuminescencinio serumo, sukėlusio liuminescenciją, atitikimas.
Kadangi virusas visą gyvenimą lieka paslėptas organizmo nerviniame audinyje, šis vaistas nėra vaistas, tačiau jis gali padaryti ligos simptomus lengviau valdomus. Sergant gripu, tokie vaistai kaip „Tamiflu“ gali sutrumpinti gripo simptomų trukmę viena ar dviem dienomis, tačiau vaistas visiškai neužkerta kelio simptomams. Kiti antivirusiniai vaistaitokie kaip ribavirinas buvo naudojami įvairioms virusinėms infekcijoms gydyti.
Kai kuris nors iš šių vaistų vartojamas vienas, didelis viruso mutacijų dažnis leidžia virusui greitai sukurti atsparumą vaistui. Virusai yra be ląstelių esybės, kurias paprastai galima pamatyti tik naudojant elektroninį mikroskopą. Virusai yra įvairūs, užkrečia archajas, bakterijas, grybus, augalus ir gyvūnus. Virusai susideda iš nukleorūgšties šerdies, apsuptos baltymo kapsidės su išoriniu lipidų apvalkalu arba be jo.
Viruso įvedimas į ląstelę, jo sąveika su ląstele-šeimininke ir reprodukcija (reprodukcija) susideda iš eilės vienas po kito einančių etapų.
1 etapas. Prasideda adsorbcijos procesas viriono ir ląstelių receptorių sąskaita. Kompleksiniuose virionuose receptoriai yra ant voko paviršiaus kaip styloidinės ataugos (gripo virusas), paprastuose virionuose - ant kapsidės paviršiaus.
2 etapas. Skirtingų virusų virusas prasiskverbia į ląstelę-šeimininkę skirtingai. Pavyzdžiui, kai kurie fagai savo procesu perveria membraną ir injekuoja į šeimininko ląstelę nukleorūgštį (žr. 8 skyrių). Kiti virusai patenka į ląstelę patekdami į viruso dalelę vakuolės pagalba, tai yra, įvedimo ląstelės membranoje vietoje susidaro depresija, tada jos kraštai uždaromi ir virusas atsiranda ląstelėje. Šis atitraukimas vadinamas viropexis.
3 etapas. „Viruso pašalinimas“ (suirimas). Reprodukcijai virusinė nukleorūgštis atlaisvinama iš apsauginių baltymų dangalų (apvalkalo ir kapsidės). Nusirengimas gali prasidėti adsorbcijos metu, arba jis gali įvykti, kai virusas jau yra ląstelės viduje.
4 etapas. Šiame etape vyksta nukleorūgščių replikacija (reprodukcija) ir virusinių baltymų sintezė. Šis etapas vyksta dalyvaujant ląstelės-šeimininkės DNR arba RNR.
5 etapas. Viriono surinkimas. Šį procesą palengvina pats baltymų dalelių susidarymas aplink viruso nukleino rūgštį. Baltymų sintezė gali prasidėti iškart po viruso nukleorūgščių sintezės arba po kelių minučių ar kelių valandų pertraukos. Kai kuriems virusams savaiminė jungtis vyksta citoplazmoje. Kiti branduolyje turi ląsteles-šeimininkes. Citoplazmoje visada susidaro išorinis apvalkalas (peplos).
6 etapas. Virionas išsiskiria iš ląstelės-šeimininkės, per virusą perkratant per ląstelės membraną arba per skylę, susidariusią ląstelėje-šeimininkėje (šiuo atveju ląstelė-šeimininkė miršta).
Virusų ir ląstelių sąveikos tipai... Pirmasis tipas, produktyvi infekcija, būdinga naujų virionų susidarymui ląstelėje šeimininkėje.
Antrasis tipas - abortinė infekcija - nukleorūgščių replikacija nutrūksta.
Trečiajam tipui būdinga viruso nukleino rūgšties įtraukimas į ląstelės-šeimininkės DNR; yra viruso ir ląstelės-šeimininkės sambūvio forma (virogenija). Šiuo atveju užtikrinamas virusų ir ląstelių DNR replikacijos sinchroniškumas. Faguose tai vadinama lizogenija.
Mikroskopinis tyrimas... Esant individualioms virusinėms infekcijoms, šeimininko ląstelių citoplazmoje arba branduoliuose pastebimi specifiniai tarpląsteliniai kūnai - inkliuzai, turintys diagnostinę vertę (pasiutligės atveju - Babesh-Negri kūnai, raupuose - Guarnieri kūnai ir kt.). Virusinių dalelių ir kūnų-inkliuzų dydžius galima dirbtinai padidinti taikant specialius preparatų apdorojimo su kandikliais ir impregnavimu metodus (pavyzdžiui, sidabravimo metodą pagal Morozovą) ir stebint panardinimo mikroskopu. Mažesni virionai, esantys už optinio mikroskopo matymo ribų, aptinkami tik elektronine mikroskopija. Ląstelių intarpų požiūrių yra įvairių. Kai kurie autoriai mano, kad jie yra virusų kolekcija. Kiti mano, kad jie atsiranda dėl ląstelės reakcijos į virusų patekimą.
Virusų genetika... Virusų modifikacija (nepaveldimi pokyčiai) atsiranda dėl ląstelės-šeimininko, kurioje virusas dauginasi, savybių. Modifikuoti virusai įgyja galimybę užkrėsti ląsteles, panašias į tas, kuriose jie buvo modifikuoti. Skirtingi virusai pasireiškia įvairiai. Pavyzdžiui, faguose keičiasi „neigiamų dėmių“ (fagų kolonijų) forma.
Mutacija - virusuose ji įvyksta veikiama tų pačių mutagenų, kurie sukelia bakterijų mutaciją (fiziniai ir cheminiai veiksniai). Nukleorūgščių replikacijos metu įvyksta mutacija. Mutacijos turi įtakos įvairioms virusų savybėms, pavyzdžiui, jautrumui temperatūrai ir kt.
Genų rekombinacija virusuose gali atsirasti tuo pačiu metu užkrėtus ląstelę šeimininkę dviem virusais, tuo tarpu atskirais genais galima keistis tarp dviejų virusų ir susidaro rekombinantai, turintys dviejų tėvų genus.
Genetiškai vėl suaktyvėja genai, kai inaktyvuotas virusas kertamas su visišku, o tai veda prie inaktyvuoto viruso išgelbėjimo.
Spontaniška ir nukreipta virusų genetika turi didelę reikšmę vystantis infekciniam procesui.
Atsparus aplinkos veiksniams... Dauguma virusų yra inaktyvuojami veikiant aukšta temperatūra... Tačiau yra išimčių, pavyzdžiui, hepatito virusas atsparus karščiui.
Į žema temperatūra virusai nėra jautrūs, ultravioletiniai saulės spinduliai turi inaktyvuojantį poveikį virusams. Išsibarsčiusios saulės spinduliai juos veikia mažiau aktyviai. Virusai yra atsparūs gliceroliui, todėl juos galima ilgai laikyti glicerine. Jie atsparūs antibiotikams (auginant virusus tiriamoji medžiaga apdorojama antibiotikais, kad nuslopintų bakterijų florą).
Rūgštys, šarmai, dezinfekcijos priemonės inaktyvuoja virusus. Tačiau kai kurie virusai, inaktyvinti formalinu, išlaiko savo imunogenines savybes, o tai leidžia naudoti formaliną vakcinoms (pasiutligės vakcinai) gaminti.
Gyvūnų jautrumas... Kai kuriems virusams imlių gyvūnų spektras yra labai platus, pavyzdžiui, daugelis gyvūnų yra jautrūs pasiutligės virusams. Kai kurie virusai veikia tik vieną gyvūnų rūšį, pavyzdžiui, šunų maro virusas veikia tik šunis. Yra virusų, kuriems gyvūnai nėra imlūs - pavyzdžiui, tymų virusas ir kt.
Organotropija virusai... Virusai gali užkrėsti tam tikrus organus, audinius ir sistemas. Pavyzdžiui, pasiutligės virusas atakuoja nervų sistemą. Raupų \u200b\u200bvirusas yra dermatropinis ir kt.
Virusų patekimas į aplinką... Iš sergančio kūno virusai gali išsiskirti su išmatomis, pavyzdžiui, poliomielito virusu ir kitais enterovirusais. Pasiutligės virusas išsiskiria su seilėmis, gripo virusas išskiriamas iš nosiaryklės gleivinės ir kt.
Pagrindiniai virusų perdavimo būdai... Oro lašeliai (gripas, raupai), maistas (poliomielitas, hepatitas A), kontaktas su buitimi (pasiutligė), užkrečiamas (encefalitas).
Antivirusinis imunitetas... Žmogaus organizmas turi įgimtą atsparumą tam tikriems virusams. Pavyzdžiui, žmonės nėra jautrūs šunų maro virusui. Gyvūnai nėra imlūs tymų virusui. Šiais atvejais antivirusinis imunitetas pagrįstas ląstelių, galinčių palaikyti virusų dauginimąsi, nebuvimu.
Antivirusinį imunitetą lemia tiek ląstelių, tiek humoralinės gynybos veiksniai, nespecifiniai ir specifiniai. Nespecifiniai veiksniai. Galingas viruso dauginimosi inhibitorius yra baltyminė medžiaga - interferonas. IN sveikas kūnas jo yra nereikšmingu kiekiu, o virusai prisideda prie interferono gamybos ir jo kiekis žymiai padidėja. Jis yra nespecifinis, nes blokuoja įvairių virusų dauginimąsi. Tačiau jis turi audinių specifiškumą, tai yra, skirtingų audinių ląstelės sudaro skirtingą interferoną. Manoma, kad jo veikimo mechanizmas slypi tame, kad jis trukdo ląstelių-šeimininkų baltymų sintezei ir taip sustabdo viruso dauginimąsi.
Specifiniai antivirusinio imuniteto veiksniai yra virusus neutralizuojantys antikūnai, hemagliutinuojantys ir nusodinantys antikūnai.
Virusų auginimo metodai... Virusai dauginasi tik gyvybingose \u200b\u200bląstelėse. Jie auginami: vištienos embrionuose (53 pav.), Žmonių ir įvairių gyvūnų audinių kultūrose, jautrių gyvūnų, imlių nariuotakojų kūne.
Pirmuoju virusologijos vystymosi laikotarpiu pagrindinis virusų tyrimo metodas buvo dirbtinė gyvūnų infekcija, tačiau šis metodas yra komplikuotas, be to, gyvūnai pasirodė esą apsaugoti nuo daugelio virusų.
Didelę reikšmę plėtojant virusologiją turėjo virusų auginimo vištienos embrionuose ir žmogaus bei gyvūnų audinių ląstelių kultūroje metodų įvedimas.
Vištienos embriono infekcija... Virusams dauginti naudojami 7–12 dienų amžiaus viščiukų embrionai, inkubuojami termostate 37 ° C temperatūroje. Būtina sąlyga teisingam embriono vystymuisi yra tam tikros oro drėgmės laikymasis, kuris gali būti sukurtas indą su vandeniu dedant į termostatą.
Vištienos embriono tinkamumą infekcijai lemia embriono judesiai ir išplėtotas kraujagyslių tinklas ant choriono-alantoicinės membranos skenuojant ovoskopu.
Viščiukų embrionuose virusai auginami skirtingose \u200b\u200bužkrėstose embriono vietose (žr. 53 pav.):
1) ant choriono-alantoicinės membranos,
2) į alantoicinę ertmę;
3) į vaisiaus ertmę;
4) į trynio maišelį.
Vištienos embrionai užkrėsti dėžutėje naudojant sterilius instrumentus. Prieš užkrėtimą vištienos embrionai du kartus nuvalomi alkoholiu suvilgytu medvilniniu tamponu.
Infekcija ant choriono-alantoicinės membranos. Po dezinfekcijos kiaušiniai atsargiai nupjauna apvalkalo gabalą nuo buko galo, pašalinama lukšto membrana ir randama choriono-alantoicinė membrana. Infekcinė medžiaga 0,1–0,2 ml, naudojant švirkštą arba Pasteur pipetę, uždedama ant choriono-alantoicinės membranos. Po infekcijos skylė uždaroma dangteliu, o tarpas tarp jos ir viščiuko embriono užpildomas parafinu.
Kitoje kiaušinio pusėje pieštuku užrašykite infekcinės medžiagos pavadinimą ir užkrėtimo datą.
Infekcija į vaisiaus ertmę. Kiaušinis yra ovoskopuotas, o šoninėje pusėje parenkama vieta, kur chorionas-alantois neturi didelių kraujagyslių. Ši sritis pažymėta pieštuku. Kiaušiniai dedami ant stovo horizontalioje padėtyje, dezinfekuojami ir specialia sterilia ietimi iki 213 mm gylio perveriama skylė lukšte, per kurią tuo pačiu atstumu tiesiai į vaisiaus vandenų ertmę įkišama adata su infekcine medžiaga. Kad įpurškiamas skystis nenutekėtų atgal, virš oro pagalvės padaroma punkcija, po kurios abi skylės užpildomos parafinu.
Infekcija į alantoic ertmę. Infekcija atliekama užtamsintoje dėžutėje. Pažymima oro erdvė, apvalkalas virš oro erdvės dezinfekuojamas ir švirkšto adata su medžiaga įkišama per kiaute esančią angą link embriono. Jei adata pateko į alantoicinę ertmę, tada embriono šešėlis pasislinko. Po infekcijos skylė užpildoma parafinu.
Infekcija trynio maišelyje. Korpusas dezinfekuojamas. Kiaušinis dedamas ant stovo buku galu į dešinę taip, kad trynio maišelis būtų nukreiptas į viršų. Skylė pradurta virš oro kameros centre. Per lukšto skylę horizontalia kryptimi iki 2-3 mm gylio įkišama švirkšto adata, kuri patenka į trynio maišelį. Medžiaga įpurškiama 0,2–0,3 ml tūrio. Įvedus medžiagą, skylė vaškuojama.
Temperatūros režimas ir inkubacijos trukmė priklauso nuo įvesto viruso biologinių savybių.
Užkrėsti kiaušinėliai tikrinami kasdien - atliekant ovoskopiją, siekiant patikrinti embriono gyvybingumą. Jei embrionai miršta pirmą dieną, tai dažniausiai būna traumos infekcijos metu. Tokie kiaušiniai yra gauti iš patirties.
Jei būtina atskirai ištirti kiekvieną embriono komponentą, medžiaga surenkama tam tikra tvarka: išsiurbiamas alantoicinis skystis, tada nupjaunama vaisiaus vandenys, choriono-alantoicinė membrana, atskiriama vaisiaus vandenų membrana, embrionas, trynio maišelis ir tik tada pašalinama choriono-alantoicinė membrana, atskiriant ją nuo vidinės. apvalkalo paviršius. Viruso buvimą užkrėstame embrione lemia būdingi užkrėstos vištienos embriono choriono-alantoicinės membranos pokyčiai.
Virusai, neturintys hemagliutinuojančio aktyvumo, aptinkami naudojant CSC.
Norėdami aptikti virusą užkrėstų embrionų alanto arba amniono skysčiuose, dedama RHA (hemagliutinaciją sukelia alanto arba amniono skysčiai arba suspensija, paruošta iš choriono-alantoicinės membranos).
Virusų kultivavimas ląstelių kultūroje... Virusams kauptis jautriose ląstelių kultūrose naudojami žmonių ir įvairių gyvūnų audiniai. Didžiausią praktinį pritaikymą gavo pirminių tripsinizuotų ir persodintų ląstelių linijų vieno sluoksnio kultūros.
Vieno sluoksnio ląstelių kultūros auginamos plokščio stiklo čiužinių induose. Ląstelių suspensija skystoje maistinėje terpėje 37 ° C temperatūroje leidžia gauti "in vitro" tam tikros histologinės struktūros ląstelių sluoksnį. Virusų buvimas audinių kultūrose nustatomas pakitus (degeneravus) ląsteles. Virusų tipas nustatomas neutralizuojant virusų poveikį, į vakcinuotą medžiagą pridedant atitinkamų tipų serumų.
Šie metodai leidžia greičiau įtraukti tyrimų rezultatus ir yra ekonomiškesni. Tais atvejais, kai virusai nesukelia citopatinio poveikio (degeneracijos) ir nesivysto viščiukų embrionuose, naudojami gyvūnų užkrėtimo metodai (žr. 11 skyrių).
Virusams auginti naudojamos nepertraukiamos ląstelės, kurios dažniau gaunamos iš piktybinių navikų ląstelių.
Vieno sluoksnio kultūros gaunamos iš žmogaus, vištienos, gyvūnų embrionų.
Vieno sluoksnio ląstelių kultūrų privalumas yra technikos paprastumas ir apskaitos paprastumas.
Ląstelių gebėjimas daugintis už kūno ribų yra susijęs su audinių diferenciacijos laipsniu. Mažiau diferencijuoti audiniai turi didesnį gebėjimą daugintis (jungiamasis, epitelinis audinys).
Pirminių audinių kultūrų paruošimo metodų esmė yra tarpląstelinio audinio sunaikinimas ir ląstelių atskyrimas tolesniam monosluoksnio gamybai.
Ląstelių disociacija atliekama veikiant proteolitinių fermentų, dažniausiai tripsino, audinį. Tripsino tirpalas skatina ląstelių atsiskyrimą, išlaikant jų gebėjimą daugintis. Norint auginti ląstelių kultūras, reikalinga maistinė terpė. Terpės sudėtis yra sudėtinga, joje yra daugybė ingredientų: aminorūgštys, gliukozė, vitaminai, mineralinės druskos, kofermentai ir kt. Audinių kultūra gaunama griežtai aseptinėmis sąlygomis. Antibiotikai (500 V penicilino ir 250 V streptomicino 1 ml) pridedami prie terpės, kad būtų slopinamas bakterinės floros augimas.
Paruoštas audinys pilamas 0,25% pašildyto tripsino tirpalu ir inkubuojamas termostate 37 ° C temperatūroje. Inkubacijos metu audinys periodiškai maišomas sukant kolbą. Trypsinizuotos ląstelės centrifuguojamos 800-1000 aps./min. 5 minutes.
Tripsinizavimas ir centrifugavimas atliekamas labai atsargiai, kad nesužeistų ląstelių. Po centrifugavimo supernatantas pašalinamas ir ląstelių nuosėdos dedamos į nedidelį tūrį terpės. Norint gauti vienalytę masę, ląstelių suspensija filtruojama per vieną marlės sluoksnį piltuve (sterili). Ląstelių suspensijos sterilumas tikrinamas inokuliuojant 0,1 ml į 2 tūbeles su cukraus sultiniu.
Ląstelių kultivavimo sėkmė priklauso nuo inokuliacijos dozės, todėl po tripsinizavimo ląstelės skaičiuojamos Gorjajevo kameroje. Suskaičiavus, ląstelių suspensija praskiedžiama maistine terpe taip, kad 1 ml būtų 500 000–1 000 000 ląstelių ir supilama į mėgintuvėlius ir čiužinius. Audinių kultūros mėgintuvėliai inkubuojami inkubatoriuje pasvirusioje padėtyje.
Inokuluotos kultūros kasdien stebimos mažo didinimo mikroskopu, siekiant nustatyti jų augimo modelius. Normaliai dauginančios ląstelės yra šviesios spalvos ir auga vienu sluoksniu. Jei ląstelės yra tamsios, grūdėtos ir nesidaugina, o tai gali būti užteršimo (netinkamo indų tvarkymo ar ingredientų užteršimo) rezultatas, tokios kultūros pašalinamos iš eksperimento.
Maistinės terpės pakeitimas praėjus 2-3 dienoms po sėjos pagerina proliferacijos intensyvumą.
Normalios, gerai dauginančios ląstelės yra užkrėstos tiriamąja medžiaga.
Persodintos kultūros daugiausia gaunamos iš piktybinių navikų. Helos padermė - moters, vardu Helena, gimdos kaklelio vėžio ląstelių kultūra (gauta 1950 m.); Hep-2 padermė išskirta iš paciento, sergančio gerklų vėžiu. Šių ląstelių augimas palaikomas laboratorijose atliekant nuoseklius pasažus. Jų ypatumas slypi tame, kad jie dauginasi ilgai. Šiuo metu šios ląstelės išgyveno tūkstančius kartų. Perėjimo metu jie praranda kai kurias morfologines ir biochemines savybes - patiria mutacijas. Tačiau jie išlieka gana tinkami juose auginti virusus. Šių ląstelių kultūrą naudoja viso pasaulio laboratorijos.
Virusas dauginasi ląstelių kultūroje skirtingu laiku, priklausomai nuo viruso savybių ir ląstelių tipo.
Viruso buvimas sprendžiamas pagal citopatinį poveikį. Mikroskope stebima ląstelių degeneracija. Citopatinio veikimo laikas ir jo pobūdis priklauso nuo viruso dozės ir savybių.
Kai kuriems virusams citopatinis poveikis nustatomas po kelių dienų (raupų virusas), kitiems - po 1–2 savaičių (hepatito virusas ir kt.).
Šiuo metu jau yra šimtai žmonių užkrėstų virusų. Kova su virusinėmis infekcijomis vykdoma skirtingais metodais. Efektyviausia yra imunizacija. Tokiu būdu buvo pašalintos raupai, sumažėjo poliomielito atvejų. Kovojant su virusinėmis infekcijomis svarbi socialinė prevencija - benamių šunų naikinimas (kova su pasiutlige), asmeninė prevencija ir kt.
Tačiau šios priemonės negali užtikrinti visų virusinių ligų pašalinimo. Mokslininkai atkakliai ieško būdų, kaip būtų galima užkrėsti virusą, nepažeidžiant ląstelės, kurioje jis yra.
Todėl natūralu, kad Sovietų Sąjungos komunistų partijos programoje virusologija įvardijama kaip viena iš pirmaujančių gamtos mokslo šakų, kuri artimiausiais metais turėtų būti plėtojama prioritetiškai.
Pagrindiniai virusų tyrimo metodai... 1. Hemagliutinacijos reakcija, uždelstos hemagliutinacijos reakcija, netiesioginės hemagliutinacijos reakcija. Komplemento rišimosi reakcija.
2. Virusų neutralizavimo audinių kultūroje reakcija.
3. Imunofluorescencijos metodas.
4. Histologinis metodas - intarpų nustatymas (maži Babesho kūnai - Negri - su pasiutlige; maži Pasheno kūnai - su raupais ir kt.).
5. Biologinis metodas.
Virusinė infekcija ląstelė yra procesų kompleksas, atsirandantis dėl ląstelės sąveikos su viruso genomu. Tarpląstelinis virionas yra inertiškas, ir šis inertiškumas išlieka tol, kol viruso genomas pradeda veikti ląstelėje, kaip savarankiškas genetinis vienetas. Pagrindinė žemiau pateiktos klasifikacijos ypatybė yra santykis tarp viruso ir ląstelės genomų. Jei viruso genomas kartojasi nepriklausomai nuo ląstelių genomo replikacijos, tai yra autonominis virusinės infekcijos tipas. Jei virusinis genomas yra įtrauktas į ląstelių genomą ir kartojasi kartu su juo, tai yra integracinis tipas. Kiekvieną iš šių dviejų pagrindinių virusinės infekcijos tipų galima suskirstyti į kelias formas, atsižvelgiant į viruso susidarymą (produktyvi ir abortinė infekcija), atsižvelgiant į ląstelės infekcinio proceso rezultatus (lizinė ir nelitinė infekcija), atsižvelgiant į viruso ir ląstelės sąveikos dinamiką (ūminė ir lėtinė infekcija). ).
Autonominis virusinės ląstelės infekcijos tipas būdingas daugumai žinomų gyvūnų ir žmonių virusų. Šio tipo infekcijose viruso genomas dauginasi nepriklausomai nuo ląstelių genomo replikacijos.
Produktyviai autonominei virusinei infekcijai būdingas visiško viruso susidarymas. Tai yra labiausiai ištirta virusų ir ląstelių sąveikos forma. Pagrindiniai viruso reprodukcijos mechanizmai, kurie buvo aptarti 3 skyriuje, buvo ištirti ir aprašyti šiai konkrečiai infekcijos formai.
Autonominė abortinė virusinė infekcija įvyksta, jei dėl viruso sąveikos su ląstele nesusidaro visiškai vertingi virionai. Paprastai tai įvyksta, kai ląstelės yra užkrėstos brokuotu virusu; kai virusas prasiskverbia į jam atsparias ląsteles; kai imlios ląstelės neleistinomis sąlygomis yra užkrėstos visiškai vertingu virusu.
Lytinė autonominė virusinė infekcija lemia ląstelių mirtį. Ląstelių mirtį sukeliantis virusas vadinamas citopatogeniniu, o ląstelių pažeidimas žymimas terminu „citopas, togeninis poveikis“. Citopatologinių virusinių infekcijų pokyčių pobūdis yra labai įvairus. Tai gali būti degu
Ląstelių nepakankamumas (citoplazmos vakuolizacija, branduolinė piknozė, chromosomų aberacijos), specifinių virusinių tarpląstelinių inkliuzų susidarymas, milžiniškų ląstelių, tokių kaip polikariocitai ar simplastos, atsiradimas. Ląstelių lizė yra paskutinis citopatogeninių virusų veikimo etapas. Kad pasireišktų citopatogeniškumas, virusui nereikia atkurti genomo ir įgyvendinti visos genetinės informacijos.
Nelitinė autonominė virusinė infekcija būdinga virusams, kurių išorinis apvalkalas yra lipoproteinas ir pumpuruodami siunčia ląsteles. Retro, ortomikso, paramikso, arenavirusai gali sukelti produktyvią ne lizinių ląstelių infekciją. Vis dėlto dažniausiai abortinėms infekcijoms būdingas nelitinis pobūdis, kai nėra arba neatsiranda genetinių virusų, reikalingų citopatogeninėms funkcijoms įgyvendinti, savybių.
Pagal infekcinio proceso dinamiką virusinės infekcijos skirstomos į ūmines ir lėtines. Kada ūminė infekcija užkrėstoje ląstelėje atsiranda virionų susidarymas, po kurio infekcinis procesas sustoja. Sergant lėtine infekcija, užkrėsta ląstelė gamina virionus ar virusinius komponentus iki natūralios mirties (ne dėl tam tikros virusinės infekcijos), o dukterinės ląstelės lieka užkrėstos. Produktyvi ir abortinė lytinė infekcija visada yra ūmi. Produktyvi ne lizinė infekcija visada yra lėtinė. Abortinė ne lizinė infekcija, kaip taisyklė, turi lėtinę eigą, tačiau ji taip pat gali pasireikšti ūmine forma.
Integraciniam virusinės infekcijos tipui būdinga fizinė virusų ir ląstelių genomų sąjunga (integracija). Šiuo atveju viruso genomas kartojasi ir veikia kaip neatskiriama korinio genomo dalis. Šis virusinės infekcijos tipas yra privalomas retrovirusams ir yra įmanomas papoviruso, adeno ir herpeso virusų atveju.
Integracines infekcijas galima klasifikuoti pagal tuos pačius parametrus kaip ir autonomines - produktyvumą, dinamiką ir baigtį; integracijos ryšys tarp viruso ir ląstelės taip pat gali būti produktyvus ir abortinis. Produktyvi integracinė infekcija būdinga retrovirusams. Abortas, integruota retrovirusinė infekcija, vyksta tomis pačiomis sąlygomis, kaip aprašyta aukščiau autonominėms abortinėms infekcijoms. Taigi trūkumų turinčių retrovirusų nėra
Naujos genomo dalys (kai kurios Rous sarkomos viruso padermės, pelių, kačių, beždžionių sarkomos virusai, Frend ir Rauscher pelių leukemijos virusai ir kt.) Savaime gali sukelti tik abortyvią integracinę infekciją, kuri esant virusui gali virsti produktyvia infekcija asistentas. Abortyvios integracinės infekcijos, kurią sukelia visiškai vertingas retrovirusas, neleidžiančiose ląstelėse, pavyzdys yra infekcija paukščių virusas Rous žinduolių ląstelių sarkoma. Tuo pačiu metu ląstelėse vyksta abortinis integracijos procesas, kuriam būdinga sumažėjusi virusinės RNR gamyba, neužbaigto virusinių baltymų asortimento kaupimasis, jų apdorojimo sutrikimas ir virionų susidarymo stoka.
Paprastai integracinės infekcijos yra nelitinės, kaip ir B, C, D tipų retrovirusų, onkogeninių papovo, adeno, herpeso virusų atveju, tačiau yra ir lizinių infekcijų, kurias sukelia paukščių retikuloendoteliozės citopatogeninis retrovirusas arba „putojantis“ virusas.
Virogeninė integracinė infekcija yra abortinė infekcija, kurią galima paversti produktyvia forma. Ląstelės su šia infekcijos forma gali spontaniškai arba veikiamos specialios (fizinės, cheminės, biologinės) įtakos, gali sukelti infekcinį virusą. Šis reiškinys vadinamas virogenija, o ląstelės yra vadinamos virogeninėmis pagal analogiją su lizogenija ir lizogeninėmis fagų-bakterijų sistemos ląstelėmis.
Daugiau apie temą Virusinių ląstelių infekcija:
- Virusinės infekcijos ir įtariamos virusinės etiologijos infekcijos
- Bakterinės ir virusinės oro pernešamos infekcijos: gripas, paragripas, adenovirusinė infekcija, respiracinė sincitinė infekcija. Bakterinė bronchopneumonija, lobarinė pneumonija.