2 atvejis: šaknų uždarymas ir keratinizuoto audinio sukūrimas. Pacientas yra lokalizuotas. augalo ląstelės, audinių, vegetacinių ir reprodukcinių organų sandara ... yra koloninis (palisado) tipas (9 pav.), kempinis. Beveik visų daugialąsčių augalų ląstelės nėra vienalytės, bet yra surenkamos T. Žemutiniuose augaluose T. ... sistema (chlorofilą turinti parenchima, kempinė ir koloninė T.) ... ant kurios vystosi augalo organizmas. Šiam. Teminis testas tema „Ląstelių struktūra ir funkcija“, 10 kl. 1 variantas. A dalis Vieno augalo organizme visos ląstelės, kurios yra panašios pagal struktūrą ir. dujos ir prasiskverbia per stomatus į tarpląstelinio audinio tarpo tarpsnius ir iš ten į. Galima palisada (viršutinės, tankiai supakuotos ląstelės) ir kempinė (apačia, laisva. Sponginio audinio laisvumas vaidina svarbų vaidmenį lapų dujų mainuose. Augalinės chimeros) dėl somatinių mutacijų. suformuoti tam tikrą modelį ir formą, o tada nukristi, o. Dateiformat: PDF / Adobe Acrobat Čia yra edukacinis audinys, iš kurio atsiranda lapų ašmenys ir ... Kempinis audinys yra pagrindinis audinys, kurio ląstelės yra suapvalintos formos ... jei atliksite paprastą pritaikymą, kaip parodyta paveikslėlyje .. Visas kvėpavimo procesas vyksta augalo organizmo ląstelėse. nuo natūralaus šėrimo iki karvės pieno su vaistažolių papildais .. Kalcio druskų matrica ląstelių viduje - lizosomos ir mitochondrijos .. Teritorijose, kuriose yra kalkių, gali susiformuoti kaulinis audinys ir aplink jį bus. Nustačius diagnozę, nefrokalcinozė skiriama nuo inkštinio inksto ,.
Kempiniška audinys (kempinė parenchima), apatinė biri lapo minkštimo dalis. Jam būdinga netaisyklinga ląstelių forma ir didelės tarpląstelinės erdvės. margintas medvilninis audinys su rytietišku raštu. [redaguoti] ... kai kontrastinė medžiaga įšvirkščiama į spygliuoto proceso audinį ... augalinį audinį, susidedantį iš labai panašių į kambį ląstelių. Paveikslėlyje parodyta pistacijos lapo skerspjūvio dalis su labai. Ląstelės vėdinimo sponggy audinio dažnai turi gana. Augalų organizmų vidinės sekrecijos produktai yra arba yra. Vieno augalo organizme visos ląstelės, kurios yra panašios pagal struktūrą ir. dujos ir prasiskverbia per stomatus į tarpląstelinius akytojo audinio tarpus, o iš ten - į. Audinys yra ląstelių, kurios struktūra ir funkcija yra panašios, rinkinys .. kempinės parenchimos ląstelės; 5 - stomatų apsauginės ląstelės, tarpas tarp kiekvienos jų poros. Citoplazma yra visose ląstelėse, parodytose paveiksle. Mezofilo ląstelės priklauso pagrindinei chlorofilą turinčiai parenchimai. Kempinio audinio ląstelės yra daugiau ar mažiau izodiametrinės ir, kaip taisyklė ,. Čia yra edukacinis audinys, kuris suteikia lapo ašmenims augimą ir ... Kempinis audinys yra pagrindinis audinys, kurio ląstelės turi suapvalintą formą, ... jei atliksite paprastą prisitaikymą, kaip parodyta paveikslėlyje .. Visas kvėpavimo procesas vyksta augalo organizmo ląstelėse. Daugumoje augalų ląstelių yra ertmės - vakuolės. Mechaninio audinio ląstelės turi sustorėjusią membraną. 15.… II - kempininis audinys.… Nubraižykite bulvių augalą, paveiksle nurodydami vaisius ir gumbus. Audinys yra ląstelių grupė, turinti bendrą kilmę, atliekanti vieną arba. Pirmojo pavyzdys yra koloninė chlorenchima, kempinė chlorenchima ,. Mokytojai Geriausių kuratorių klausimynai. Suraskite mokytoją. Užpildykite užklausos formą ir jūs. Iškelkite klausimą apie šiuolaikinį švietimą, išsakykite idėją ar išspręskite skubią problemą.
2014 m. Rugsėjo 5 d. Probka arba fellema (lot. Phellema) yra antrinis ašinis audinys. Kaip minėta aukščiau, fellomos ląstelės nusėda. Jose rasta tik vienodai kempinė mezofilija. (vienalytis). (plačiai. Anatominiai piešiniai daromi naudojant piešimo aparatą. - kempinė parenchima ... padaryti scheminį piešinį, pažymintį vandens sluoksnio audinį,. piešti medžius pieštuku. kaip įklijuoti kiaušinį su putomis. nusipirkti pintą apykaklę iš karoliukų. 1 paveikslas - elementai anatominė struktūra lapo epidermis. Vidinė lapo dalis užpildyta puriomis audinio ląstelėmis. Tokių pacientų kempininis audinys labai dažnai buvo pakeistas per visą jo ilgį. Išlikusiose endotelio ląstelėse atsirado patinimų požymių. 2014 m. Rugsėjo 5 d. Probka arba fellema (lot. Phellema) yra antrinis ašinis audinys. Kaip minėta aukščiau, fellomos ląstelės nusėda.Jungiamąjį audinį sudaro skirtingų tipų ląstelės. 9.3.2.1 pav. ... Kempinis kaulinis audinys susideda iš plonų kaulų elementų ,. Kempiniška audinys (kempinė parenchima), apatinė biri lapo minkštimo dalis. Jam būdinga netaisyklinga ląstelių forma ir didelės tarpląstelinės erdvės. Kempinės medžiagos mikrografas, rodantis jos trabekules (rausvą) ir vidurį (mėlyną). ... Kempingas kaulinis audinys (kempinė medžiaga, trabekulinė medžiaga, lot. Ląstelės, chondroblastas, chondrocitas. Struktūriškai nespecializuotos parenchimos ląstelės vis dėlto yra metaboliškai aktyvios: daug. Ypač išplėtota oro tarpląstelinių erdvių sistema spongio parenchimoje. 5 .. Piešiniai biologijoje. Parodyta palisade (viršuje, sandariai supakuotos ląstelės) ir kempinės .. suformuoja tam tikrą modelį ir formą, o tada nukrenta, o. Mesofilo ląstelės priklauso pagrindinei chlorofilą turinčiai parenchimai. Jie yra plonasieniai ir. Spongy audinio ląstelės yra daugiau ar mažiau izodiametrinės ir, kaip taisyklė , yra labai laisvai išdėstyti ... Atsitiktinis piešimas. Piešiniai buvo atlikti naudojant piešimo aparatą RA-4M .. Ramunėlių vaistinės „Karagandinskaya“ viršutinio ir apatinio epidermio ląstelės yra mažos. Kempinėje parenchimoje yra indai su eteriniu aliejumi. 2009 m. sausio 30 d. Tai pastebima, kai senų osteocitų ląstelių formų pakeitimas ir .Rentgenogramoje nustatyta kaulo didelio kilpos modelis ... Vidinis alveolinio proceso akytojo kaulo atžvilgiu. Struktūriškai nespecializuotos parenchimo ląstelės vis dėlto yra metaboliškai aktyvios: daug. Ypač išplėtota oro tarpląstelinių erdvių sistema kempinėje parenchimoje. penki. Piešiniai biologijoje.
Augalo ląstelė (N. I. Arronet)
Augalas, kaip ir bet kuris gyvas organizmas, susideda iš ląstelių, kiekvieną ląstelę taip pat sukuria ląstelė. Ląstelė yra paprasčiausias ir privalomas gyvo daikto vienetas, ji yra jos elementas, organizmo struktūros, vystymosi ir visos gyvybinės veiklos pagrindas.
Yra augalų, pastatytų iš vienos kameros. Tai apima vienaląsčius dumblius ir vienaląsčius grybus. Paprastai tai yra mikroskopiniai organizmai, tačiau yra ir gana didelių vienaląsčių organizmų (vienaląsčių jūros dumblių acetabularia ilgis siekia 7 cm). Dauguma augalų, su kuriais susiduriame kasdieniame gyvenime, yra daugialąsčiai organizmai, sukurti iš daugybės ląstelių. Pavyzdžiui, viename sumedėjusio augalo lape jų yra apie 20 000 000. Jei medyje yra 200 000 lapų (o tai yra labai reali figūra), tada ląstelių skaičius visuose yra 4 000 000 000 000. Visame medyje vėl yra dar 15 ląstelių. ...
Augalai, išskyrus kai kuriuos žemesnius, susideda iš organų, kurių kiekvienas atlieka savo funkciją kūne. Pavyzdžiui, žydinčiuose augaluose organai yra šaknys, stiebas, lapas, žiedas. Kiekvienas organas paprastai yra pastatytas iš kelių audinių. Audinys yra ląstelių, panašių savo struktūra ir funkcija, rinkinys. Kiekvieno audinio ląstelės turi savo specialybę. Dirbdami pagal savo specialybę, jie prisideda prie viso augalo gyvenimo, kurį sudaro derinys ir sąveika skirtingi tipai įvairių ląstelių, organų, audinių darbas.
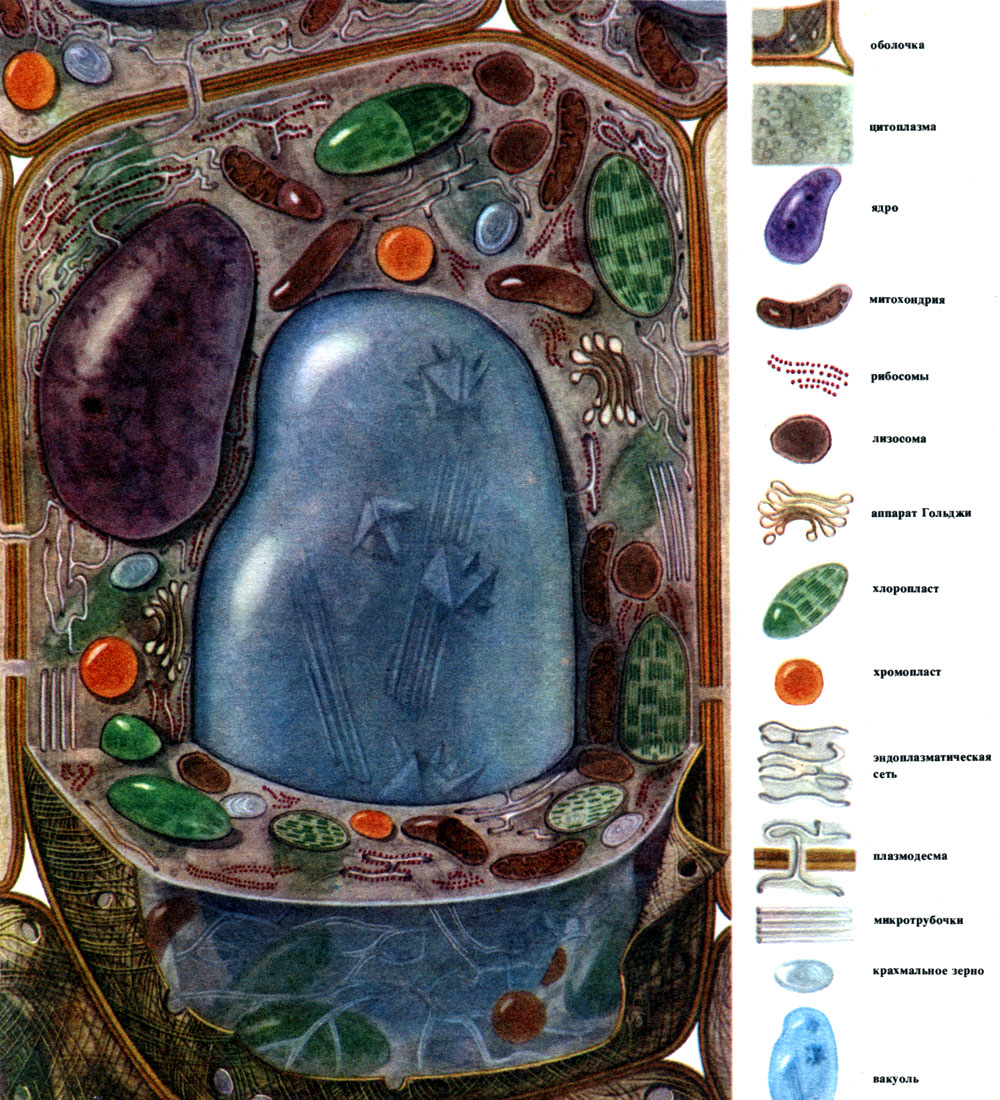
Pagrindiniai, dažniausiai pasitaikantys ląstelių komponentai yra branduolys, citoplazma su daugybe įvairių struktūrų ir funkcijų organelių, apvalkalas ir vakuolė. Membrana dengia ląstelės išorę, po ja yra citoplazma, joje yra branduolys ir viena ar daugiau vakuolių. Tiek skirtingų audinių ląstelių struktūra, tiek savybės labai skiriasi dėl jų skirtingos specializacijos. Išvardyti pagrindiniai komponentai ir organeliai, kurie bus aptariami toliau, juose yra nevienodai vystomi, turi nevienodą struktūrą ir kartais vieno ar kito komponento gali visiškai nebūti.

Pagrindinės audinių grupės, iš kurių yra pastatyti aukštesnio augalo vegetatyviniai (tiesiogiai nesusiję su reprodukcija) organai, yra šios: vientisas, pagrindinis, mechaninis, laidus, šalinamasis, meristematinis. Kiekvienai grupei paprastai priklauso keli panašios specializacijos audiniai, tačiau kiekvienas jų sukonstruotas savaip iš tam tikro tipo ląstelių. Organų audiniai nėra izoliuoti vienas nuo kito, bet sudaro audinių sistemas, kuriose keičiasi atskirų audinių elementai. Taigi, mediena yra mechaninio ir laidžio, o kartais ir pagrindinio audinio, sistema.


Paimkime, pavyzdžiui, du aukštesnio augalo organus - lapą ir stiebą. Ir pamatysime, kiek įvairios struktūros ir funkcijos yra audinius formuojančios ląstelės (22 ir 23 pav.). Taip pat pamatysime, kaip kiekvieno audinio ląstelių struktūra pritaikoma jų specialiųjų funkcijų vykdymui.


Dengiantys audiniai - epidermis, kamštis, pluta - apsaugo augalų organus nuo neigiamo poveikio: nuo išdžiūvimo, perkaitimo, hipotermijos, spinduliavimo energijos, mechaninių pažeidimų, per didelio drėkinimo, nuo svetimų organizmų, pavyzdžiui, bakterijų, prasiskverbimo. Epidermis dengia tiek lapų paviršius, tiek jaunus ūglius, tiek žiedlapius. Paprastai tai yra vienas ląstelių sluoksnis. Tos ląstelių sienos, kuriomis jos liečiasi, paprastai yra vingiuotos, o vienos iškyša užpildo kaimyninės įgaubtą. Tai skatina tvirtą ląstelių ryšį, dėl kurio susidaro vienas tankus sluoksnis. Išorinės ląstelių sienos, besiribojančios su oru, yra storesnės.

Epidermio ląstelės išskiria medžiagas, kurios ant jų nusėda lauke plėvelės pavidalu (pavyzdžiui, kutinas, kuris suformuoja tankią odelę, ir vaškas; kutinas taip pat prasiskverbia į išorines epidermio ląstelių sienas). Storas išorinis apvalkalas ir išorinė plėvelė iš cutino ir vaško suteikia izoliacines, apsaugines šio audinio savybes. Per jo sluoksnį sunku prasiskverbti į dujas, vandenį, bakterijas. Tačiau oras būtinai turi patekti į žaliųjų augalo dalių vidų, o vandens garai ir deguonis išeina. Dujų, taip pat vandens garų, mainai tarp atmosferos ir vidinių jaunų organų audinių, padengtų epidermiu, vyksta stomatų pagalba.
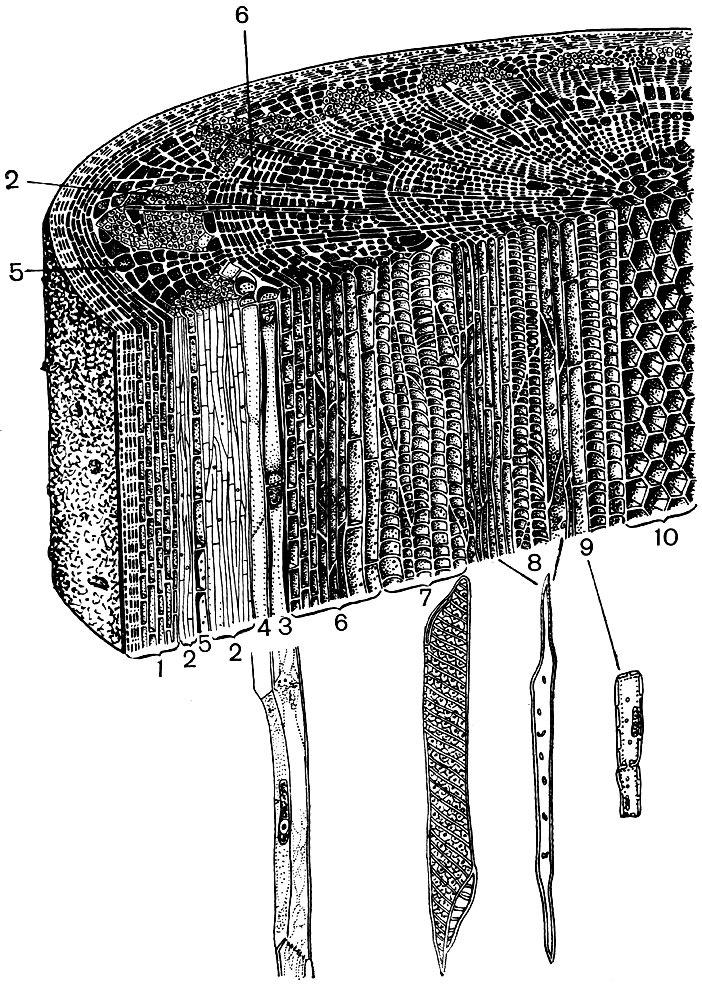
Stomas formuoja specialios epidermio ląstelės, tarp kurių dviejų (vadinamosios stomatų apsauginės ląstelės) yra reguliuojamo dydžio anga - stomatalo tarpas. Per jį atliekamas ryšys tarp atmosferos ir ore esančių tarpląstelinių lapų ar kitų organų, kuriuos padengia epidermis, storio erdvių. Stomatų uždaromos ląstelės, veikiamos drėgmės ar šviesos sąlygų pokyčių, keičia savo formą, užsidaro viena su kita arba atsiveria. Tai darydami jie atsiveria arba, atitinkamai, uždaro stomatalinį tarpą. Šviesoje, kai augalas fotosintezės ir jam reikia anglies dioksido antplūdžio iš atmosferos, stomatalinės spragos yra atviros. Jie užsidaro naktį; apsauginės ląstelės karštuoju paros metu uždaro stomatų spragas, kurios apsaugo augalą nuo didelio vandens praradimo, nuo suvytimo.
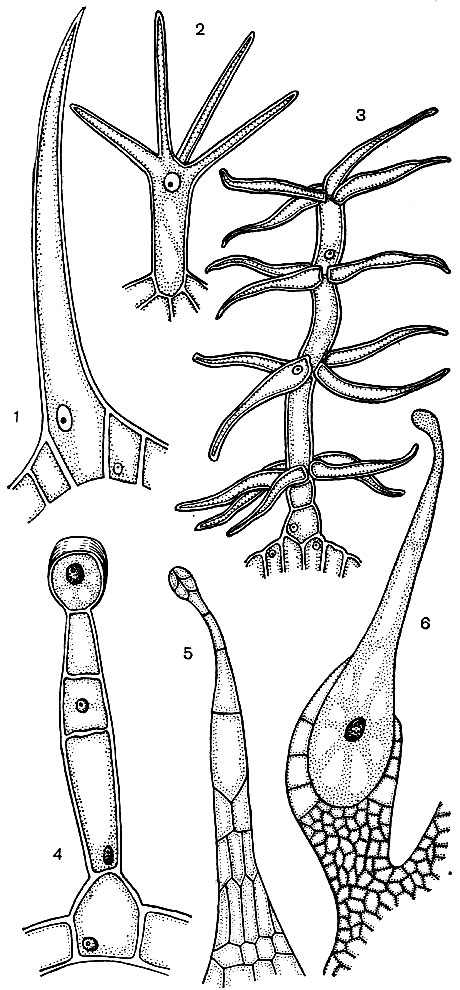
Epidermio ląstelės dažnai formuoja ataugas - plaukelius (24 pav.). Kartais tai yra daugialąsčiai dariniai, kitais atvejais kiekvienas plaukelis yra vienos ląstelės procesas, gulintis bendrame epidermio sluoksnyje. Jie atlieka apsauginį, palaikantį (pavyzdžiui, vijoklinius ar šliaužiančius augalus) išskyrimo vaidmenį. Svarbūs šaknies plaukeliai - šaknies epidermio ląstelių vamzdiniai ataugos. Jie padidina pastarųjų įsiurbimo paviršių. Manoma, kad 14 milijonų šoninių vieno rugių augalo šaknų išauga iki 14 milijardų šaknų plaukelių. Šaknų paviršiaus plotas yra 232 m 2, o plaukų - 400 m 2. Daugelio sėklų ar vaisių epidermis suformuoja plaukelius, kurie skatina sėklų, taigi ir augalų, sklaidą. Dažnai žmogus naudoja plaukus. To pavyzdys yra medvilnė, kuri yra medvilnės sėklų epidermio plaukai; jos plaukai turi storus celiuliozės apvalkalus ir siekia 70 mm ilgį.
Daugiamečių augalų stiebe po epidermiu, norint jį pakeisti, išsivysto grubesnis apsauginis audinys - kamštis. Jo ląstelės miršta, praranda protoplastą ir susideda tik iš storų sienų, kurios supa ertmę, užpildytą oru ar dervinėmis medžiagomis. Sienos yra įmirkytos suberinu, todėl jos yra nepralaidžios vandeniui ir sandarios, šilumą izoliuojančios, taip pat tvirtos ir atsparios. Jie taip pat gali ligifikuoti. Šio audinio pavyzdys yra beržo žievė. Jis yra gana plonas, tačiau kamštinio ąžuolo kamščio storis siekia kelias dešimtis centimetrų. Kamštį kai kuriose vietose pertraukia lęšiai - kito audinio, kuris yra laidus vandeniui ir orui, dariniai. Per lęšius vidinės bagažinės dalys keičiamos su aplinka. Lęšiai vystosi ties stomata.
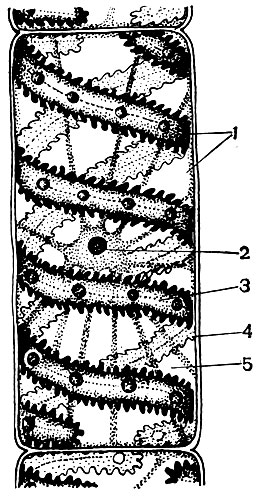
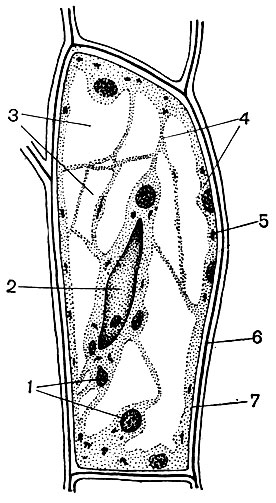
Bastinės ląstelės yra kamiene po vidiniais audiniais. Bastas yra kelių audinių elementų sistema: laidus, atraminis, pagrindinis. Svarbiausias bastelio elementas yra sieto vamzdeliai. Jie pastatyti iš pailgų ląstelių, pailgos išilgai stiebo, sujungtos viena su kita galuose. Tai yra gyvos, bet be branduolio ląstelės, kurių citoplazmos elementai yra šalia sienos. Šių ląstelių sandūroje esantys lukštai turi daug skylių, todėl pertvaros yra tarsi sietas. Dėl to gretimos ląstelės bendrauja tarpusavyje ir taip pasirodo sujungtos ilguose vamzdeliuose, besidriekiančiuose per lapų gyslas ir lapkočius, išilgai stiebo ir šaknies.
Per sietelių mėgintuvėlių sistemą žaliose augalų dalyse susiformavę produktai pereina į visas kitas jo dalis ir juos maitina. Pagrindinis gabenamas produktas yra sacharozė. Sietų vamzdeliai yra laidaus audinio elementas. Šalia sietelio vamzdelių ląstelių yra palydovinės ląstelės. Juose yra branduolių, o jų protoplastas turi tiesioginius ryšius su nebranduolinių sietinių ląstelių citoplazma. Bastoje taip pat yra sričių, susidedančių iš parenchiminių ląstelių (t. Y. Ląstelių, kurių ilgis, plotis ir aukštis yra artimi dydžiams). Juose nusėda įvairios medžiagos - krakmolas, aliejai, dervos. Tai yra pagrindinio audinio ląstelės.
Be to, bastoje yra atraminio audinio elementų - kailio pluoštų. Tai labai ilgos ląstelės su sustorėjusiomis sienomis; ląstelės ilgis tūkstančius kartų gali viršyti jos plotį. Linuose plonų, pailgų kailinių pluoštų ilgis siekia 4 cm, o ramių - net 35 cm. Paprastai tai yra negyvos ląstelės, neturinčios protoplasto. Jie atlieka mechaninę funkciją, sukuria stiebo stiprumą.
Dauguma specializuotų ląstelių nesugeba daugintis. Tačiau augalas auga visą savo gyvenimą, o per visą gyvenimą jame susidaro naujos ląstelės. Jie vystosi iš edukacinių - meristematinių - audinių ląstelių. Reprodukcija dalijant yra meristematinių ląstelių specializacija, jų funkcija organizme. Iš jų atsiradusios ląstelės vystosi, virsta vienomis ar kitomis specializuotomis ląstelėmis - sietelių vamzdelių, kamštinių ląstelių, pluoštų ir kt. Ląstelėmis. Mokomasis audinys yra skirtingose \u200b\u200baugalo dalyse (pavyzdžiui, augimo vietose - ūglių, šaknų viršūnėse). Paprastai kamieno storyje yra keli mokomojo audinio sluoksniai. Po kamštienos sluoksniu yra filogeno ląstelės, kurios, dalydamosi, papildo kamštienos ir plutos ląstelių skaičių. Kastio ląstelių sluoksnis yra basto viduje. Iš jų generuojamų ląstelių, susidarančių už kambio ribų, išsivysto bastinės ląstelės (25 pav.). Patekę į vidų, tai yra arčiau kamieno ašies, atsiranda medienos ląstelės.
Mediena susideda iš laidžių, atraminių ir pagrindinių audinių elementų. Pirmieji apima medienos pluoštus - ilgas negyvas ląsteles su ligifikuotomis sienelėmis; antrajam - indai, kurie yra daugelio ląstelių susiliejimo rezultatas; į trečią - sumedėjusios parenchimos ląstelės. Ląstelės, kurios skiriasi nuo kambialinių elementų į kraujagyslių ląsteles (indus), auga ilgio ir pločio. Jų šoninės sienos yra sustorėjusios ir ligifikuojamos. Tačiau sienose lieka poros, kurias uždaro tik plonas apvalkalas. Gretimų ląstelių sandūrose pertvaros išnyksta, protoplastas miršta. Dėl to susidaro ilgi indai, susidedantys iš kai kurių sienų. Laidžioji sistema tęsiasi per šaknį ir stiebą į lapą. Per tokius indus nuo šaknų iki visų augalo organų vyksta kylantis vandens ir jame ištirpusių druskų srautas.
Kamieno centre yra pagrindinės ląstelės - apvalios arba daugiakampės parenchimo ląstelės. Tai yra pagrindinio audinio elementai. Kartais jie yra tuščiaviduriai ir juose yra oro; kartais jie užpildomi atsarginėmis maistinėmis medžiagomis, įvairiais kristalais, taninais. Jų sienas galima susieti. Radialiniai spinduliai prasiskverbia į medieną ir bastą. Jų ląstelės yra kambio dariniai ir turi saugojimo funkciją. Basto lygyje šie spinduliai pastebimai išsiplečia.
Lapų minkštime, tarp viršutinio ir apatinio epidermio, yra pagrindinis audinys - ląstelės su plonomis membranomis ir daug žalių plastidų - chloroplastai. Šiose ląstelėse vyksta fotosintezė. Viršutiniai sluoksniai susideda daugiausia iš pailgų ląstelių, glaudžiai greta vienas kito - šios koloninės parenchimos. Apatiniai sluoksniai atsilaisvina tarp ląstelių, yra tarpląstelinės erdvės - erdvės, užpildytos oru; tai puri parenchima. Į parenchimą prasiskverbia išsišakojusios venos, kurios yra pastatytos iš mechaninių (įvairių pluoštų) ir laidžių (sietų vamzdelių ir vandeningųjų sluoksnių) audinių ryšulių.
![]()
Šis trumpas ir schematiškas stiebo, lapo struktūros plano pateikimas parodo, kaip skiriasi to paties augalo ląstelių dydis, forma, struktūra, funkcijos. Jei paimsite skirtingus augalus, galite susitikti su dar didesne ląstelių įvairove. Tačiau to paties pavadinimo audinių ląstelės, net ir augaluose, kurie yra labai nutolę vienas nuo kito, paprastai yra panašūs, nes šios ląstelės atlieka panašias funkcijas. Beržo lapo epidermio ląstelės ir, pavyzdžiui, kiaulpienės yra panašesnės viena į kitą nei kiaulpienės ir jos sietinės ląstelės epidermio ląstelės. Išskirtiniai ląstelės bruožai pirmiausia siejami su ypatinga jos funkcija. Ląstelės, kurių specializacija yra mechaninė funkcija, turi sustorėjusias ir dažnai ligifikuojamas membranas. Fotosintezės specializacija lemia chloroplastų atsiradimą ląstelėse. Laidavimo funkcijos specializacija siejama su ląstelių pailgėjimu, protoplasto praradimu, ląstelių membranų pokyčiais sandūrose, dėl kurių bendrauja gretimų ląstelių ertmės. Ląstelėms, kurių specializacija yra apsauginė funkcija, būdingi įvairūs išorinių sienų pokyčiai, plaukelių buvimas ir gebėjimas gaminti apsaugines medžiagas.
Specializuojantis maistinių medžiagų kaupimo funkcijai gali padidėti ląstelių dydis, atsirasti jose labai didelės vakuolės. Meristematinių, intensyviai besidalijančių ląstelėse yra ypač išvystytos tos tarpląstelinės struktūros, kurios užtikrina medžiagų - sudedamųjų ląstelės dalių - sintezę.
Atsižvelgiant į specializacijos pobūdį, kai kurie ląstelės struktūros ir darbo ypatumai yra labai stipriai išvystyti, o kiti, priešingai, yra mažai išvystyti arba jų nėra visiškai. Kaip matėme, kai kurios ląstelės, tapusios specializuotomis, žūsta ir yra negyvos būsenos, kai jos pradeda atlikti savo ypatingą funkciją daugialąsčiame organizme (kamštinėse ląstelėse, medienos pluoštuose, induose).
Daugialąsčių gijinių dumblių ir vienaląsčių augalų ląstelės yra labai įvairios ir savitos (26 ir 27 pav.). Be to, bet kurio pastarojo ląstelė labai skiriasi nuo daugialąsčių augalų ląstelių. Ji viena turi atlikti keletą funkcijų, kurios daugialąsčiuose augaluose yra padalintos tarp skirtingų audinių ląstelių.
Tuo pačiu metu net labai skirtingos ląstelės turi gilų struktūros ir funkcijos panašumą. Daugialąsčiams organizmams tai, visų pirma, yra dėl to, kad visos kūno ląstelės (jei neįtrauksime vegetatyvinės reprodukcijos, kurioje visas daugialąsčio kompleksas iškart atskiriamas nuo motininio augalo kaip naujas individas ar jo rudimentas) yra tos pačios pirmtakinės ląstelės palikuonys. Todėl, kad ir kokios specializuotos būtų ląstelės, jos turi bendrą kilmę, todėl yra giminės.
Antra, bendri bruožai skirtingų rūšių augalų ląstelių struktūroje siejami su tuo, kad visi augalai yra susiję vienokiu ar kitokiu laipsniu. Visi augalai evoliucijos būdu išsivystė iš bendrų vienaląsčių protėvių. Bendrus bruožus šiuolaikinių augalų ląstelės paveldi iš senovės kamieninių ląstelių. Tai taip pat siejama su bendromis augalų ir gyvūnų ląstelių struktūros ir darbo ypatybėmis.
Trečia, panašumas yra dėl to, kad visos gyvos ląstelės, nesvarbu, kokią ypatingą funkciją jos atlieka organizme, pirmiausia turi užtikrinti savo gyvenimą. Ląstelės pasisavina maistines medžiagas, jas apdoroja, įgyja energijos ir kuria savo kūną, kvėpuoja, atsikrato nereikalingų medžiagų, kovoja su įvairia žala, reaguoja į išorinių sąlygų pokyčius, atstatydamos savo gyvybinę veiklą ir auga. Visi šie procesai skirtingose \u200b\u200bląstelėse vykdomi panašiu būdu ir pasitelkiant to paties tipo struktūras, kurios pagal struktūrą yra bendros ne tik skirtingoms augalų, bet ir augalų bei gyvūnų ląstelėms. Reikia pasakyti, kad bet kurios specialios ląstelės funkcijos atlikimas yra pagrįstas jos bendromis savybėmis. Viena ar kita savybė, gebėjimas, būdingas visoms ląstelėms, ypač stipriai vystosi specializuotoje ląstelėje ir užtikrina, kad ląstelė atliktų pagrindinę, specialią funkciją. Tie bendri bruožai, be kurių neįmanoma atlikti šios ypatingos funkcijos, yra išsaugoti ląstelėje, o likusi dalis gali būti prarasta. Negyvos specializuotos ląstelės yra kraštutinis to atvejis. Ypatinga tokių ląstelių funkcija yra susijusi su jų membrana; protoplastas reikalingas tik tol, kol jis sukuria apvalkalą; po to jis miršta, o visa ląstelė susideda tik iš negyvo apvalkalo, kuris tinka augalo reikmėms.
Atsižvelgiant į bendrąsias augalų ląstelių struktūros ir gyvybinės veiklos ypatybes, patogu kalbėti apie tam tikrą tipinę ląstelę, kuri absorbavo bendrus įvairių specializuotų ląstelių bruožus. Tokią ląstelę, nors jos nėra „gryna forma“, galima net pavaizduoti (7 lentelė, 28 pav.). Iš tikrųjų ląstelių jie labiau panašūs į tokias „tipines“ lapų parenchimos ląsteles (8 lentelė, 29 pav.).
Pereikime prie augalų ląstelės struktūros ir darbo aprašymo.
5 lentelė. Angiosperms: viršuje kairėje - magnolijos žiedas; viršuje dešinėje - obuolių žiedai; žemiau yra lotosas.
6 lentelė. Angiosperms: viršuje (iš kairės į dešinę) - varpai, pelargoniumas, šafranas (krokas); apačioje kairėje - astrai; apačioje dešinėje - varnalėša.
Lauke augalo ląstelė yra padengta membrana, kurios storis ir struktūra skirtingose \u200b\u200bląstelėse nėra vienodi. Jį formuojančios medžiagos gaminamos citoplazmoje ir nusėda už jos ribų, palaipsniui sukuriant apvalkalą. Šios medžiagos pirmiausia yra didelių molekulių polisacharidai - pektinas, hemiceliuliozė ir celiuliozė nedideliais kiekiais. Jie sudaro vadinamąjį pirminį apvalkalą. Ji yra gana elastinga, ląstelei augant, ji išsitempia ir taip pat auga, todėl netrukdo ląstelių augimui. Tačiau tai sukuria tam tikrą ląstelės stiprumą ir sugeba apsaugoti ją nuo mechaninių pažeidimų. Yra ląstelių, kurioms trūksta tokios pirminės membranos - tai yra kai kurios ląstelės, kurios naudojamos nelytiniam ir lytiniam dauginimuisi (dumblių ir žemesnių grybų zoosporos ir lytinės ląstelės, aukštesnių augalų vyriškosios lyties ląstelės). Daugelis ląstelių turi ne tik pirminę, bet ir antrinę membraną. Jis susidaro po pirminiu ir yra pastatytas daugiausia iš celiuliozės. Celiuliozė yra polisacharidas, kurio molekulės sudaro geriausius siūlus - mikrofibrilius. Korpuse celiuliozės gijos panardinamos į amorfinę medžiagą, susidedančią iš pektino junginių. Kai kuriose ląstelėse šios celiuliozės mikrofibrilės yra išsidėsčiusios per visą ląstelės ilgį žiedais; dėl to tokios ląstelės gali ištiesti ilgį, bet ne į plotį (pavyzdžiui, kamieno indų ląstelės). Kiti turi siūlus išilgai; tokios membranos turinčios ląstelės yra ištemptos skersai, bet išilgai labai tvirtos. Kituose jie išsidėstę įstrižai, formuodami spiralę (medvilnės sėklų epidermio plaukeliai, kailinių pluoštų). Visa tai primena gelžbetonines konstrukcijas, o celiuliozės siūlai čia atlieka geležinių strypų, o pektino medžiagos - cemento vaidmenį. Ląstelės su antrine membrana yra labai patvarios. Jie sudaro mechaninius augalo atraminius audinius. Kartais antrinė membrana atlieka ir maistingų produktų sandėlio vaidmenį: ją sudarančios medžiagos gali būti paverstos kitomis, paprastesnėmis, kurios vartojamos kaip maistas.
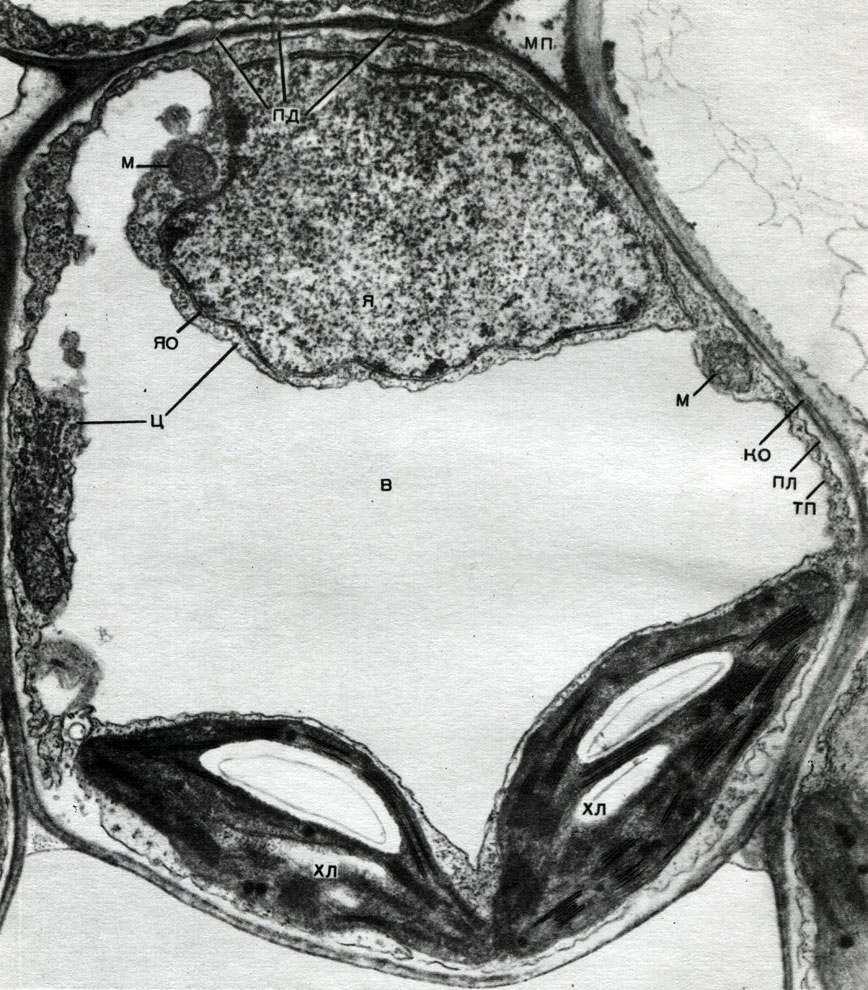
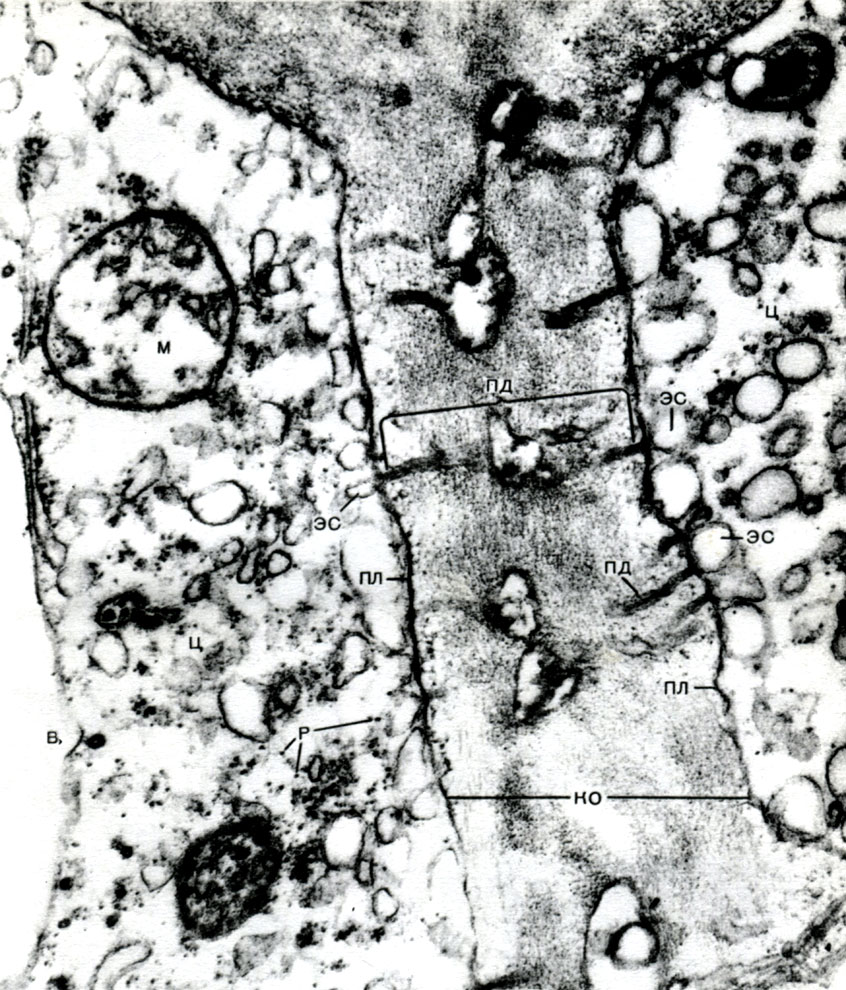
Korpuse yra nesutirštėjusios vietos - poros (pirminiame apvalkale jos vadinamos urvų laukais), per kurias vyksta ryšys tarp kaimyninių ląstelių. Ploni citoplazmos virvelės praeina per porų laukus ir poras. Tai plazmodesmatos, surišančios kaimyninių ląstelių citoplazmą (9 lentelė). Jie keičiasi medžiagomis tarp kaimyninių ląstelių. Plasmodesmata kartu su laidžių audinių elementais sujungia kūno ląsteles ir audinius į vieną visumą.
Keitimasis medžiagomis ir jaudulio sklidimas leidžia ląstelėms daryti įtaką vienas kito vystymuisi ir darbui, o kiekvienas audinys veikia visų kitų audinių gyvybines funkcijas. Tai sukuria visų vieno organizmo dalių, viso augalo, darbo koordinavimą.
Daugelyje ląstelių ląstelių membrana su amžiumi yra įmirkyta medžiagomis, kurios dar labiau ją sustiprina. Lignino kaupimasis jame lemia apvalkalo lignifikaciją. Medienos ląstelių ir dažnai kailinių pluoštų sienos, sėklos žievės ląstelių lukštai, kartais perikarpas (30 pav.) Ir net žiedlapiai tampa lignifikuoti. Ląstelės su ligifikuotomis sienelėmis yra ne tik stipresnės, bet ir mažiau laidžios mikrobams ir vandeniui. Kai kurių kitų ląstelių membranos - vientisieji audiniai, taip pat ant žaizdų paviršių - tampa kamštienos, mirkomos suberine. Tada ląstelių turinys išnyksta, nes suberinas nelaidus vandeniui ir dujoms, tačiau tokių negyvų ląstelių audinys apsaugo vidines gyvąsias augalo dalis nuo kenksmingų išorinių poveikių. Kaip jau minėta, išorinės epidermio ląstelių membranos paprastai yra impregnuotos arba padengtos kutinu ir vašku, ir tai apsaugo ląsteles nuo pernelyg garuojančio vandens, nuo infekcijos. Žarną taip pat galima įmirkyti mineralais, daugiausia kalcio druskomis.
Tačiau ląstelių gyvenime yra atvejų, kai jų membrana turėtų tapti mažiau patvari nei anksčiau. Tai atsitinka, pavyzdžiui, sėklų daiginimo metu.
Polisacharidų ląstelių membrana yra būdingas augalo ląstelės struktūros bruožas, kuris ją išskiria iš gyvūnų ląstelės.
Membrana arba ląstelės sienelė yra apsauginis darinys. Po membrana yra citoplazma. Jo išorinis sluoksnis, esantis greta membranos, yra paviršiaus ląstelės membrana - plazmalema. Tai yra į riebalus panašių ir baltymų molekulių sluoksnių derinys. Tokios membranos vadinamos lipoproteinų membranomis („lipos“ - riebalai, „baltymai“ - baltymai). Tokio dizaino membrana atriboja citoplazmą nuo vakuolių; ši membrana vadinama tonoplastu. Daugelis ląstelių organelių yra pastatyti iš lipoproteinų membranų. Tačiau kiekvienu atveju membrana yra pastatyta iš į riebalus panašių medžiagų (lipidų) ir baltymų, būdingų būtent šiai membranai. Kokybinė lipidų ir ypač baltymų įvairovė yra milžiniška, todėl didžiulė membranų įvairovė skiriasi savybėmis tiek toje pačioje ląstelėje, tiek skirtingose \u200b\u200bląstelėse.
Plazmalema reguliuoja medžiagų patekimą į ląstelę ir jų išėjimą iš jos, užtikrina selektyvų medžiagų įsiskverbimą į ląstelę ir iš jos. Skirtingų medžiagų prasiskverbimo per membraną greitis yra skirtingas. Pro jį gerai prasiskverbia vanduo ir dujinės medžiagos. Riebaluose tirpios medžiagos taip pat lengvai prasiskverbia, tikriausiai dėl to, kad joje yra lipidų sluoksnis. Daroma prielaida, kad membranos lipidinis sluoksnis yra prasiskverbęs poromis. Tai leidžia riebaluose netirpstančioms medžiagoms prasiskverbti pro membraną. Poros turi elektrinį krūvį, todėl jonų prasiskverbimas per jas nėra visiškai laisvas. Tam tikromis sąlygomis porų krūvis keičiasi, ir tai reguliuoja membranų pralaidumą jonams. Tačiau membrana nevienodai pralaidi skirtingiems jonams, turintiems tą patį krūvį, ir skirtingoms, nepakrautoms, artimo dydžio molekulėms. Tai pasireiškia svarbiausia membranos savybe - jos pralaidumo selektyvumu: vienoms molekulėms ir jonams ji yra geriau praleidžiama, kitoms - blogiau.
Medžiaga juda į ląstelę, difunduodama į ją, jei jos koncentracija lauke yra didesnė už koncentraciją viduje. Priešingu atveju jis turėtų pasklisti iš ląstelės. Bet, patekusios į ląstelę, šios medžiagos molekulės gali iškart dalyvauti medžiagų apykaitoje ir transformuotis į kitas medžiagas. Tada šios medžiagos koncentracija ląstelės viduje vėl sumažėja, o nauja jo molekulių dalis difunduoja į ląstelę. Dėl to ląstelės, kurios aktyviai vartoja tą ar tą medžiagą, pastaroji intensyviai patenka iš aplinkinių ląstelių.
Kitas ląstelės sustiprintos medžiagos absorbcijos mechanizmas yra jos prisijungimas prie baltymų ar kitų medžiagų. Jungdamasi medžiaga palieka viduląstelinį tirpalą ir netrukdo tolesnei tos pačios medžiagos difuzijai. Jis toliau patenka į ląstelę ir joje susikaupia susieta forma. Taigi kai kurių jūros dumblių ląstelėse jodo koncentracija milijonus kartų viršija jo koncentraciją jūros vandenyje.
Visais šiais atvejais kalbame apie medžiagų judėjimą difuzijos būdu. Tačiau ląstelių membranos taip pat turi mechanizmus, perkeliančius medžiagas per save iš mažesnės koncentracijos į didesnę. Tarp membranos baltymų yra baltyminių medžiagų, kurių darbas yra įvairių medžiagų perkėlimas iš vienos membranos pusės į kitą. Jie vadinami transporto fermentais. Tikriausiai toks transportinis fermentas su tarpine medžiaga sudaro tarpinį produktą; pastarasis praeina per membraną, tada suskaidomas į pradines dalis, po kurių fermentas grįžta atgal, o pernešta medžiaga lieka kitoje membranos pusėje. Šio proceso detalės nėra žinomos, tačiau pats tokio aktyvaus medžiagų gabenimo faktas nekelia abejonių. Skirtingai nei difuzinis, pasyvus, fermentinis transportas reikalauja energijos sąnaudų. Tikriausiai būtent dėl \u200b\u200bšio aktyvaus jonų pernešimo šaknies epidermio ląstelės sugeba absorbuoti iš dirvožemio augalui reikalingas neorganines medžiagas ir paskui pernešti jas per augalą iš ląstelės į ląstelę.
Chemiškai ar fiziškai pasikeitus išorinei ir vidinei aplinkai, ląstelių membranos keičia jų pralaidumą, taip pat jos selektyvumo laipsnį ir pobūdį. Tuo remiasi medžiagų judėjimo į ląstelę ir iš jos reguliavimo mechanizmai. Maistinių medžiagų membranų pralaidumo pokytis atsispindi ląstelės medžiagų apykaitos procesų intensyvume, joje vykstančių sintezių pobūdyje ir per visą jos gyvenimą. Natrio ir kalio jonų membranų pralaidumo pokytis yra ląstelių sužadinimo sklidimo mechanizmas - biologinės srovės atsiradimas ir judėjimas. Prie membranos paviršiaus pritvirtintas didelis skaičius katijonų, daugiausia kalio jonų. Todėl jis neša teigiamą krūvį iš išorės. Veikiant dirgikliams, kalio jonai yra atskirti nuo paveiktos zonos, srities krūvis sumažėja ir jis tampa elektronegatyvus kaimyninių membranos sričių atžvilgiu. Šis elektronegatyvumas savo ruožtu dirgina kaimynines sritis, tuo pačiu pašalindamas jų krūvį, depoliarizuodamas juos. Elektronegatyvumo banga plinta per membraną - tai biologinė srovė. Tada kalio jonai vėl sėdi ant membranos, suteikdami jai pradinį krūvį - po depoliarizacijos bangos seka atkūrimo banga. Tokios membranos taip pat suformuoja plazmodesmatos paviršius, praeinančius per ląstelių sienas ir sujungiančius kaimyninių ląstelių citoplazmą. Biologinė srovė taip pat juda išilgai jų, plinta iš ląstelės į ląstelę. Augalų ląstelių biologinės srovės vis dar menkai suprantamos. Tačiau akivaizdu, kad jie yra signalo metodas, naudojamas augalų ląstelėje, siekiant sukelti kai kurias chemines reakcijas, o kitas - slopinti. Faktas yra tas, kad visi cheminiai procesai, vykstantys ląstelėje ir sudarantys jos gyvybinės veiklos esmę, atliekami dalyvaujant biologiniams katalizatoriams - fermentams. Kiekviena reakcija įmanoma tik tada, kai ją suaktyvinantis fermentas yra aktyvus. Dauguma fermentų suaktyvėja veikiant tam tikriems neorganiniams katijonams: K., Na. , Ca .., Mg .., Mn ... Biosrovė, keisdama įelektrintų medžiagų membranų pralaidumą, sukuria sąlygas joms prasiskverbti per membranas ir kontaktuoti su fermentais. Taigi, biologinė srovė suaktyvina tam tikrus fermentus ir tokiu būdu reguliuoja bei nukreipia medžiagų apykaitą ląstelėje. Medžiagų apykaitos reguliavimas biologinėmis srovėmis yra tik vienas iš daugelio būdų reguliuoti viduląstelinę medžiagų apykaitą.
Tai, kas buvo pasakyta apie ląstelės paviršiaus membranos - plazmalemos - pralaidumą, taip pat taikoma ir kitoms tarpląstelinėms membranoms, įskaitant tas, kuriose pastatyta daug ląstelių organelių.
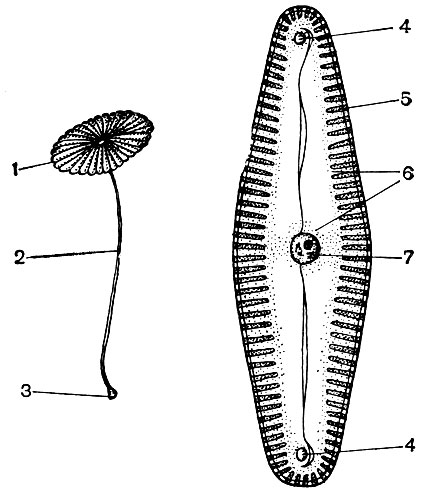
Citoplazma, kadaise laikyta homogenišku koloidiniu baltymų medžiagų tirpalu, iš tikrųjų yra sudėtingai sudaryta. Plėtojant mikroskopinius tyrimo metodus, buvo išaiškintos vis subtilesnės citoplazmos struktūros detalės. Citoplazmoje buvo atrasti įvairūs organeliai (organeliai) - struktūros, kurių kiekviena atlieka tam tikras fiziologines ir biochemines funkcijas. Svarbiausi citoplazmos organeliai yra mitochondrijos, endoplazminis tinklas (endoplazminis tinklas), Golgi aparatas, ribosomos, plastidės, lizosomos. Judrios ląstelės (dumblių zoosporos ir gametos, asiūklių spermatozoidai, paparčiai, cikladai, kai kurie vienaląsčiai ir kolonijiniai dumbliai) turi judėjimo organelius - vėliavėles.
Ypač daug naujų faktų apie smulkią citoplazmos struktūrą atnešė ir atneša elektroninį mikroskopą, kuris leidžia ištirti pačių organelių struktūros detales. Šiuolaikiniai biofizikiniai ir biocheminiai metodai leidžia atskirti tam tikrus citoplazmos organelius gryna forma ir juos tirti cheminė sudėtis ir jų funkcijos. Už ląstelės ribų, sudėtingos sudėties aplinkoje, daugelis organelių sugeba atlikti darbą, kurį jie daro būdami ląstelėje.
Citoplazmos dalis, į kurią paniro organeliai ir kuri iki šiol atrodo be struktūros, vadinama pagrindine citoplazmos arba hialoplazmos medžiaga. Hyalo plazma anaiptol nėra pasyvus užpildas, o aktyviai veikianti citoplazmos dalis. Jame vyksta nemažai gyvybiškai svarbių cheminių procesų, jame yra daug fermentinių baltymų, kurių pagalba šie procesai ir atliekami.
Ląstelių gyvenimas susideda iš nuolatinio cheminio darbo, kuris visumoje vadinamas metabolizmu. Iš esmės ląstelė yra chemijos gamykla, gaminanti platų produktų asortimentą ir nepriklausomai išgaunanti jai pagaminti reikalingą energiją. Jos produktai yra medžiagos, kurių jai pačiai reikia palaikyti savo gyvenimą (auginant ir vystantis kurti savo kūną, pakeisti susidėvėjusias dalis) ir reprodukcijos metu kurti dukterines ląsteles bei kitų kūno ląstelių poreikius.
Visas ląstelėje vykstančias chemines reakcijas galima suskirstyti į dvi grupes. Todėl kai kurios iš tų ar tų medžiagų suskaidomos į mažesnes molekules. Dėl kitų mažų molekulių medžiagų sintetinamos didesnių molekulių medžiagos. Bet kurios medžiagos molekulė susideda iš atomų, kurie laikomi kartu cheminėmis jungtimis, tai yra tam tikru koncentruotos potencialios cheminės energijos kiekiu. Išardžius molekulę, ryšiai nutrūksta ir išsiskiria jų cheminė energija. Sintezei, tai yra, norint iš mažų formuoti didesnę molekulę, būtina sukurti naujas chemines jungtis, Būtina į juos įdėti tam tikrą energijos dalį. Priešingai, skilimo reakcijos galiausiai vyksta su energijos išsiskyrimu, nes jų metu nutrūksta ryšiai tarp atomų. Ląstelėje vykstančių skilimo reakcijų biologinė prasmė yra ta, kad jos išskiria cheminę energiją, kurią ląstelė panaudoja sintezės reakcijoms ir kitų rūšių darbams (elektriniams, mechaniniams, medžiagų transportavimo darbams) gaminti. Kadangi ląstelė per visą gyvenimą sintetino įvairias medžiagas, ji turi nuolat skaidyti kitas medžiagas. Išlaisvinta energija iš karto nenaudojama sintezėms. Pirma, jis laikomas susidarant specialioms medžiagoms - cheminės energijos kaupėjams - adenozino trifosforo rūgščiai (ATP) ir panašiems junginiams. Tinkamu laiku ir tinkamame ląstelės taške ATP yra padalijamas ir atiduodamas energija ląstelei reikalingos medžiagos sintezei.
Visos cheminės reakcijos ląstelėje - tiek sintezė, tiek irimas - atliekamos fermentų pagalba. Fermentai yra baltyminės medžiagos, greitinančios reakcijų eigą. Šis pagreitis yra toks didelis, kad be fermentų tokios reakcijos apskritai nebūtų įmanomos ląstelėje. Yra atvejų, kai fermento dėka reakcija pagreitėja 1011 kartų. Tai reiškia, kad reakcija, kuri baigiasi fermento dalyvavimu per 0,01 sek., Be jo, vyktų 31 metus. Akivaizdu, kad tokios reakcijos be fermento būtų tiesiog nerealios.
Be to, fermentų dėka ląstelių cheminių reakcijų eiga yra kontroliuojama ir reguliuojama. Fermentų aktyvumas ląstelėje keičiasi atsižvelgiant į tam tikros medžiagos ar energijos poreikį, tai yra dėl šių fermentų darbo. Kai reikia medžiagos, tie fermentai, kurių dėka ji susidaro, vėl aktyvuojasi arba sintetinami. Medžiagų susidarymas, energijos išsiskyrimas ir kaupimas yra vienas po kito einančių reakcijų rezultatas, visos fermentų grandinės darbo rezultatas. Visos gyvų ląstelės dalių struktūros yra pastatytos būtent iš fermentų ir iš medžiagų, kurios šiuos fermentus laiko kartu. Šiuo atveju fermentai, dalyvaujantys gretimose, nuosekliai vykstančiose reakcijose, yra vienas šalia kito. Jie perneša konvertuojamų medžiagų molekules tarpusavyje tarsi palei konvejerį, ir kiekvienas iš jų atlieka savo molekulės darbo operaciją.
Hialoplazmoje yra fermentų, kurie skaido gliukozės molekules į paprastesnes piruvo rūgšties molekules. Šiuo atveju išsiskirianti energija kaupiama susidarant ATP molekulėms. Tas pats procesas vyksta ir ląstelės branduolyje. Tačiau didžioji energijos dalis gaunama specialiuose citoplazmos organeliuose - mitochondrijose, nes yra gilesnis medžiagų skaidymas.
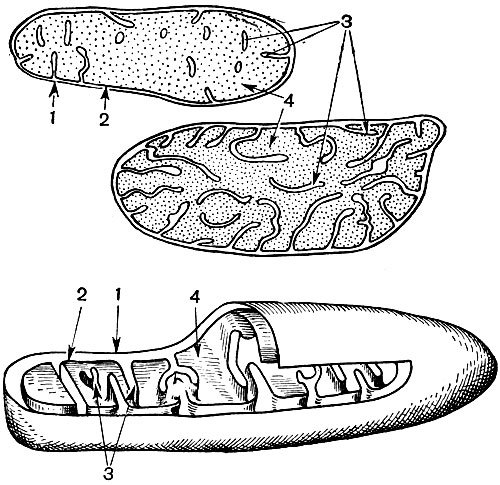
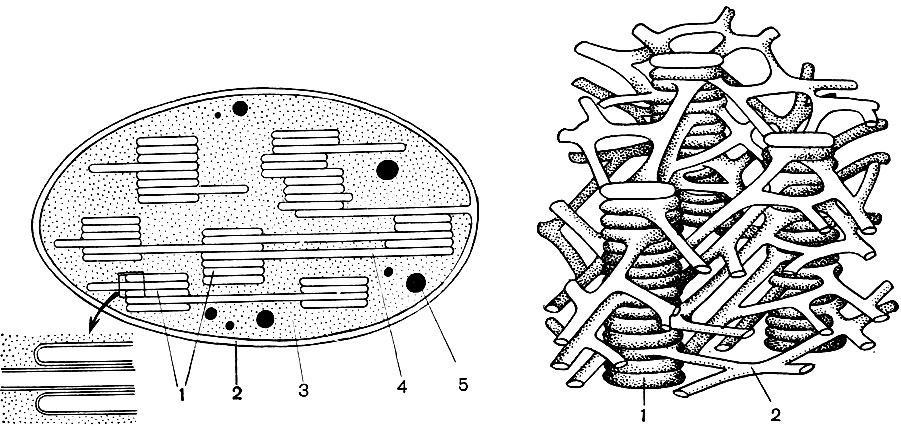
Mitochondrijos yra maži apvalios arba pailgos formos kūnai, 0,5 - 1,5 mikrono dydžio, tai yra, bakterijos dydžio. Jų skaičius ląstelėje paprastai yra didelis, apie 100 - 3000. Tačiau yra ląstelių, kuriose yra nedidelis mitochondrijų skaičius. Taigi, fucus jūros dumblių spermoje yra tik 4 mitochondrijos, o vienaląsčiuose dumbliuose - vienas. Mitochondrijos yra matomos šviesos mikroskopu, tačiau jų smulkią struktūrą galima tirti tik elektroniniu mikroskopu (10 lentelė, struktūros schema - 31 pav.). Mitochondrijos yra dariniai, pastatyti iš lipoproteinų membranų, panardintų į pagrindinę medžiagą - matricą. Mitochondrinę membraną sudaro dvi membranos, tarp kurių yra tarpas.
Vidinė apvalkalo membrana sukelia daugybę invazijų viduje, tai yra cristae. Tarp jų yra matrica. Tiek vidinė mitochondrijų membranos membrana, tiek jos suformuotos krizos yra pastatytos pagal išsidėsčiusių fermentų eiliškumą. Dėl klosčių - cristae, mitochondrijų viduje esantis membranų darbinis paviršius yra labai didelis. Mitochondrijų matricoje, tai yra tarp cristae, randama nemažai fermentų.
Šių fermentų derinys vykdo viduląstelinį kvėpavimą ir kvėpavimo metu išsiskyrusios energijos kaupimą ATP pavidalu. Mitochondrijų darbas yra glaudžiai susijęs su procesais, vykstančiais hialoplazmoje, kur vyksta pirmieji gliukozės ir kitų medžiagų skaidymo į piruvos rūgštį etapai. Mitochondrijose vyksta tolesnis jos skilimas. Piruvinė rūgštis prasiskverbia į mitochondrijas ir čia, žingsnis po žingsnio, ji oksiduojama iki anglies dioksido ir vandens, tuo pačiu metu vartojamas deguonis. Tai yra tarpląstelinis kvėpavimas, kurio metu ląstelė, skaidydama ir oksiduodama medžiagas, gamina daug energijos, kurią vėliau gali panaudoti įvairiems poreikiams tenkinti.
Pirmasis gliukozės molekulės skaidymo etapas, kurio metu ji padalijama per pusę ir vyksta hialoplazmoje, suteikia ląstelei tik dvi ATP molekules.
Dėl antrojo etapo, lemiančio visišką gliukozės „degimą“, susidaro dar 36 ATP molekulės. Todėl mitochondrijos pagal savo funkciją yra ląstelės jėgainės, mašinos, skirtos gauti didžiausią energijos kiekį. Pats gliukozės skilimo produktų skilimas įvyksta mitochondrijų matricoje, o ATP susidaro dėl reakcijų, vykstančių ant jos vidinių membranų, įskaitant kvėpavimo fermentus ir fermentus, užtikrinančius ATP susidarymą. Krizių skaičius mitochondrijose gali skirtis. Kuo jų daugiau, tuo didesnis mitochondrijų biocheminis aktyvumas.
Mes čia kalbame apie gliukozę kaip medžiagą, kurią skaidydama ląstelė gamina energiją. Gliukozė yra pagrindinė, bet ne vienintelė tokia medžiaga. Jo molekulėje yra šešių anglies atomų, sujungtų vienas su kitu, griaučiai. Dėl ilgos ir sudėtingos reakcijų grandinės jo molekulė susmulkinama, oksiduojama ir galiausiai padalijama į šešias neorganinės medžiagos - anglies dioksido (CO 2) molekules, kurių kiekvienoje molekulėje yra tik vienas anglies atomas, ir ji yra labai oksiduota. Susumavus visas paskesnes gliukozės oksidacijos reakcijas ir neįtraukus visų tarpinių produktų, galima gauti bendrą šio proceso reakciją:
Krakmolas lengvai virsta gliukoze, po kurio jis skyla aukščiau aprašytu būdu. Baltymai ir riebalai suteikia įvairių organinių rūgščių, kurios paverčiamos tarpiniais gliukozės skilimo produktais, o vėliau oksiduojamos taip pat, kaip ir pastarosios, ir tų pačių fermentų pagalba.
Visiškas biologinis organinių medžiagų oksidavimas yra panašus į jo degimą. Abiem atvejais rezultatas yra išsiskyręs anglies dioksidas, vanduo ir energija. Tačiau degimo metu ši energija išsiskiria šilumos pavidalu ir visiškai iš karto; vykstant biologinei oksidacijai, cheminių ryšių energija išsiskiria dalimis, o pagrindinė jos dalis yra surišta, pereidama į ATP cheminio fosfatinio ryšio energiją. Todėl ląstelė gauna tokios koncentracijos energijos koncentratą, kuris tada tinkamu laiku ir tinkamu momentu gali būti naudojamas kuriant naujus cheminius ryšius, sintetinant naujas medžiagas, taip pat gaminant kitokio pobūdžio darbus - elektrinius, mechaninius, taip pat atliekant medžiagų transportavimą. iš aplinkos į ląstelę, iš ląstelės į aplinką ir iš ląstelės į ląstelę.
Kuo aktyvesnė ląstelės gyvybinė veikla, tuo didesnis jos energijos poreikis ir tuo daugiau mitochondrijų. Jie gali būti netolygiai pasiskirstę toje pačioje ląstelėje: ląstelės dalyje, kuri šiuo metu dirba aktyviau, jų yra daugiau.
Mitochondrijos sugeba sintetinti kai kurias medžiagas, kurias jos pačios sudaro. Dėl to mitochondrijos gali daugintis.
Endoplazminis tinklas yra citoplazmos organoidas, kuriame sintetinama labai daug medžiagų (10 lentelė). Endoplazminis tinklas yra kanalų sistema, prasiskverbianti į citoplazmą ir kurios vienose vietose siaurėja, kitose plečiasi, susidaro arba cisternos, arba plokšti maišeliai, arba išsišakojantys vamzdeliai. Visų šių darinių sienos yra pastatytos iš membranų, kuriose yra fermentų.
Kaip ir kitose ląstelės membraninėse formacijose, fermentai tinkle yra išdėstyti tvarkingai. Šiuo atveju kaimyniniai fermentai vykdo nuosekliai vykstančias reakcijas (darbo operacijas), o jų grupė vykdo visą reakcijų grandinę, vedančią sukurti tą ar tą medžiagą.
Atskirkite agranulinį (lygų) ir granuliuotą endoplazminį tinklą. Granuliuoto tinklelio kanalų išoriniame paviršiuje yra daugybė mažų organelių - ribosomų, kurių funkcija yra baltymų molekulių sintezė. Agranulinis endoplazminis tinklas, kuris kiekybiškai vyrauja prieš granuliuotą augalų ląstelėje, ribosomų neturi.
Be to, endoplazminis tinklas yra daugelio rūšių fermentinio medžiagų virsmo, daugiausia jų sintezės, konvejeris, taip pat greitkelių sistema, palei kurią medžiagos juda ląstele. Tinklas prasideda nuo išorinės branduolio membranos membranos ir, išsišakodamas, artėja prie įvairių citoplazmos organelių, taip pat prie plazmalemos. Taigi jis sujungia visas ląstelės dalis. Be to, endoplazminio tinklo kanalai praeina pro plazmodesmatą, jungdami kaimyninių ląstelių tinklą.
Be to, endoplazminio tinklo membranos išskiria citoplazmą į daugybę skyrių, dėl kurių ląstelė negali būti vaizduojama kaip vienalytė masė, kurioje sumaišomos įvairios medžiagos. Sąlygos viename skyriuje gali labai skirtis nuo kitų; joje vykstantys procesai gali vykti tik jame, o vienas kitame vyksta skirtingi procesai.
Galiausiai, endoplazminio tinklelio membranos yra tie paviršiai, kuriais sklinda biologinės srovės, kurie yra signalai, kurie keičia selektyvų membranų pralaidumą ir tuo pačiu fermentų aktyvumą. Dėl to kai kurios cheminės reakcijos yra išjudinamos, kitos yra slopinamos - metabolizmas paklūsta reguliavimui ir vyksta koordinuotai.
Daugelis ląstelėje sintetinamų medžiagų turi būti sukoncentruotos ir iš ląstelės išsiskyrusios arba į išorinę aplinką, arba į tarpląstelinę vakuolę. Be to, ląstelė taip pat koncentruoja medžiagas, kurios patenka į ją iš kitų ląstelių, pavyzdžiui, jei jas laiko atsargose. Šį darbą atlieka diktozomos. Paprastai augalo ląstelėje yra kelios diktiozomos, o visas jų rinkinys vadinamas „Golge“ aparatu (arba kompleksu) ir duota ląstele. Kiekviena diktiozoma yra sukrautų membranų sistema (10 lentelė - pjūvis, 32 pav. - tūrinė diagrama). Ertmės tarp membranų, kurios sudaro šį organoidą, atrodo kaip siauri plyšiai, tada plokšti maišeliai - cisternos, tada burbuliukai. Jų forma keičiasi veikiant organoidui ir, matyt, priklauso nuo tarpmembraninių erdvių užpildymo išsiskiriančiomis ir susikaupusiomis medžiagomis laipsnio. Susiformavusios ir peraugusios pūslelės atskiriamos nuo organoido. Akivaizdu, kad daugelis ląstelių vakuolių, apsuptų membranų - tonoplastu, yra Golgi aparato veiklos produktas; tai yra nuo jo atsiplėšę ir po to padidėję burbuliukai.
„Golgi“ aparatas ypač sukurtas šalinimo (sekrecijos) ląstelėse, kuriose kaupiasi įvairios medžiagos arba iš kurių išsiskiria įvairios medžiagos. Jis sintetina ir išskiria medžiagas, kurios sudaro ląstelės sienelę.
Lizosomos yra gana maži (apie 0,5 mikrono skersmens) suapvalinti kūnai - dar vienas citoplazmos organoidas. Jie yra padengti membrana - lipoproteinų membrana. Lizosomų turinys yra fermentai, virškinantys baltymus, angliavandenius, nukleorūgštis ir lipidus. Lizosomos membrana užkerta kelią fermentų išsiskyrimui iš organoido į hialoplazmą, kitaip pastarieji būtų suardomi šiais fermentais.
Galima manyti, kad lizosomos yra Golgi aparato veiklos produktai, nuo jo atsiskyrę burbuliukai, kuriuose šis organoidas yra sukaupęs virškinimo fermentus.
Tas ląstelės dalis, kurios miršta jos vystymosi metu, sunaikina lizosomų fermentai. Mirusioje ląstelėje lizosomos sunaikinamos, fermentai randami citoplazmoje, o visa ląstelė, išskyrus membraną, virškinama.
Ribosomos yra labai maži organeliai, apie 250A skersmens. Jie yra beveik sferinės formos. Kai kurie iš jų yra pritvirtinti prie išorinių (hialoplazminių) membranų paviršių, kurie sudaro granuliuoto endoplazminio tinklo kanalus; kai kurie yra laisvos būklės hialoplazmoje. Ląstelėje gali būti iki 5 milijonų ribosomų. Jie yra "baltymų sintezės aparatai. Todėl jų yra ypač daug ląstelėse, kurios aktyviai formuoja baltymą - augančiose ląstelėse, ląstelėse, išskiriančiose baltymines medžiagas. Ribosomų taip pat yra mitochondrijose ir chloroplastuose, kur jie sintetina dalį baltymų, iš kurių jie yra sukurti. organelės.
Organelės, vadinamos mikrotubulėmis, yra daugelyje ląstelių. Jų vardas kalba apie jų formą - tai vamzdžiai, kurių viduje yra kanalas. Jų išorinis skersmuo yra apie 250A. Kartais tai yra dvigubi vamzdžiai - du pavieniai, gulintys vienas šalia kito ir turintys bendrą sienelę, skiriančią jų ertmes. Mikrovamzdelių sienos yra pastatytos iš baltymų molekulių. Manoma, kad mikrovamzdeliai siejami su citoplazmos ir jos darinių susitraukiamuoju (motoriniu) aktyvumu. Iš jų, kaip ir iš pastato dalių, matyt, yra pastatytos vėliavos, organoido, susitraukiančios struktūros, kurių pagalba juda kai kurie vienaląsčiai ir kolonijiniai dumbliai, taip pat ląstelės, kurios naudojamos daugelio žemesnių augalų dauginimui. Ląstelių dalijimosi metu iš mikrovamzdelių susidaro verpstės siūlai, kurie bus aptariami toliau. Dalijimosi laikotarpiu mikrovamzdeliai susirenka į grupes ir suformuoja šias gijas. Pasidalijimo pabaigoje gijos vėl suyra į atskirus mikrovamzdelius. Ląstelėse ar jų dalyse, kuriose trūksta tankios membranos, mikrovamzdeliai gali atlikti palaikomąją funkciją, sudarydami vidinį ląstelės skeletą.
Plastidai yra organeliai, būdingi tik augalų ląstelėms. Paprastai tai yra dideli kūnai, aiškiai matomi šviesos mikroskopu.
Yra 3 rūšių plastidai: bespalviai - leukoplastai, žali - chloroplastai, spalvoti kitomis spalvomis - chromoplastai. Kiekviena plastidų rūšis turi savo struktūrą ir atlieka savo prigimtines funkcijas. Tačiau galimi plastidžių perėjimai iš vieno tipo į kitą. Taigi bulvių gumbų žalumą lemia jų leukoplastų persitvarkymas į chloroplastus. Morkų šaknų pasėlyje leukoplastai pereina į chromoplastus. Visi trys plastidų tipai yra suformuoti iš proplastidų.
Proplastidai yra bespalviai kūnai, panašūs į mitochondrijas, tačiau šiek tiek didesni už juos. Jų daug randama meristeminėse ląstelėse. Leukoplastų yra nedažytų augalų dalių (vaisių, sėklų, šaknų, lapų epidermio) ląstelėse. Jų forma neapibrėžta. Dažniausiai yra leukoplastų, kuriuose nusėda krakmolas (jis susidaro iš cukrų). Yra leukoplastų, kurie kaupia baltymus. Rečiausiai yra riebalais užpildyti leukoplastai; jie susidaro senstant chloroplastams. Reikšmingų skirtumų tarp leukoplastų ir proplastidų nėra.
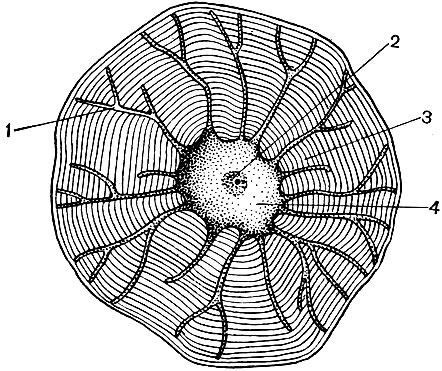
Chloroplastai yra aukštesnių augalų plastidės, kuriose vyksta fotosintezės procesas, tai yra, naudojant šviesos spindulių energiją organinėms medžiagoms susidaryti iš neorganinių medžiagų (anglies dioksido ir vandens), tuo pačiu metu deguonį išskiriant į atmosferą. Chloroplastai yra abipusiai išgaubto lęšio formos, jų dydis yra apie 4 - 6 mk. Jie randami lapų narenchiminėse ląstelėse ir kitose žaliose aukštesnių augalų dalyse. Jų skaičius ląstelėje svyruoja nuo 25 iki 30.
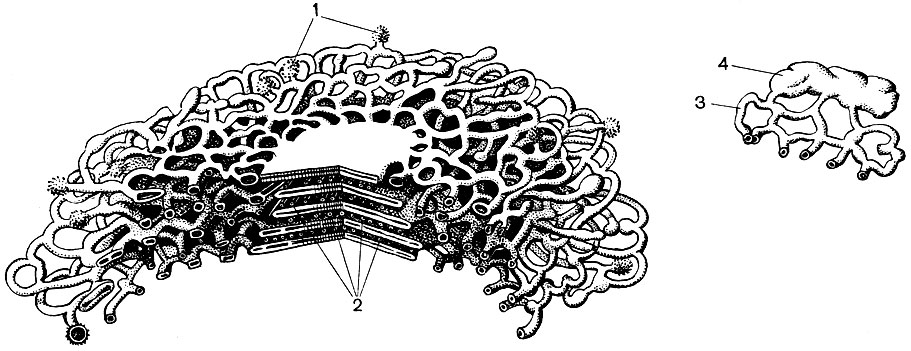
Chloroplasto struktūra pateikta 11 ir 12 lentelėse (pjūvio vaizdas) ir schema, pavaizduota 33 paveiksle. Lauke chloroplastas yra padengtas membrana, susidedančia iš dviejų lipoproteinų membranų. Pagal jį pagrindinėje medžiagoje (stromos) yra užsakyta daugybė formacijų - lamelių. Jie suformuoja plokščius maišelius, kurie įprastuose stenduose guli vienas ant kito. Šie stoikai, kurie primena monetas, sukrautas į koloną, vadinami grūdais. Per jas praeina ilgesnės lamelės, todėl visi chloroplastų grūdeliai sujungiami į vieną sistemą. Granules sudarančių membranų sudėtyje yra žalias pigmentas - chlorofilas. Čia vyksta fotosintezės šviesos reakcijos - šviesos spindulių absorbcija chlorofilu ir šviesos energijos pavertimas sužadintų elektronų energija. Elektronai, kuriuos sužadina šviesa, t. Y. Turintys energijos perteklių, atsisako energijos, kad suskaidytų vandenį ir sintezuotų ATP. Kai vanduo suyra, susidaro deguonis ir vandenilis. Deguonis išsiskiria į atmosferą, o vandenilį suriša baltymas ferredoksinas. Tada ferredoksinas vėl oksiduojamas, išleidžiant šį vandenilį į reduktorių, sutrumpintai kaip NADP. NADP pereina į redukuotą formą - NADP-H 2. Taigi, fotosintezės šviesos reakcijų rezultatas yra ATP, NADP-H 2 ir deguonies susidarymas, sunaudojama vandens ir šviesos energija.
ATP sukaupia daug energijos - tada ji naudojama sintezei, taip pat kitiems ląstelės poreikiams tenkinti. NADP-H 2 yra vandenilio akumuliatorius, kurį jis lengvai atiduoda. Todėl NADP-H 2 yra cheminis reduktorius. Daugelis biosintezių yra susijusios su redukcija, o NADPH 2 šiose reakcijose veikia kaip vandenilio tiekėjas.
Be to, naudojant chloroplastinius stromos fermentus, tai yra, už granulių ribų, vyksta tamsios reakcijos: vandenilis ir ATP esanti energija naudojami atmosferos anglies dvideginiui (CO2) sumažinti ir įtraukti jį į organinių medžiagų sudėtį. Pirmoji organinė medžiaga, susidariusi fotosintezės metu, patiria daugybę pertvarkymų ir sukelia visą augale sintetinamų ir jo kūną sudarančių organinių medžiagų įvairovę. Nemažai šių virsmų vyksta čia pat, chloroplastų stromoje, kur yra fermentų cukrams, riebalams susidaryti, taip pat viskas, kas reikalinga baltymų sintezei. Tada cukrūs gali arba pereiti iš chloroplasto į kitas ląstelių struktūras, o iš ten - į kitas augalų ląsteles, arba sudaryti krakmolą, kurio grūdelius dažnai galima pamatyti chloroplastuose. Riebalai taip pat nusėda chloroplastuose, arba lašų pavidalu, arba paprastesnių medžiagų pavidalu, riebalų pirmtakai, palieka chloroplastą.
Medžiagų komplikacija siejama su naujų cheminių ryšių sukūrimu ir paprastai reikalauja energijos. Jo šaltinis yra ta pati fotosintezė. Faktas yra tas, kad didelė fotosintezės metu susidariusių medžiagų dalis vėl suyra hialoplazmoje ir mitochondrijose (visiško degimo atveju - medžiagomis, kurios yra fotosintezės pradinė medžiaga - C0 2 ir H 2 0). Dėl šio proceso, kuris iš esmės yra atvirkštinis fotosintezės procesui, energija, susikaupusi anksčiau suskaidytų medžiagų cheminiuose ryšiuose, išsiskiria ir - vėl naudojant ATP pagalbą - išleidžiama naujiems sintetinių molekulių cheminiams ryšiams susidaryti. Taigi reikšminga fotosintezės gamybos dalis reikalinga tik tam, kad šviesos energija būtų surišta ir, ją pavertus chemine, panaudotų visiškai skirtingų medžiagų sintezei.
Šių sintezių statybinė medžiaga naudojama tik dalis organinių medžiagų, susidariusių fotosintezės metu.
Fotosintezės (biomasės) gamyba yra milžiniška. Per metus žemės rutulyje jų yra apie 1010 tonų. Augalų sukurtos organinės medžiagos yra vienintelis gyvybės šaltinis ne tik augalams, bet ir gyvūnams, nes pastarieji apdoroja paruoštas organines medžiagas, tiesiogiai maitindami augalus ar kitus gyvūnus, kurie savo ruožtu jie minta augalais. Taigi fotosintezė yra viso šiuolaikinio gyvenimo Žemėje šerdis. Visi augalų ir gyvūnų medžiagų ir energijos virsmai yra pirminių fotosintezės produktų medžiagų ir energijos pertvarkymai, rekombinacijos ir perdavimas. Fotosintezė yra svarbi visoms gyvoms būtybėms ir tam, kad vienas iš jos produktų yra laisvasis deguonis, kilęs iš vandens molekulės ir patekęs į atmosferą. Manoma, kad visas deguonis atmosferoje gaunamas fotosintezės būdu. Tai būtina kvėpuojant augalams ir gyvūnams.
Chloroplastai sugeba judėti aplink ląstelę. Esant silpnam apšvietimui, jie yra po langelio siena, nukreipta į šviesą. Tai darydami, jie didesniu paviršiumi tampa šviesa. Jei šviesa per intensyvi, jie atsisuka į ją ir išsirikiuoja palei atspalvius, lygiagrečius šviesos spinduliams. Esant vidutiniam apšvietimui, chloroplastai užima padėtį, esančią tarp dviejų kraštutinių. Bet kokiu atveju pasiekiamas vienas rezultatas: chloroplastai yra palankiausiomis fotosintezės apšvietimo sąlygomis. Toks chloroplastų judėjimas (fototaksis) yra vienos iš dirglumo rūšių augalų apraiška.
Chloroplastai turi tam tikrą autonomiją ląstelių sistemoje. Jie turi savo ribosomas ir medžiagų rinkinį, kurie lemia daugelio pačių chloroplastų baltymų sintezę. Taip pat yra fermentų, kurių darbas lemia lipidų, iš kurių susidaro lamelės, ir chlorofilo susidarymą. Kaip matėme, chloroplastas taip pat turi autonominę energijos gamybos sistemą. Dėl viso to chloroplastai gali savarankiškai kurti savo struktūras. Yra net nuomonė, kad chloroplastai (kaip ir mitochondrijos) atsirado iš kai kurių žemesnių organizmų, kurie apsigyveno augalo ląstelėje ir pirmiausia su ja pateko į simbiozę, o vėliau tapo neatsiejama jos dalimi - organoidu.
Žemesniuose augaluose fotosintezę taip pat atlieka specializuotos, nors ir ne taip gerai išsivysčiusios, kaip chloroplastų, membranų struktūros. Fotosintetinėse bakterijose membranos, kuriose yra chlorofilo, sudaro tinklą, kuris persmelkia bakterijos kūną. Mėlynuosiuose dumbliuose fotosintetinės membranos susilieja į plokščius burbulus. Žaliuosiuose ir kituose dumbliuose šių membranų sistema nuo likusios ląstelės yra atskirta dengiančiąja membrana ir suformuoja specialų organoidą - chromatoforą. Chromatoforų skaičius ląstelėje yra nedidelis; dažnai ląstelėje yra tik vienas chromatoforas. Jų forma labai skiriasi skirtingų rūšių dumbliuose.
Spirogyroje chromatoforas yra juostos formos, spiralės formos, išilgai ląstelių sienelių; Clostridium tai yra briaunoti cilindrai; zignema turi žvaigždinius kūnus.
Chromoplastai atsiranda arba iš proplastidų, arba iš chloroplastų, arba iš leukoplastų. Jų vidinė membranos struktūra yra daug paprastesnė nei chloroplastų. Gran nėra, stromoje yra daug geltonos arba oranžinės spalvos pigmento. Chromoplastų yra žiedlapių, vaisių ir šakniavaisių ląstelėse.
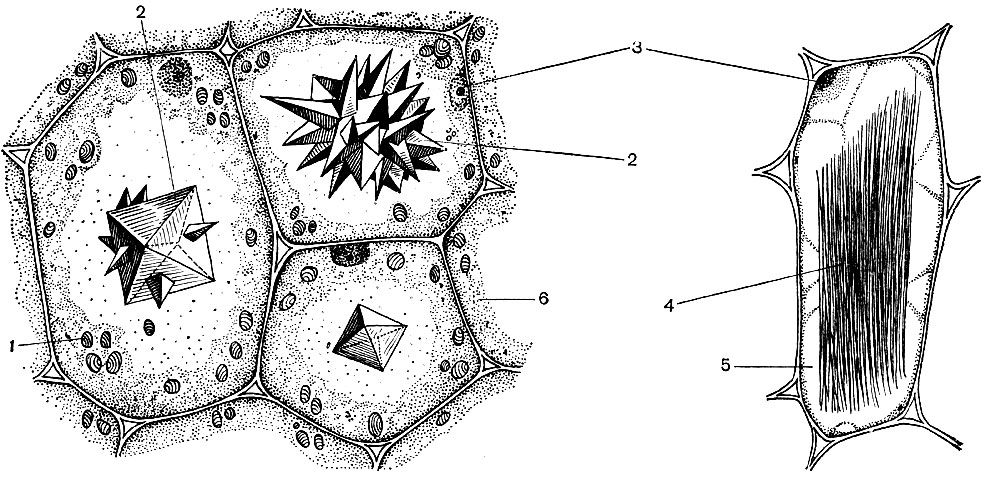
Tipiškoje augalų ląstelėje yra didelė vakuolė, užpildyta skysčio turiniu. Dažnai vakuolė užima beveik visą ląstelės tūrį, todėl citoplazma yra tik plonas sluoksnis, esantis šalia ląstelės membranos. Jaunos ląstelės turi keletą mažų vakuolių, kurios vystosi ląstelei ir susilieja į vieną. Vakuolės - ląstelių sulčių - kiekis yra daugelio medžiagų vandeninis tirpalas: cukrus, amino rūgštys, kitos organinės rūgštys, pigmentai (dažikliai), vitaminai, taninai, alkaloidai, glikozidai, neorganinės druskos (nitratai, fosfatai, chloridai), o kartais ir baltymai.
Visos šios medžiagos yra ląstelės atliekos. Kai kurie jų laikomi vakuolinėse (ląstelių) sultyse kaip atsarginės medžiagos ir laikui bėgant vėl patenka į citoplazmą naudoti. Kiti yra medžiagų apykaitos produktai, pašalinti iš citoplazmos. Taigi, oksalo rūgštis pašalinama į vakuolę; vakuoliniame sape kalcio oksalato kristalai dažnai būna nusėdę - kartais pavienių kristalų pavidalu, kitais atvejais šios druskos kristalų - daugialypių (druzų) arba panašių į adatų (rafų) - konglomerato pavidalu, parodytų 34 paveiksle.
Didžioji dauguma augalų (išskyrus prokariotinius organizmus) kiekviena gyva ląstelė turi branduolį arba kelis branduolius. Ląstelė be branduolio gali gyventi tik trumpą laiką. Sietelių mėgintuvėlių anucleated ląstelės yra gyvos ląstelės. Bet jie negyvena ilgai. Visais kitais atvejais nebranduolinės ląstelės yra negyvos.

Branduolys visada glūdi citoplazmoje. Branduolio forma gali būti skirtinga - apvali, ovali, stipriai pailga, netaisyklingai daugialypė. Kai kuriose ląstelėse jo veikimo metu pasikeičia branduolio kontūrai, jo paviršiuje susidaro įvairaus dydžio skiltelės. Branduolių dydžiai nėra vienodi skirtingų augalų ląstelėse ir skirtingose \u200b\u200bto paties augalo ląstelėse. Jaunose, meristematinėse ląstelėse yra gana dideli branduoliai, kuriuose jie gali užimti iki 3/4 visos ląstelės tūrio. Santykinis ir kartais absoliutus branduolių dydis išsivysčiusiose ląstelėse yra daug mažesnis nei jaunų.
Lauke šerdis yra padengta apvalkalu, susidedančiu iš dviejų membranų, tarp kurių yra tarpas - perinuklearinė erdvė. Korpusą pertraukia poros. Iš išorės iš dviejų apvalkalo membranų išauga ataugos, tiesiogiai pereinančios į citoplazmos endoplazminio tinklo sieneles. Tiek poros, tiek tiesioginis endoplazminio tinklo sujungimas su perinuklearine erdve suteikia glaudų kontaktą tarp branduolio ir citoplazmos.
Branduolio turinys yra granuliuota pagrindinė medžiaga (branduolio sultys, arba nukleoplazma), kurioje yra tankesnės struktūros - chromosomos ir branduolys. Branduolys yra aparatas, skirtas iš šios medžiagos sintezuoti ribosomų medžiagą ir jų surinkimo vietą.
Chromosomos yra sudarytos iš daugybės deoksiribonukleino rūgšties (DNR) molekulių, sujungtų su histono baltymo molekulėmis. DNR molekulės yra ilgos, kompleksiškai supakuotos dvigubos grandinės. Kiekviena molekulė susideda iš dviejų sruogų, spirališkai susisukusių aplink vieną kitą. Savo ruožtu gija yra labai daug vadinamųjų nukleotidų grandinė. Nukleotidas yra azoto bazės, angliavandenių (dezoksiribozės) ir fosforo rūgšties junginys.
Kiekviename iš nukleotidų yra viena iš šių keturių azoto bazių: adeninas, guaninas, citozinas arba timinas. Atitinkamai DNR išskiriami 4 skirtingi nukleotidai: adeninas (A), guaninas (G), citozinas (C) ir timinas (T). Nukleotidai sujungiami per jų fosfatų grupes, taip sudarant ilgą grandinę. Dvi grandinės, susisukusios aplink viena kitą ir sudarančios vieną DNR molekulę, sujungiamos jų nukleotidų azoto bazių chemine sąveika (vadinamosios vandenilio jungtys). Pagrindai formuoja poras - vienas pagrindas iš vienos grandinės, kitas iš antros. Diagrama, pavaizduota 35 paveiksle, suteikia idėją, kokia tvarka nukleotidai sujungiami į grandinę, o dvi grandinės - tarpusavyje.
Nors visos dezoksiribonukleino rūgšties molekulės yra pastatytos pagal aprašytą vieną planą, jų specifinė kokybinė sudėtis skiriasi; jie taip pat skiriasi molekulių dydžiu. DNR molekulėje yra iki 50-100 tūkstančių bazių porų, tačiau jų porų skaičius kiekvienoje molekulėje skiriasi nuo nukleotidų porų skaičiaus bet kurioje kitoje molekulėje. Be to, yra 4 skirtingi nukleotidų tipai, ir kiekviena DNR molekulė turi savo būdingą kiekvienos rūšies nukleotidų procentą. Kitaip tariant, kiekvienos DNR molekulės kiekybinis santykis A: G: C: T yra skirtingas. Ir galiausiai kiekvienoje DNR molekulėje A, G, C, T nukleotidų kaitaliojimo tvarka būdinga tik šiai molekulei. Tik dviejų 50 tūkstančių bazių porų, net ir gretimų, prasiskverbimas dramatiškai pakeičia visos molekulės savybes; tas pats, pakeisdamas bent vieną porą, tarkim Nuo - G iki A - T arba G - Iz; tas pats - vienos poros nebuvimas iš tų pačių 50 tūkstančių. Realiai DNR molekulės gali skirtis viena nuo kitos ne viena grandimi, ne viena ar dviem nukleotidų poromis, bet vienu metu didžiuliu jų skaičiumi. DNR molekulėje galimų nukleotidų porų permutacijų skaičius yra begalinis, atitinkamai, begalinis skirtingų molekulių skaičius, kurių kiekviena turi savo savybių. Nukleotidų seka DNR molekulėje yra užšifruotas konkretaus baltymo, būdingo tam tikrai ląstelei, sudėties įrašas. Kiekvienoje DNR molekulėje nuosekliai sutampa kelių baltymų sudėties įrašai, o visose branduolio chromosomose visose DNR molekulėse - įrašai apie visų ląstelės baltymų sudėtį, kurie gali būti sintetinami joje per visą jos gyvenimą. Šių užkoduotų įrašų esmė yra tokia.
Kiekvieno baltymo molekulės yra aminorūgščių grandinės, sujungtos nuosekliai. Yra apie 20 skirtingų amino rūgščių, ir baltymo pobūdį lemia tai, iš kurių amino rūgščių susideda jo molekulė, koks yra jų bendras kiekis molekulėje ir kokia seka jie yra susiję vienas su kitu. Pavyzdžiui, DNR dalis, atsakinga už tam tikro baltymo sudėtį (kiekvienas toks skyrius vadinamas genu), yra įrašas apie tam tikrą aminorūgščių seką, kuri sudaro tam tikro baltymo molekulę, bendrą jų skaičių joje. Kas 3 vienas po kito einantys DNR grandinės nukleotidai žymi (koduoja) vieną atitinkamos baltymo molekulės aminorūgštį. Kiti 3 nukleotidai koduoja kitą aminorūgštį ir taip toliau. Galimų skirtingų 3 nukleotidų derinių iš keturių skirtingų tipų skaičius yra daugiau nei pakankamas 20 aminorūgščių kodavimui. Įdomu tai, kad tas pačias amino rūgštis visame gyvajame pasaulyje koduoja tie patys nukleotidų deriniai. Todėl viename gene jo nukleotidų kompozicija koduoja visų baltymų, kuriuos galima sintetinti tam tikroje ląstelėje, aminorūgščių sudėtį.
Ląstelių baltymai yra fermentai. Fermentai lemia visų reakcijų, sudarančių ląstelių gyvybinės veiklos esmę, eigą. Visų kitų ląstelės medžiagų susidarymas ir transformacija priklauso nuo tam tikrų fermentų buvimo - riebalų, angliavandenių, alkaloidų, dervų ir kt. kuriame užkoduota kito fermentų rinkinio sudėtis. Štai kodėl informacija apie visų baltymų, kurie gali susidaryti ląstelėje, sudėtį yra informacija apie visas ląstelės ir organizmo savybes. Toliau pamatysime, kad ši informacija yra paveldima, tai yra, kad ji visiškai perduodama iš ląstelės į ląstelę jų dauginimosi metu ir iš motininio augalo į dukterinius augalus.
Ši informacija yra saugoma chromosomose. Tačiau šios informacijos realizavimas - baltymų sintezė - juose nevyksta. Chromosomų genai veikia tik kaip šios sintezės iniciatoriai. Kai ląstelei reikia suformuoti tam tikrą baltymą, suaktyvinamas genas, kuriame užšifruota šio baltymo sudėtis. Tai reiškia, kad DNR grandinės dalyje, kuri sudaro šį geną, susidaro vadinamosios informacinės ribonukleino rūgšties (i-RNR) molekulės.
Šių molekulių, kurios yra vienos nukleotidų grandinės, struktūra ir sudėtis atspindi geno, ant kurio jos susidarė, nukleotidų struktūrą. Taigi informacija apie būsimo baltymo sudėtį yra nukopijuota.
M-RNR molekulių susidarymas (daug jų susidaro ant kiekvieno geno) taip pat reiškia informacijos dauginimąsi, tarytum, daugybės identiškų matricų, atspindinčių to paties geno struktūrą, atspaudą ir tokiu būdu perduodant informaciją apie tam tikro baltymo aminorūgščių sudėtį jų molekulių nukleotidų sekoje. Šie genų atspaudai pereina iš branduolio į citoplazmą. Čia iššifruojama informacija, esanti matricose - i-RNR molekulėse, informacija realizuojama, ji iš nukleotidų sekos kalbos išverčiama į aminorūgščių sekos kalbą, t. Y. Į tam tikros kompozicijos baltymų molekulių sintezę.
Dekoduojant ir sintezuojant, be i-RNR molekulės, dalyvauja daugybė įvairių transportinių ribonukleino rūgščių (t-RNR) molekulių, ribosomos ir daugybė fermentų. Aminorūgštys jungiasi prie t-RNR - molekulės molekulės. Kiekviena iš 20 aminorūgščių rūšių turi savo t-RNR. Taigi, t-RNR molekulė turi cheminių grupių, kurios gali atpažinti jos aminorūgštį, pasirinkdamos ją iš visų galimų aminorūgščių. Tai atsitinka naudojant specialius fermentus. Atpažinusi aminorūgštį, t-RNR užmezga ryšį su ja. Ribosoma yra prijungta prie i-RNR molekulės pradžios, kuri, judėdama palei i-RNR, polipeptidinėje grandinėje jungiasi vienas su kitu būtent su tomis aminorūgštimis, kurių eilę užšifruoja duotos i-RNR nukleotidų seka. Šis informacijos skaitymas, jo dekodavimas įvyksta dėl specialaus transporto RNR esančio mechanizmo, kurio molekulės sustato būtent tas aminorūgštis, kurios yra „pažymėtos“, „užrašytos“ i-RNR molekulės nukleotidų seka. Ribosoma yra aparatas, skirtas chemiškai surišti aminorūgštis į baltymo molekulę.
Taigi, bendra schema yra tokia. Ant aktyvuoto geno - vienos iš DNR molekulės grandinių atkarpos - sintetinamos nukleotidų grandinės, i-RNR molekulės, kurių sudėtis tiksliai atspindi geno sudėtį, todėl turi šio geno koduojamo baltymo sudėties įrašą. Ribosomos citoplazmoje, remiantis informacija, kurią iš geno pernešė m-RNR molekulės, naudojant transportines RNR, tam tikroje sekoje sujungiamos skirtingos aminorūgštys, todėl susidaro baltymo molekulė, kurios sudėtis užkoduota šiame gene.
I-RNR molekulių susidarymas ant aktyvinto geno yra komanda ir speciali programa griežtai apibrėžto baltymo sintezei. Ta pati mRNR molekulė naudojama kaip planas kuriant daugybę tų pačių baltymų molekulių. Tačiau tai yra gana trumpalaikė, todėl norint ilgalaikę naujų to paties baltymo molekulių sintezę, ant to paties geno reikia suformuoti identiškas, tačiau naujas m-RNR molekulių kopijas. Perėjus genui į neaktyvią būseną - ir tai atsitinka, kai ląstelėje išnyksta tam tikro baltymo poreikis - jis yra užblokuotas, nustoja formuotis i-RNR ir netrukus šio baltymo sintezė sustoja. Ląstelės gyvenimo metu jai reikia įvairių baltymų. Kiekvieną kartą suveikia juos apibrėžiantys genai ir susidaro atitinkama i-RNR molekulių sudėtis.
Kiekviena ribosoma per savo gyvenimą gamina daug skirtingų baltymų molekulių. Jis gali veikti bet kurios i-RNR pagrindu, o rezultatas, jo sukurto baltymo pobūdis priklauso tik nuo i-RNR, turinčios sąlytį, su kuria ribosoma šį kartą veikė, sudėties.
Taigi ląstelės branduolys atlieka šias tarpusavyje susijusias funkcijas. Joje kaupiama informacija apie visų baltymų, kurie gali būti sintetinami tam tikroje ląstelėje per gyvenimą, sudėtį. (Išimtis yra keletas mitochondrijų ir chloroplastų baltymų. Jų sudėtis yra užkoduota jų pačių šių organelių DNR, kur yra šios DNR. Čia vietoje vyksta visi šiose DNR esančios informacijos dekodavimo etapai, įskaitant šių baltymų sintezę naudojant savo ribosomų.)
Branduolys kaupia informaciją apie visas ląstelės ir organizmo savybes. Branduolys tinkamu laiku organizuoja kiekvieno iš šių baltymų sintezę. Ląstelių dalijimosi metu kartu su branduolio dalijimusi visa ši informacija visiškai perduodama kiekvienam naujai susiformavusiam branduoliui, kiekvienai naujai ląstelei. Tai įmanoma dėl to, kad prieš dalijantis visa chromosomų genų medžiaga savaime padvigubėja, susidaro dvi vienodos jos kopijos ir viena iš jų patenka į kiekvieną naują branduolį. Kiekvienos ląstelės branduolyje yra visas genas, būdingas tam tikram organizmui. Tačiau ne visi genai veikia įvairių specializuotų ląstelių gyvenimo metu. Vienose ląstelėse veikia viena genų dalis, kitose - kita, trečiose - trečia. Štai kodėl vieno organizmo skirtingų audinių ląstelės skiriasi viena nuo kitos. Nemaža dalis genų išlieka pasyvi, tik „kaupia informaciją“ nuo gimimo iki ląstelės ar organizmo mirties. Be to, tam tikroje ląstelėje esantys genai neveikia vienu metu: kai kurie yra aktyvūs vienu ląstelės gyvenimo periodu, tam tikru jos vystymosi etapu; kiti - iš kitos ir t.
Kodėl vieni genai lieka neaktyvūs, kaip kiti įsijungia ir išsijungia, yra labai sudėtinga ir svarbi problema, kuri dabar intensyviai tiriama. Atrodo, kad baltymai-histonai vaidina svarbų vaidmenį blokuojant ir atblokuojant genus. Jie yra chromosomų dalis, būdami kartu su DNR. Galbūt geno suaktyvinimas, „išlaisvinimas“ įvyksta, kai histono molekulė yra atjungta nuo atitinkamos DNR dalies, taip atskleidžiant jos grandines, leidžiant joms atsiskleisti ir pradėti veikti chemiškai. Histono prijungimas lemia geno blokavimą. Tačiau nepakankamai aišku, kaip kontroliuojamas histono pritvirtinimas ir atsiskyrimas.
Genų medžiaga, kaip sakoma, yra chromosomose. Todėl branduolio darbas paveldimai informacijai saugoti, dauginti ir perkelti iš ląstelės į ląstelę, organizuoti įvairių baltymų sintezę per ląstelės gyvenimą pirmiausia yra chromosomų darbas. Nors jie egzistuoja visą ląstelės gyvenimą, branduolyje jie aiškių struktūrų pavidalu išskiriami tik dalijant ląsteles. Šiuo metu mikroskopijos metodais galima nustatyti chromosomų ir chromosomų medžiagos kondensaciją kaip atskirus, aiškiai apibrėžtus darinius.
Chromosomos yra skirtingos formos. Tai arba tiesios, arba išlenktos lazdos, arba įvairūs kabliukai, ovalūs kūnai, rutuliai (36 pav.). Jų dydis labai skiriasi. Kiekvienos tam tikros rūšies augalo ląstelės branduolyje yra tas pats chromosomų rinkinys (ar rinkiniai) iš griežtai apibrėžto skaičiaus skirtingų, tačiau griežtai apibrėžtų chromosomų. Visuose aukštesniuose ir kai kuriuose žemesniuose augaluose per savo gyvenimo ciklą pakaitomis keičiasi dvi kartos: su ląstelėmis, kurių branduoliuose yra vienas (haploidinis, n) chromosomų rinkinys, ir su ląstelėmis, kurių branduoliuose yra dvigubas (diploidinis, 2 n) chromosomų rinkinys. Haploidinis rinkinys susideda iš n skirtingų chromosomų, po vieną iš kiekvieno tipo. Skaitinė vertė yra griežtai pastovi visų tam tikros rūšies visų augalų haploidinėms ląstelėms. Taigi laukinėms braškėms šis skaičius yra 7, žirniams - taip pat 7, bet pupelėms - 11, obelei - 17, molinei kriaušei - 51, cukranendrėms - 60. Diploidinis rinkinys yra du haploidiniai, sulankstyti kartu. Jame yra dvi kiekvieno tipo chromosomos. Jei žirnyje haploidiniame rinkinyje yra 7 skirtingos chromosomos, tada diploidiniame rinkinyje yra 7 skirtingos chromosomų poros, o kiekvienoje poroje yra dvi tos pačios chromosomos. Nesunku apskaičiuoti, kad išvardytiems augalams braškių diploidų rinkinys (2n) yra 14, žirnių - 14, pupelių - 22, obelų - 34, molinės kriaušės - 102 ir cukranendrių - 120 chromosomų.
Samanų ir paparčių gyvenimo cikle organizmai kaitaliojasi su diploidinėmis ir haploidinėmis ląstelėmis. Paparčiuose šie organizmai gyvena nepriklausomą, izoliuotą gyvenimą. Samanose diploidinis augalas gyvena ant haploidinio augalo. Diploidiniame organizme susidaro haploidinės ląstelės - sporos. Kiekvienas iš jų dygdamas duoda haploidinį organizmą, tai yra organizmą, pastatytą iš haploidinių ląstelių. Jame susidaro haploidinės lytinės ląstelės - gametos.
Apvaisinimo metu vyriškos ir moteriškos lytinės ląstelės susijungia, susidaro viena ląstelė - zigota, turinti vieną branduolį - abiejų lytinių ląstelių branduolių susiliejimo rezultatas. Iš kiekvienos gametos zigotos branduolys gauna haploidinį chromosomų rinkinį ir dėl to turi dvigubą, diploidinį jų rinkinį. Organizmas išsivysto iš zigotos, kurios kiekvienoje ląstelėje yra diploidinis chromosomų rinkinys.

Diploidiniame rinkinyje dvi kiekvienos poros chromosomos yra identiškos formos, vidinės struktūros ir jose yra genų, kontroliuojančių homogeniškų požymių išvaizdą (36 pav.). Jie vadinami homologinėmis chromosomomis. Vienas iš jų yra iš haploidinio tėvo gametos rinkinio, kitas iš motinos. Todėl dviviečiuose organizmuose vienas jų turi genų, kurie lemia jo jurisdikcijoje esančių požymių raidą pagal tėvo tipą, antrasis - pagal motinos tipą.
Homologiškos kitos poros chromosomos vienodai lemia kitos simbolių serijos, trečiosios poros-trečiosios eilės, raidą ir tt Nors tėvo ir motinos organizmai priklauso tai pačiai rūšiai, jų paveldimos savybės nėra tapačios - jie taip pat turi individualių skirtumų. Todėl kai kurie vienos homologinės chromosomos genai nėra tapatūs atitinkamiems antrosios genams.
Haploidinis rinkinys, kuris yra diploido dalis ir kilęs iš tėvo gametos, turi savo tėvystės paveldimumą savo individualiais bruožais, o haploidinis rinkinys iš motinos gametos - motinišką. Dviejų haploidinių rinkinių, kurie kartu sudaro vieną diploidą, homogeniškų, bet ne visada identiškų genų kompleksinė sąveika lemia, kokie bruožai pasireikš diploidiniame palikuonyje, kuris iš esmės yra tėvo ir motinos hibridas.
Gymnospermuose ir angiospermuose kartos keičiasi iš principo, kaip ir samanose ir paparčiuose, tačiau jų haploidinė fazė labai sumažėja ir dažnai ją reprezentuoja tik ląstelių grupė. Ji gyvena ne savarankiškai, o haploidinio augalo kūne. Angiospermose patelė haploidinė karta yra uždaryta kiaušialąstėje esančiame embriono maišelyje, o vyriška - žiedadulkių grūduose.
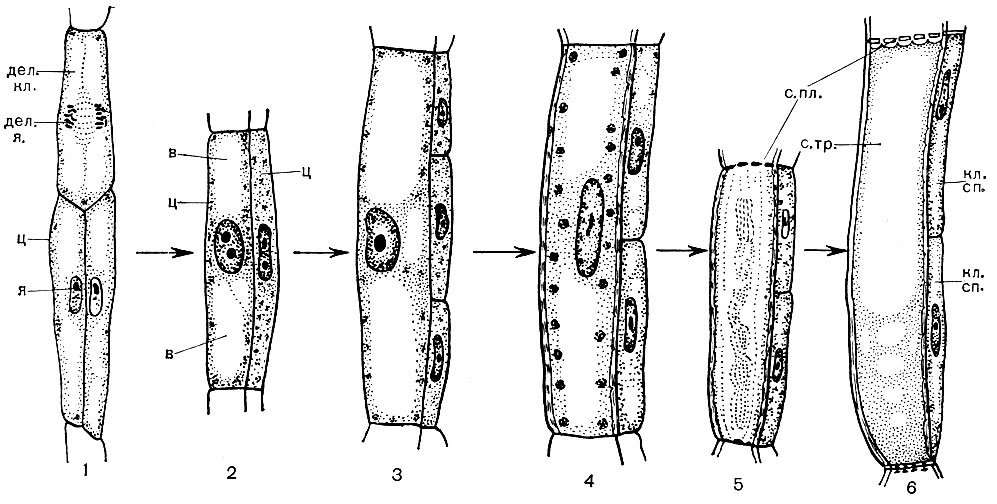
Dar prieš ląstelių dalijimąsi kiekviena DNR molekulė kiekvienoje chromosomoje pritvirtina savo kopiją - tos pačios molekulės sekundę. Todėl visa paveldima ląstelės medžiaga padvigubėja, o kiekviena chromosoma dabar susideda iš dviejų lygių dalių - chromatidų. Be to, ląstelės „užduotis“ yra padalinti kiekvieną chromosomą į chromatidus, griežtai jas vienodai paskirstyti tarp būsimų dukterinių ląstelių: kiekvienai iš jų turi būti siunčiama po vieną chromatidę iš kiekvienos chromosomos. Tai daroma taip. Prieš pat ląstelių dalijimąsi chromosomos stipriai kondensuojasi ir susitraukia. Tada jie išsidėstę vienoje plokštumoje - išilgai branduolio pusiaujo, kiekvieno jų chromatidai nukreipti į vieną ląstelės polių, kiti - į priešingą pusę. Branduolio apvalkalas išnyksta, o branduolys taip pat ištirpsta. Tarp ląstelės polių atsiranda gijos, kurios kartu sudaro verpstės formą. Gijos surenkamos iš mikrovamzdelių, jungiančių vienas kitą. Verpstę sudaro dviejų rūšių siūlai. Kai kurios yra ištisinės ląstelės, einančios iš vieno poliaus į kitą. Kiti traukia, kiekvienas iš jų sujungia stulpą su viena iš chromatidžių. Chromosomoje yra vieta - kinetochoras, prie kurio pritvirtinti traukiamieji siūlai - vienas iš vieno poliaus, kitas iš kito. Be to, chromatidės skiriasi. Chromatidai, kurie anksčiau sudarė kiekvieną chromosomą, yra atskirti vienas nuo kito ir, traukiami siūlais, išsiskiria į priešingus ląstelės polius. Todėl kiekviename poliuje surenkamas vienas pilnas chromatidų rinkinys, kuris dabar yra chromosomos. Po to suklio siūlai suyra, aplink kiekvieną chromosomų rinkinį susidaro branduolio apvalkalas, chromosomos labai išsipučia (despiralizuojasi). Kiekviename branduolyje atsiranda branduolys. Branduolys įgyja nesidalijančių ląstelių branduoliams būdingą struktūrą. Vidutinėje ląstelės plokštumoje susidaro pertvara, padalijanti ląstelę į dvi dukterines ląsteles.
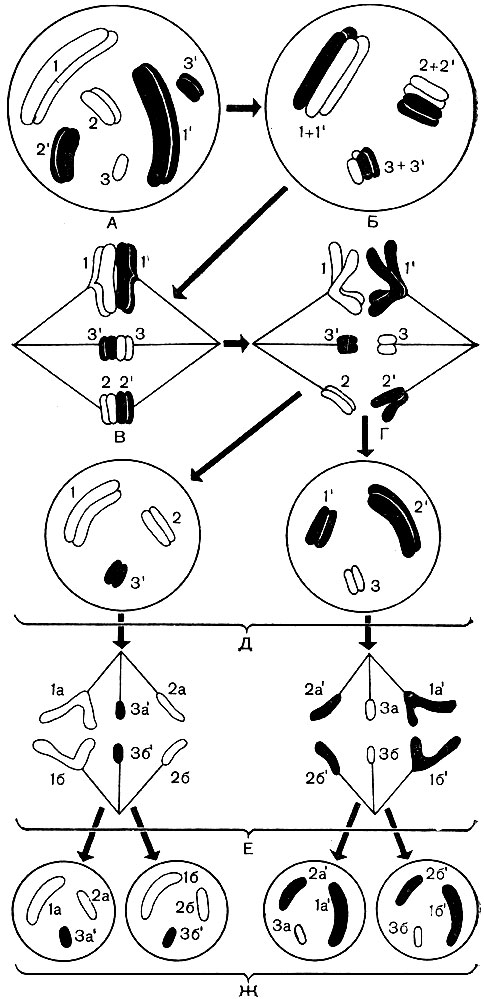
Paveikslėlis: 38. „Sąlyginės“ ląstelės chromosomų elgesio schema mejozės metu. A - vaizduojamas diploidinių ląstelių branduolys, kuriame yra 3 poros chromosomų (1 ir 1 "- 1-oji homologinių chromosomų pora, 2 ir 2" - 2-oji pora, 3 ir 3 "- 3-ioji pora). 2n \u003d 6,3 motinos organizmo chromosomos yra lengvos (skaičiai be insulto); jų porinės homologinės chromosomos, kilusios iš tėvo organizmo, yra pajuodusios (skaičiai su insultu). Homologinės chromosomos žymimos bendru skaičiumi. Kiekviena chromosoma susideda iš dviejų chromatidų. B - homologinių chromosomų susiliejimas. Šiame etape homologinės chromosomos keičiasi atskiromis sekcijomis tarpusavyje. Dėl to vyksta tam tikras motinos ir tėvo paveldimos medžiagos persiskirstymas tarp chromosomų (perėjimas per). C - susidaro verpstės siūlai, prisitvirtinę prie chromosomų, branduolio membrana išnyksta. D - homologinės chromosomos išsiskiria į priešingus polius. ląstelių, poliuose yra viena homologinė chromosoma iš kiekvienos poros, bendras chromosomų skaičius kiekviename poliuje yra perpus mažesnis nei pradiniame branduolyje A. D - vaizdas kiekviename yra du branduoliai su haploidiniu chromosomų rinkiniu; vienas iš jų gavo daugiau močiutės (šviesios, motiniškos A branduolio atžvilgiu) ir mažiau senelio (pajuodusios, tėvo atžvilgiu A branduolio atžvilgiu) chromosomų, kitos - priešingai; taigi nauji branduoliai nėra visiškai identiški savo paveldimos medžiagos sudėtyje; jų skirtumas taip pat atsiranda dėl perėjimo, vykstančio su chromosomomis B stadijoje. E - G - kiekvieno haploido branduolio D mitozinis dalijimasis: išilginis kiekvienos chromosomos skaidymasis, chromosomų divergencija į polius, dviejų haploidinių branduolių susidarymas iš kiekvieno branduolio D. Dėl to atsirado 4 ląstelės. su haploidiniais branduoliais vietoj vienos ląstelės su diploidiniu chromosomų rinkiniu
Organelės nėra paskirstytos griežtai vienodai tarp dukterinių ląstelių, tačiau tada ląstelėse sintetinamos jų sudedamosios dalys, surenkamos naujos kiekvienos rūšies organelių kopijos ir atstatomas jų skaičius kiekvienoje ląstelėje. Pačios ląstelės auga. Chromosomose paveldima medžiaga yra dubliuojama, po to chromosomos, kaip ir prieš ląstelių dalijimąsi, susideda iš dviejų chromatidžių. Ląstelė yra paruošta naujai dalybai.
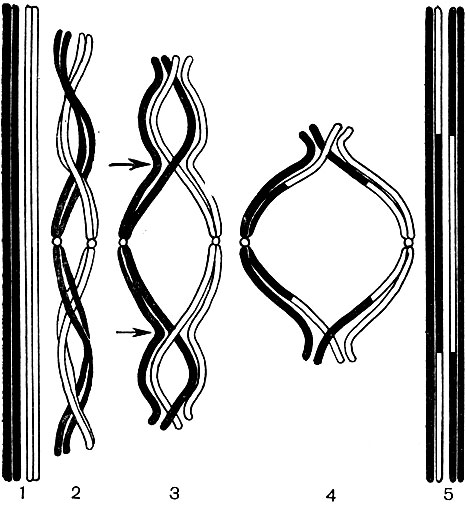
Paveikslėlis: 38A. Kryžminio schema mejozės metu B stadijoje, pav. 38. 1 - dvi homologinės chromosomos, kurių kiekviena susideda iš dviejų seserinių chromatidžių; 2 - homologinės chromosomos yra susipynusios viena su kita; 3 - kiekviena chromosoma yra padalinta į ją sudarančias chromatides; chromosomos pradeda viena kitą atstumti ir išsiskirti, tačiau kai kuriais momentais (paveiksle - keturiose) jos išlieka susijusios; seserinės chromatidės yra dvigubai padidintos centrinėje, o ne seserinės - abiejose kraštinėse kilpose; taškai, kur įmanoma kirsti, rodomi rodyklėmis; 4 - chromatidai apsikeitė vietomis, o chromosomos, patyrusios savotišką abipusę hibridizaciją, išsiskiria; 5 - dvi išsiskyrusios, bet jau „hibridinės“ (skirtingai nuo 1 pavaizduotos) chromosomos
Kai diploidinė ląstelių karta pakeičiama haploidine, įvyksta vadinamasis redukcinis branduolio dalijimasis - mejozė. Mejozės metu (38 pav.) Kiekvienos poros homologinės chromosomos artėja viena prie kitos, glaudžiai viena prie kitos susilieja išilgai ir pasisuka. Atskiros sekcijos keičiamos tarp gretimų homologinių chromosomų. Dėl to kai kurie iš tėvo chromosomų genų perkeliami į motinos chromosomų sudėtį, o atitinkami motinos chromosomų genai užima laisvas vietas tėvo chromosomose (kryžminimasis per reiškinį - 38A pav.). Išvaizda šios ir kitos chromosomos dėl to nesikeičia, tačiau skiriasi jų kokybinė sudėtis. Tėvo ir motinos paveldėjimas yra perskirstomas ir maišomas. Toliau branduolio apvalkalas ir branduolys ištirpsta, susidaro verpstės aparatas, toks pats kaip mitozės metu. Homologinės chromosomos yra atskiriamos ir verpstės siūlų pagalba išsiskiria į ląstelės polius. Viename polyje yra vienas haploidinis chromosomų rinkinys (po vieną homologinę chromosomą iš kiekvienos poros), kitas turi antrą haploidinį rinkinį.
Po mejozės susidarę haploidiniai branduoliai dalijasi iš mitozės. Šiuo atveju kiekviena haploidų rinkinio chromosoma yra padalinta į dvi chromatides, jos išsiskiria ir susidaro dukterinės haploidinės ląstelės.
Mejozė skiriasi nuo mitozės dviem pagrindiniais būdais. Pirma, mejozės metu kiekviena homologinių chromosomų pora sukimba ir tarp jų pasikeičia dalis paveldimos medžiagos, ko nėra mitozės metu. Antra, mejozės metu homologinės chromosomos išsiskiria iš ląstelės polių, po vieną iš kiekvienos poros, o mitozės metu kiekviena chromosoma padalijama į chromatidus, o chromatidės - į polius, po vieną iš kiekvienos chromosomos. Dėl mejozės haploidinės ląstelės susidaro iš diploidinės ląstelės.
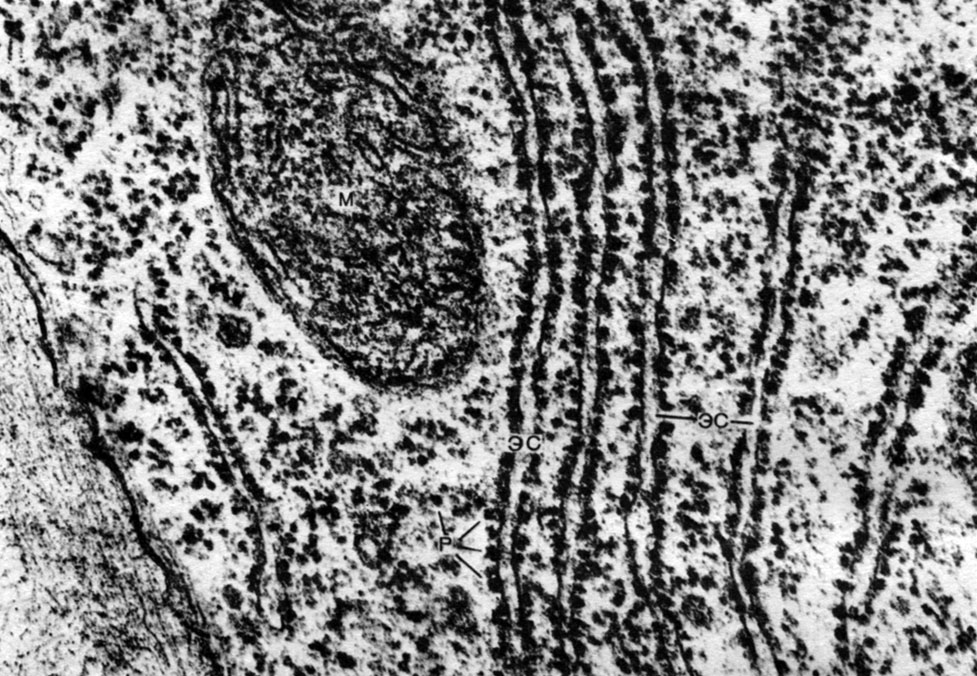
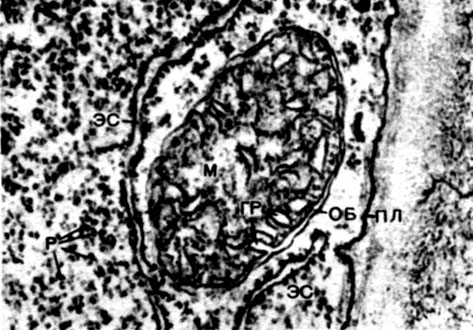
10 lentelė. Augalų ląstelių struktūra. Golgi aparatas ridiko (Raphanus sativus) besivystančių šaknų plaukų citoplazmoje. M.F. Danilovos elektronų mikrografas (padidinimas x 52000): ag - Golgi aparatas, matomi plokščių maišelių ir burbuliukų pjūviai
Kai kurių audinių ląstelėse jų vystymosi metu įvyksta neišsami mitozė: branduoliuose esanti chromosomos medžiaga padvigubėja, chromosomos dalijasi pusiau, tačiau užuot suformavusios du branduolius, jos lieka pirminiame branduolyje. Nuo to momento joje yra ne diploidinis, o tetraploidinis (keturvietis) chromosomų rinkinys. Procesas, lemiantis šį chromosomų dubliavimąsi vieno branduolio viduje, vadinamas endomitoze - vidine mitoze. Jei tai atsitinka vienam branduoliui du kartus, tada jis tampa oktoploidiniu (aštuonis kartus) rinkiniu ir kt. Ląstelės, kurių branduoliai turi daugiau nei du chromosomų rinkinius, vadinamos poliploidinėmis, tai yra multiploidinėmis. Kai kuriais atvejais ląstelių poliploidija padidina jų gyvybingumą, nes kiekvieną geną dubliuoja keli kiti to paties genai. Vienos kokybės genai veikia vieningai, o bet kurio iš jų pažeidimas nepraranda nustatyto bruožo, nes jį kompensuoja likusių homogeniškų genų darbas. Daugeliu atvejų poliploidinės ląstelės yra didesnės ir turtingesnės savo turiniu nei diploidinės. Buvo sukurtos poliploidinių kultūrinių augalų veislės, kurios padidino ekonomines savybes.
Mokslas, tiriantis ląstelę, vadinamas citologija. Ląstelė yra visų gyvų būtybių ir visų gyvenimo procesų pagrindas; todėl daugumą svarbiausių bendrųjų biologinių problemų galima išspręsti dalyvaujant citologijai ir jos tyrimų metodams.
Citologijos pagalba išsprendžiama daug praktiškai svarbių klausimų: kova su augalų ligomis (grybinėmis, bakterinėmis), naujų kultivuotų augalų veislių veisimas, hibridinių veislių sterilumo (nevaisingumo) įveikimas.
Šiuolaikinė citologija šias problemas tiria naudodama įvairius metodus - mikroskopinius, biocheminius, biofizinius, genetinius - ir glaudžiai sąveikaudama su kitais susijusiais biologijos mokslais.