VIRUSAI - mažiausi infekcinių ligų sukėlėjai. Išvertus iš lotynų virusas reiškia „nuodas, nuodingas principas“. Iki XIX amžiaus pabaigos. terminas „virusas“ medicinoje buvo vartojamas norint nurodyti bet kokį infekcinį agentą, sukeliantį ligą. Šis žodis savo šiuolaikinę reikšmę įgijo po 1892 m., Kai rusų botanikas DI Ivanovskis nustatė tabako mozaikos ligos (tabako mozaikos) sukėlėjo „filtruojamumą“. Jis parodė, kad šia liga užsikrėtusių augalų ląstelių sultys, praeinančios per specialius filtrus, sulaikančius bakterijas, išlaiko galimybę sukelti tą pačią ligą sveikiems augalams. Po penkerių metų vokiečių bakteriologas F. Löffleris atrado dar vieną filtruojamą veiksnį - galvijų snukio ir nagų ligos sukėlėją. 1898 m. Olandų botanikas M. Beijerinkas pakartojo šiuos eksperimentus išplėstine versija ir patvirtino Ivanovskio išvadas. Jis pavadino „filtruojamą nuodingą principą“, sukeliantį tabako mozaiką, „filtruojamu virusu“. Šis terminas buvo naudojamas daugelį metų ir palaipsniui buvo paverstas vienu žodžiu - „virusas“.
1901 m. Amerikos karo chirurgas W. Reedas su kolegomis nustatė, kad geltonosios karštinės sukėlėjas yra ir filtruojamas virusas. Geltonoji karštinė buvo pirmoji žmogaus liga, kuri buvo nustatyta kaip virusinė, tačiau prireikė dar 26 metų, kad jos virusinė kilmė būtų galutinai įrodyta.
Visuotinai pripažįstama, kad virusai atsirado atskirus (autonomiškus) ląstelės genetinius elementus, kurie, be to, gavo galimybę pernešti iš organizmo į organizmą. Normalioje ląstelėje kelios genetinių struktūrų rūšys juda, pavyzdžiui, matrica arba informacinė, RNR (mRNR), transpozonai, intronai ir plazmidės. Tokie mobilūs elementai galėjo būti virusų pirmtakai arba pirmtakai.
Ar virusai yra gyvi organizmai? 1935 m. Amerikiečių biochemikas W. Stanley išskyrė tabako mozaikos virusą kristalinės formos, taip įrodydamas jo molekulinę prigimtį. Gauti rezultatai sukėlė karštas diskusijas apie virusų pobūdį: ar tai gyvi organizmai, ar tik aktyvuotos molekulės? Iš tiesų, užkrėstos ląstelės viduje virusai pasireiškia kaip sudėtingesnės sudėtingos gyvosios sistemos komponentai, tačiau už ląstelės ribų jie yra metaboliškai inertiški nukleoproteinai. Virusai turi genetinės informacijos, tačiau negali jos savarankiškai įgyvendinti neturėdami savo baltymų sintezės mechanizmo. Išsiaiškinus virusų struktūros ir dauginimosi ypatybes, klausimas, ar jie gyvi, pamažu prarado savo reikšmę.
VIRUSŲ STRUKTŪRA
Išbaigta struktūra ir užkrečiama, t.y. galinti sukelti infekciją, viruso dalelė už ląstelės ribų vadinama virionu. Viriono šerdyje („šerdyje“) yra viena molekulė, o kartais dvi ar daugiau nukleorūgšties molekulių. Baltymų apvalkalas, padengiantis viriono nukleino rūgštį ir apsaugantis ją nuo žalingo aplinkos poveikio, vadinamas kapsiidu. Viriono nukleorūgštis yra genetinė viruso medžiaga (jo genomas) ir ją žymi dezoksiribonukleino rūgštis (DNR) arba ribonukleino rūgštis (RNR), tačiau niekada šie du junginiai nėra vienu metu. (Chlamidijose, riketsijose ir visuose kituose „tikrai gyvuose“ mikroorganizmuose yra ir DNR, ir RNR.) Mažiausių virusų nukleorūgštyse yra trys ar keturi genai, o didžiausiuose virusuose - iki šimto genų.
Be kapsidės, kai kurie virusai taip pat turi išorinį apvalkalą, susidedantį iš baltymų ir lipidų. Jis susidaro iš užkrėstos ląstelės membranų, kuriose yra įterptų viruso baltymų. Sąvokos „nuogi virionai“ ir „nepadengti virionai“ vartojami sinonimu. Mažiausių ir paprasčiausiai išsidėsčiusių virusų kapsulės gali būti tik vienos ar kelių rūšių baltymų molekulės. Keletas tų pačių arba skirtingų baltymų molekulių sujungiamos į subvienetus, vadinamus kapsomeromis. Savo ruožtu kapomerai formuoja taisyklingas viruso kapsidės geometrines struktūras. Skirtinguose virusuose kapsidės forma yra būdingas viriono bruožas (bruožas).
Virionai, turintys spiralės tipo simetriją, kaip ir tabako mozaikos virusas, turi pailgos cilindro formą; baltymų apvalkalo, kurį sudaro atskiri subvienetai - kapsomerai, viduje yra suvyniota nukleorūgščių (RNR) spiralė. Virionai, turintys ikosaedrinį simetrijos tipą (iš graikų kalbos eikosi - dvidešimt, hedra - paviršius), kaip ir poliovirusas, turi sferinę, tiksliau sakant, daugialypę formą; jų dangteliai yra pastatyti iš 20 taisyklingų trikampių briaunų (paviršių) ir atrodo kaip geodezinis kupolas.
Atskiriems bakteriofagams (bakterijų virusams; fagams) būdingas mišrus simetrijos tipas. Prie vadinamojo. Iš „uodeguotų“ fagų galva atrodo kaip sferinis kapidas; ilgas vamzdinis procesas - nuo jo „uodega“.
Yra dar sudėtingesnės struktūros virusų. Poksviruso virusai (raupų virusai) neturi taisyklingo, tipiško kapsidės: vamzdinės ir membraninės struktūros yra tarp šerdies ir išorinio apvalkalo.
Genetinė informacija, užkoduota viename gene, paprastai gali būti laikoma instrukcija, kaip ląstelėje gaminti specifinį baltymą. Tokią instrukciją ląstelė suvokia tik tuo atveju, jei ji siunčiama mRNR pavidalu. Todėl ląstelės, kuriose genetinę medžiagą vaizduoja DNR, turi šią informaciją „perrašyti“ (perrašyti) į papildomą mRNR viruso baltymų kopiją (taip pat žr. NUKLEINES RŪGŠTIS). DNR turintys virusai replikacijos būdu skiriasi nuo RNR turinčių virusų.
DNR paprastai egzistuoja dvigubų grandinių struktūrų pavidalu: dvi polinukleotidinės grandinės yra sujungtos su vandeniliu ir susuktos taip, kad susidarytų dviguba spiralė. RNR, priešingai, paprastai egzistuoja kaip viengrandės struktūros. Tačiau atskirų virusų genomas yra arba vienos grandinės DNR, arba dvigubos grandinės RNR. Virusinės nukleorūgšties, dvigubos arba vienkartinės, grandinės (grandinės) gali būti linijinės arba uždarytos žiede.
Pirmasis viruso replikacijos etapas yra susijęs su viruso nukleino rūgšties prasiskverbimu į ląstelę-šeimininkę. Šį procesą gali palengvinti specialūs fermentai, kurie yra viriono kapsidės arba išorinio apvalkalo dalis, o apvalkalas lieka už ląstelės ribų arba virionas praranda jį iškart, kai prasiskverbia į ląstelę. Virusas randa ląstelę, tinkamą reprodukcijai, „raktų užraktu“ kontaktuodamas atskiras savo kapidės dalis (arba išorinį apvalkalą) su ląstelės paviršiaus specifiniais receptoriais. Jei ląstelės paviršiuje nėra specifinių („atpažįstančių“) receptorių, tai ląstelė nėra jautri virusinei infekcijai: virusas į ją neprasiskverbia.
Siekiant realizuoti genetinę informaciją, į ląstelę prasiskverbusi viruso DNR specialiais fermentais perrašoma į mRNR. Gauta mRNR persikelia į ląstelių baltymų sintezės „gamyklas“ - ribosomas, kur ji pakeičia ląstelių „žinutes“ savo „instrukcijomis“ ir yra išverčiama (skaitoma), dėl ko sintetinami virusiniai baltymai. Pati virusinė DNR yra kartojama (dubliuojama) dalyvaujant kitam fermentų rinkiniui, tiek virusiniam, tiek priklausančiam ląstelei.
Susintetintas baltymas, kuris naudojamas kapsidui sukurti, ir daugybe kopijų padauginta viruso DNR sujungia ir suformuoja naujus „dukterinius“ virionus. Susiformavę virusiniai palikuonys palieka panaudotą ląstelę ir užkrėtė naujas: viruso dauginimosi ciklas kartojasi. Kai kurie virusai pumpuruodami nuo ląstelės paviršiaus užfiksuoja dalį ląstelės membranos, į kurią virusų baltymai buvo įterpti „iš anksto“, ir taip įgyja membraną. Kalbant apie ląstelę-šeimininkę, ji galiausiai pasirodo sugadinta ar net visiškai sunaikinta.
Kai kuriuose DNR virusuose pats reprodukcijos ciklas ląstelėje nėra susijęs su tiesiogine viruso DNR replikacija; vietoj to viruso DNR įterpiama (integruojama) į ląstelės-šeimininkės DNR. Šiame etape virusas, kaip vienas struktūrinis darinys, išnyksta: jo genomas tampa ląstelės genetinio aparato dalimi ir ląstelės dalijimosi metu netgi pasikartoja kaip ląstelės DNR dalis. Tačiau vėliau, kartais po daugelio metų, virusas gali vėl atsirasti - suveikia viruso baltymų sintezės mechanizmas, kuris, derinamas su viruso DNR, sudaro naujus virionus.
Kai kuriuose RNR virusuose genomas (RNR) gali tiesiogiai veikti kaip mRNR. Tačiau ši savybė būdinga tik virusams, turintiems „+“ RNR grandinę (ty, kai RNR turi teigiamą poliškumą). Virusuose, turinčiuose RNR giją „-“, pastaroji pirmiausia turi būti „perrašyta“ į „+“ giją; tik po to prasideda viruso baltymų sintezė ir virusas dauginasi.
Vadinamieji retrovirusai turi RNR kaip genomą ir turi neįprastą genetinės medžiagos perrašymo būdą: užuot perrašę DNR į RNR, kaip tai vyksta ląstelėje ir būdinga DNR turinčiams virusams, jų RNR yra perrašoma į DNR. Tada dviguba grandinė viruso DNR yra įtraukiama į ląstelės chromosomų DNR. Tokios virusinės DNR matricoje sintetinama nauja virusinė RNR, kuri, kaip ir kiti, lemia viruso baltymų sintezę. Taip pat žr. RETROVIRUSAI.
Jei virusai iš tikrųjų yra mobilūs genetiniai elementai, kurie gavo „autonomiją“ (nepriklausomybę) nuo savo šeimininkų genetinio aparato (skirtingų tipų ląstelių), tai skirtingos virusų grupės (su skirtingu genomu, struktūra ir replikacija) turėjo atsirasti nepriklausomai viena nuo kitos. Todėl neįmanoma visiems virusams sukurti vieno kilmę, susiejančio juos remiantis evoliuciniais santykiais. Gyvūnų taksonomijoje naudojami „natūralios“ klasifikacijos principai virusams netinka.
Nepaisant to, virusų klasifikavimo sistema yra būtina praktikoje, o ją sukurti bandyta ne kartą. Pasirodė, kad produktyviausias metodas pagrįstas struktūrinėmis ir funkcinėmis virusų savybėmis: norėdami atskirti skirtingas virusų grupes, jie apibūdina savo nukleorūgšties tipą (DNR ar RNR, kurių kiekviena gali būti viengrandė arba dviguba), jos dydį (nukleotidų skaičius nukleorūgščių grandinėje). rūgštys), nukleorūgščių molekulių skaičius viename virione, viriono geometrija ir kapidės bei viriono išorinio apvalkalo struktūrinės ypatybės, šeimininko tipas (augalai, bakterijos, vabzdžiai, žinduoliai ir kt.), virusų sukeltos patologijos ypatybės (ligos simptomai ir ligos pobūdis), antigeninės virusinių baltymų savybės ir organizmo imuninės sistemos reakcijos į viruso įvedimą ypatumai.
Mikroskopinių ligų sukėlėjų, vadinamų viroidais (t. Y. Į virusus panašių dalelių), grupė ne visai tinka virusų klasifikavimo sistemai. Viroidai sukelia daugybę įprastų augalų ligų. Tai yra mažiausi infekcijos sukėlėjai, neturintys net paprasčiausio baltymo apvalkalo (randami visuose virusuose); jie susideda tik iš vienos grandinės RNR, uždarytos žiede.
VIRALINĖS LIGOS
Daugelis virusų, tokių kaip tymai, herpesas ir iš dalies gripas, žmonės yra pagrindinis natūralus rezervuaras. Šie virusai perduodami ore esančiais lašeliais arba kontaktuojant.
Kai kurių paskirstymas virusinės ligos, kaip ir kitos infekcijos, kupina staigmenų. Pavyzdžiui, antisanitarinėmis sąlygomis gyvenančių žmonių grupėse beveik visi vaikai ankstyvas amžius turi poliomielitą, paprastai lengvą, ir įgyja imunitetą. Jei gyvenimo sąlygos šiose grupėse pagerės, vaikai jaunesnio amžiaus poliomielitas paprastai neserga, tačiau liga gali pasireikšti vyresniame amžiuje, o tada ji dažnai būna sunki.
Daugelis virusų negali ilgai išsilaikyti gamtoje esant nedideliam šeimininkų rūšių plitimo tankiui. Dėl primityvių medžiotojų ir augalų rinkėjų populiacijos trūkumo susidarė nepalankios sąlygos kai kuriems virusams egzistuoti; todėl labai tikėtina, kad kai kurie žmogaus virusai atsirado vėliau, atsiradus miesto ir kaimo gyvenvietėms. Manoma, kad tymų virusas iš pradžių egzistavo tarp šunų (kaip karščiavimo sukėlėjas), o žmonių raupai galėjo atsirasti dėl karvių ar pelių raupų evoliucijos. Naujausi viruso evoliucijos pavyzdžiai yra įgytas imunodeficito sindromas (AIDS). Yra įrodymų apie genetinius žmogaus imunodeficito virusų ir afrikinių žaliųjų beždžionių panašumus.
„Naujos“ infekcijos paprastai būna sunkios, dažnai mirtinos, tačiau ligos sukėlėjo evoliucijos metu jos gali tapti lengvesnės. Geras pavyzdys yra miksomatozės viruso istorija. 1950 m. Šis virusas, endeminis Pietų Amerikai ir gana nekenksmingas vietiniams triušiams, buvo įvežtas į Australiją kartu su europinėmis šių gyvūnų veislėmis. Australijos triušių, kurie anksčiau nebuvo susidūrę su šiuo virusu, liga buvo mirtina 99,5% atvejų. Po kelerių metų mirtingumas nuo šios ligos labai sumažėjo, kai kuriose vietovėse iki 50%, o tai paaiškinama ne tik „silpninančiomis“ (silpninančiomis) viruso genomo mutacijomis, bet ir padidėjusiu triušių genetiniu atsparumu ligai, ir abiem atvejais veiksminga natūrali atranka galingas natūralios atrankos slėgis.
Virusų dauginimąsi gamtoje palaiko įvairių rūšių organizmai: bakterijos, grybai, pirmuonys, augalai, gyvūnai. Pavyzdžiui, vabzdžiai dažnai kenčia nuo virusų, kurie kaupiasi jų ląstelėse didelių kristalų pavidalu. Augalus dažnai veikia maži ir paprasčiausiai sutvarkyti RNR virusai. Šie virusai net neturi specialių mechanizmų patekti į ląstelę. Juos neša vabzdžiai (kurie minta ląstelių sultimis), apvaliosios kirmėlės ir kontaktiniai, užkrėsdami augalą, jei jis yra mechaniškai pažeistas. Bakteriniai virusai (bakteriofagai) turi sudėtingiausią būdą, kaip pristatyti savo genetinę medžiagą į jautrią bakterijų ląstelę. Pirma, fago „uodega“, panaši į ploną vamzdelį, prisitvirtina prie bakterijos sienos. Tuomet specialūs „uodegos“ fermentai ištirpina bakterijos sienelės dalį ir fago genetinė medžiaga (dažniausiai DNR) įšvirkščiama į susidariusią skylę per „uodegą“, tarsi per švirkšto adatą.
Daugiau nei dešimt pagrindinių virusų grupių yra patogeniškos žmonėms. Tarp DNR turinčių virusų tai yra raupų virusų (sukeliančių raupus, vakcinijas ir kitas raupų infekcijas), herpeso grupės virusų (herpeso opos ant lūpų, vėjaraupiai), adenovirusų (ligų) šeima. kvėpavimo takai ir akys), papovavirusų šeima (karpos ir kitos odos ataugos), hepadnavirusai (hepatito B virusas). Yra daug daugiau RNR turinčių virusų, kurie yra patogeniški žmonėms. Pikornavirusai (iš lot. Pico - labai maži, angliška RNR - RNR) yra mažiausi žinduolių virusai, panašūs į kai kuriuos augalų virusus; jie sukelia ūminį poliomielitą, hepatitą A peršalimas... Mixovirusai ir paramiksovirusai yra įvairių gripo formų, tymų ir kiaulytės (kiaulytės) priežastis. Arbovirusus (iš angliškų nariuotakojų nešiojamų - „nešioja nariuotakojus“) - didžiausią virusų grupę (daugiau nei 300) - nešioja vabzdžiai, jie yra erkių ir japonų encefalito, geltonosios karštinės, arklių meningoencefalito, kolorado erkių platinamos karštinės, Škotijos encefalito ir kitų ligų sukėlėjai. ... Reovirusai, kurie yra gana reti žmonių kvėpavimo takų ir žarnyno ligų sukėlėjai, tapo ypatingo mokslinio susidomėjimo objektu dėl to, kad jų genetinę medžiagą vaizduoja dvigubos grandinės fragmentuota RNR. Taip pat žr. VENERALINĖS LIGOS; VĖJARAUPIAI; HEPATITAS; FLU; DENGE FEVER; MONONUCLEOSIS INFEKTYVI; MASOS; KRASNUHA; MENINGITIS; GAMTINIS POISKAS; POLIO; Kvėpavimo sistemos virusinės ligos; Kiaulytė; ĮGYTOS IMUNODEFICENCIJOS SINDROMAS (AIDS); ENCEFALITAS.
Kai kurių ligų, tarp jų ir labai sunkių, sukėlėjai nepriskiriami nė vienai iš minėtų kategorijų. Į specialią lėtųjų grupę virusinės infekcijos visai neseniai buvo minėta, pavyzdžiui, Creutzfeldto-Jakobo liga ir Kuru - degeneracinės smegenų ligos, kurių inkubacinis periodas buvo labai ilgas. Tačiau paaiškėjo, kad juos sukelia ne virusai, o mažiausi baltyminio pobūdžio infekciniai veiksniai - prionai (žr. PRION).
Gydymas ir profilaktika. Virusų dauginimasis yra glaudžiai susijęs su užkrėsto organizmo ląstelių baltymų ir nukleino rūgščių sintezės mechanizmais. Todėl sukurti vaistus, kurie selektyviai slopina virusą, tačiau nekenkia organizmui, yra nepaprastai sunki užduotis. Nepaisant to, paaiškėjo, kad didžiausiuose herpeso ir raupų virusuose genominė DNR koduoja daugybę fermentų, kurie savo savybėmis skiriasi nuo panašių ląstelių fermentų, ir tai buvo pagrindas vystytis antivirusiniai vaistai... Iš tiesų buvo sukurti keli vaistai, kurių veikimo mechanizmas pagrįstas viruso DNR sintezės slopinimu. Kai kurie junginiai, kurie yra per daug toksiški bendram vartojimui (į veną arba per burną), yra tinkami vietiniam vartojimui, pavyzdžiui, akių infekcijoms herpeso virusu.
Yra žinoma, kad žmogaus organizmas gamina specialius baltymus - interferonus. Jie slopina virusinių nukleorūgščių vertimą ir taip slopina viruso dauginimąsi. Genų inžinerijos dėka bakterijų gaminami interferonai tapo prieinami ir yra išbandomi medicinos praktikoje (žr. „GENE ENGINEERING“).
Veiksmingiausi natūralios organizmo gynybos elementai yra specifiniai antikūnai (specialūs imuninės sistemos gaminami baltymai), kurie sąveikauja su atitinkamu virusu ir tokiu būdu veiksmingai užkerta kelią ligos vystymuisi; tačiau jie negali neutralizuoti viruso, kuris jau pateko į ląstelę. Pavyzdys yra herpeso infekcija: Herpes virusas išlieka nervinių mazgų (ganglijų) ląstelėse, kur antikūnai negali jo pasiekti. Retkarčiais virusas suaktyvėja ir sukelia ligos atkryčius.
Paprastai dėl infekcinio agento prasiskverbimo organizme susidaro specifiniai antikūnai. Organizmui gali padėti dirbtinai padidinti antikūnų gamybą, įskaitant imuniteto stiprinimą iš anksto, skiepijant. Tokiu būdu, vykdant masinę vakcinaciją, raupų liga praktiškai buvo pašalinta visame pasaulyje. Taip pat žiūrėkite skiepijimą ir imunizaciją.
Šiuolaikiniai vakcinacijos ir imunizacijos metodai yra suskirstyti į tris pagrindines grupes. Pirma, tai yra susilpnėjusios viruso padermės naudojimas, kuris skatina antikūnų gamybą organizme, veiksmingus prieš patogeniškesnę padermę. Antra, įvedamas užmuštas virusas (pavyzdžiui, inaktyvuotas formalinu), kuris taip pat sukelia antikūnų susidarymą. Trečias variantas yra vadinamasis. „Pasyvioji“ imunizacija, t.y. paruoštų „svetimų“ antikūnų įvedimas. Gyvūnas, pavyzdžiui, arklys, yra imunizuojamas, tada antikūnai iš jo kraujo išskiriami, išgryninami ir panaudojami pacientui, kad būtų sukurtas betarpiškas, bet trumpalaikis imunitetas. Kartais antikūnai naudojami iš sergančio asmens kraujo (pavyzdžiui, tymai, erkinis encefalitas).
Virusų kaupimasis. Norint paruošti vakcinas, būtina kaupti virusą. Šiuo tikslu dažnai naudojami besivystantys viščiukų embrionai, kurie yra užkrėsti šiuo virusu. Tam tikrą laiką inkubavus užkrėstus embrionus, jose dėl reprodukcijos susikaupęs virusas surenkamas, gryninamas (centrifuguojant ar kitu būdu) ir, jei reikia, inaktyvuojamas. Labai svarbu iš viruso preparatų pašalinti visas balasto priemaišas, kurios skiepijimo metu gali sukelti rimtų komplikacijų. Žinoma, ne mažiau svarbu įsitikinti, kad preparatuose nelieka neaktyvuoto patogeninio viruso. Pastaraisiais metais įvairios ląstelių kultūros buvo plačiai naudojamos virusams kaupti.
VIRUSŲ TYRIMO METODAI
Bakteriniai virusai pirmieji tapo išsamių tyrimų objektu kaip patogiausiu modeliu, kuris turi daug privalumų, palyginti su kitais virusais. Visas fago replikacijos ciklas, t.y. laikas nuo bakterinės ląstelės užkrėtimo iki dauginamų virusinių dalelių išsiskyrimo iš jos įvyksta per vieną valandą. Kiti virusai paprastai kaupiasi kelias dienas ar net ilgiau. Netrukus prieš Antrąjį pasaulinį karą ir netrukus po jo pabaigos buvo sukurti metodai atskiroms viruso dalelėms tirti. Plokštės su maistiniu agaru, ant kurių buvo užaugintas bakterijų ląstelių vienasluoksnis (ištisinis sluoksnis), yra užkrėstos fago dalelėmis, naudojant serijinius skiedimus. Daugindamasis virusas užmuša „apsaugotą“ ląstelę ir prasiskverbia į kaimynines ląsteles, kurios taip pat žūsta sukaupus fagų palikuonis. Negyvų ląstelių plotas plika akimi matomas kaip ryški dėmė. Tokios dėmės vadinamos „neigiamomis kolonijomis“ arba plokštelėmis. Sukurtas metodas leido ištirti atskirų viruso dalelių palikuonis, aptikti genetinę virusų rekombinaciją ir nustatyti genetinę fago replikacijos struktūrą ir detales, kurios anksčiau atrodė neįtikėtinos.
Darbas su bakteriofagais prisidėjo prie metodinio arsenalo išplėtimo tiriant gyvūnų virusus. Prieš tai stuburinių gyvūnų virusų tyrimai buvo atliekami daugiausia su laboratoriniais gyvūnais; tokie eksperimentai buvo labai sunkūs, brangūs ir nebuvo labai informatyvūs. Vėliau atsirado naujų metodų, pagrįstų audinių kultūrų naudojimu; fagų eksperimentuose naudojamos bakterijų ląstelės buvo pakeistos stuburinių ląstelėmis. Tačiau norint ištirti virusinių ligų vystymosi mechanizmus, eksperimentai su laboratoriniais gyvūnais yra labai svarbūs ir tebevykdomi šiuo metu.
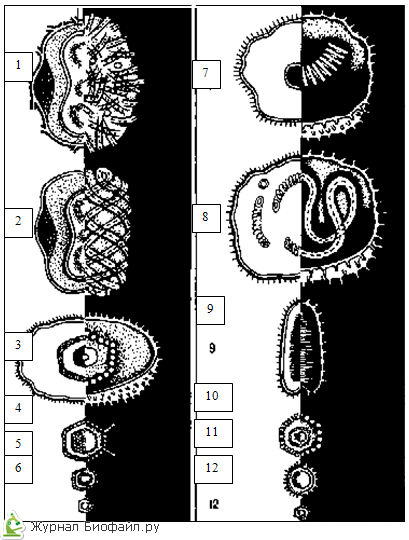
ICOSAHEDRIC SIMMETRIJOS TIPAS, parodytas adenoviruso, kapsomerų arba baltymo viruso subvienetų struktūros diagramoje, sudaro izometrinį baltymo apvalkalą, susidedantį iš 20 taisyklingų trikampių.
SPIRALINĖS SIMETRIJOS ATVEJU, parodyta tabako mozaikos viruso struktūros schemoje, kapsomerai ar viruso subvienetai sudaro spiralę aplink tuščiavidurį vamzdinį šerdį.
JUNGTINĖ arba mišri virusų simetrija gali būti pateikiama skirtingais variantais. Diagramoje parodyta bakteriofago dalelė turi taisyklingos geometrinės formos „galvutę“ ir „uodegą“ su spiralės simetrija.
Virusų kristalizacija
1932 m. Jaunam amerikiečių biochemikui Wendillui Stanley buvo paprašyta ištirti virusus. Stenlis pradėjo išspausdamas sulčių butelį iš tonos tabako lapų, užkrėstų tabako mozaikos virusu. Jis pradėjo tirti sultis naudodamas jam prieinamus cheminius metodus. Jis paveikė įvairias sulčių frakcijas įvairiais reagentais, tikėdamasis gauti gryną virusinį baltymą (Stanley buvo įsitikinęs, kad virusas yra baltymas). Kartą Stanley gavo beveik gryną baltymo dalį, kuri savo sudėtimi skyrėsi nuo augalų ląstelių baltymų. Mokslininkas suprato, kad prieš jį yra tai, ko jis taip atkakliai siekė. Stanley išskyrė nepaprastą baltymą, ištirpino jį vandenyje ir įdėjo tirpalą į šaldytuvą. Kitą rytą vietoj kolbos skaidrus skystis ten buvo gražūs šilkiniai adatos formos kristalai. Stanley gavo šaukštą šių kristalų iš tonos lapų. Tada Stanley išpylė keletą kristalų, ištirpino juos vandenyje, sudrėkino marlę šiuo vandeniu ir patrino ja sveikų augalų lapus. Augalų sultys patyrė daugybę cheminių poveikių. Po tokio „masinio apdorojimo“ virusai greičiausiai turėjo mirti.
Sutrinti lapai serga. Taigi, keistas viruso savybes papildė dar viena - galimybė kristalizuotis.
Kristalizacijos efektas buvo toks didžiulis, kad Stanley ilgą laiką atsisakė minties, kad virusas yra būtybė. Kadangi visi fermentai yra baltymai, daugybė fermentų taip pat didėja, kai organizmas vystosi ir jie gali kristalizuotis, Stanley padarė išvadą, kad virusai yra gryni baltymai, veikiau fermentai.
Netrukus mokslininkai įsitikino, kad įmanoma kristalizuoti ne tik tabako mozaikos, bet ir daugybę kitų virusų.
Po penkerių metų anglų biochemikai F. Bowdenas ir N. Peary rado klaidą Stanley apibrėžime: 94% tabako mozaikos viruso turinio sudarė baltymai, o 6% - nukleino rūgštis. Iš tikrųjų virusas buvo ne baltymas, o nukleoproteinas - baltymų ir nukleino rūgšties derinys.
Kai tik elektronų mikroskopai tapo prieinami biologams, mokslininkai nustatė, kad viruso kristalus sudaro keli šimtai milijardų dalelių, glaudžiai suspaustų. Viename poliomielito viruso kristale yra tiek dalelių, kad jos gali ne kartą užkrėsti visus Žemės gyventojus. Kai pavyko atskiras viruso daleles ištirti elektroniniame mikroskope, paaiškėjo, kad jos yra skirtingos formos, tačiau visada išorinis virusų apvalkalas susideda iš baltymų, kurie skiriasi skirtingais virusais, o tai leidžia juos atpažinti naudojant imunologines reakcijas, o vidinį turinį vaizduoja nukleino rūgštis, kuri yra paveldimumo vienetas.
Virusų komponentaiDidžiausi virusai (raupų virusai) savo dydžiu artėja prie mažų bakterijų, mažiausi (encefalito, poliomielito, snukio ir nagų ligos sukėlėjai) - prie didelių baltymų molekulių. Kitaip tariant, virusai turi savo milžinus ir nykštukus. (žr. 1 pav.) Virusams matuoti naudojama konvencija, vadinama nanometru (nm). Vienas nm yra milimetro milimetro dalis. Skirtingų virusų dydžiai svyruoja nuo 20 iki 300 nm.
Taigi, virusus sudaro keli komponentai:
šerdis - genetinė medžiaga (DNR ar RNR). Genetiniame viruso aparate yra informacijos apie kelis baltymų tipus, kurie reikalingi naujam virusui susidaryti.
baltyminis sluoksnis, vadinamas kapsidu. Vokas dažnai statomas iš vienodų pasikartojančių subvienetų - kapsomerų. Kapsomerai formuoja struktūras su dideliu simetrijos laipsniu.
Papildoma lipoproteinų membrana. Jis susidaro iš ląstelės-šeimininkės plazmos membranos. Jis randamas tik santykinai dideliuose virusuose (gripas, pūslelinė). Šis išorinis apvalkalas yra ląstelės-šeimininkės branduolinės arba citoplazminės membranos fragmentas, iš kurio virusas patenka į tarpląstelinę aplinką. Kartais išoriniuose sudėtingų virusų apvalkaluose be baltymų angliavandeniai yra, pavyzdžiui, gripo ir herpeso sukėlėjai.
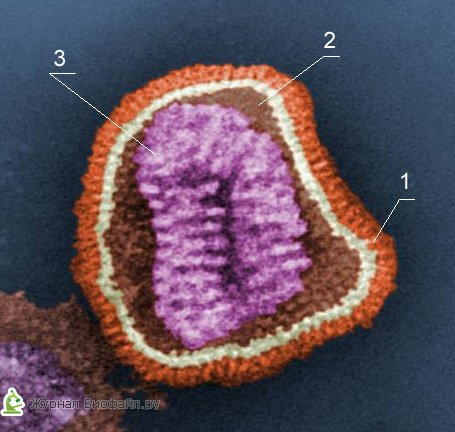
1. Papildomas apvalkalas
2. Kapsomeras (baltyminis kailis)
3. Šerdis (DNR arba RNR)
Kiekvienas virionų komponentas turi tam tikras funkcijas: baltymų apvalkalas apsaugo juos nuo neigiamo poveikio, nukleorūgštis yra atsakinga už paveldimas ir infekcines savybes ir vaidina pagrindinį vaidmenį virusų kintamumo atžvilgiu, o fermentai dalyvauja jų dauginimosi procese.
Struktūriškai sudėtingesniuose virusuose, be baltymų ir nukleorūgščių, yra angliavandenių ir lipidų. Kiekviena virusų grupė turi savo baltymų, riebalų, angliavandenių ir nukleorūgščių rinkinį. Kai kurių virusų sudėtyje yra fermentų.
Skirtingai nuo paprastų gyvų ląstelių, virusai nevartoja maisto ir negamina energijos. Jie negali daugintis be gyvos ląstelės dalyvavimo. Virusas pradeda daugintis tik patekęs į tam tikros rūšies ląsteles. Pavyzdžiui, poliomielito virusas gali gyventi tik žmonių ar labai organizuotų gyvūnų, pavyzdžiui, nervų ląstelėse beždžionė. Bakterinių virusų struktūra yra šiek tiek kitokia.
Viruso sąveika su ląsteleVirš ląstelės esantys virusai yra kristalai, tačiau patekę į ląstelę jie „atgyja“. Jų dauginimasis vyksta ypatingai, nepalyginamai. Pirmiausia virionai patenka į ląstelę ir išsiskiria viruso nukleorūgštys. Tuomet „surenkamos“ būsimų virionų detalės. Dauginimasis baigiasi surinkus naujus virionus ir išleidus juos į aplinką.
Virusų susitikimas su ląstelėmis prasideda jo adsorbcija, tai yra prisirišimu prie ląstelės sienos. Tada prasideda viriono įvedimas arba prasiskverbimas į ląstelę, kurį ji pati vykdo. Tačiau paprastai prieš viruso prasiskverbimą į ląstelės citoplazmą jis prisijungia prie specialaus receptoriaus baltymo, esančio ant ląstelės paviršiaus. Prisirišimas prie receptoriaus atliekamas dėl viruso dalelės paviršiuje esančių specialių baltymų, kurie „atpažįsta“ atitinkamą receptorių ant jautrios ląstelės paviršiaus. Dešimtys ir net šimtai virionų gali būti adsorbuoti vienoje ląstelėje. Ląstelės paviršiaus plotas, prie kurio prisijungė virusas, pasineria į citoplazmą ir virsta vakuole. Vakuolė, kurios siena susideda iš citoplazminės membranos, gali susilieti su kitomis vakuolėmis arba su branduoliu. Taip virusas patenka į bet kurią ląstelės dalį. Šis procesas vadinamas viropexis.
Infekcinis procesas prasideda, kai į ląstelę patekę virusai pradeda daugintis, t. įvyksta viruso genomo dubliavimasis ir savaiminis kapsidės susidarymas. Norint atsirasti reduplikacijai, nukleino rūgštis turi būti išskirta iš kapsidės. Sintetavus naują nukleino rūgšties molekulę, ji apsirengia, o ląstelės citoplazmoje sintetinami virusiniai baltymai suformuoja kapsidą. Virusinių dalelių kaupimasis veda prie jų išėjimo iš ląstelės. Kai kuriems virusams tai įvyksta „sprogus“, dėl to pažeidžiamas ląstelės vientisumas ir ji žūva. Kiti virusai išskiriami panašiai kaip pumpurai. Tokiu atveju kūno ląstelės gali išlaikyti savo gyvybingumą ilgą laiką.
Kitas būdas prasiskverbti į bakteriofagų ląstelę. Storos ląstelių sienelės neleidžia receptoriaus baltymui kartu su prijungtu virusu panirti į citoplazmą, kaip atsitinka, kai gyvūnų ląstelės yra užkrėstos. Todėl bakteriofagas į ląstelę įleidžia tuščiavidurę lazdelę ir per ją išstumia jos galvoje esančią DNR (arba RNR). Bakteriofago genomas patenka į citoplazmą, o kapsidas lieka lauke. Bakterinės ląstelės citoplazmoje prasideda bakteriofago genomo replikacija, jo baltymų sintezė ir kapsidės susidarymas. Praėjus tam tikram laikotarpiui, bakterijų ląstelė miršta, o brandžios fago dalelės patenka į aplinką.
Nuostabu, kaip dešimtys ir net šimtus kartų mažesnės už ląsteles virusai sumaniai ir užtikrintai valdo korinį ekonomiką. Daugindamiesi jie išeikvoja ląstelių išteklius ir giliai, dažnai negrįžtamai, sutrikdo medžiagų apykaitą, o tai galiausiai sukelia ląstelių mirtį.
Augalų virusų forma paprastai yra lazdelės formos ir apvali. Lazdelės formos virusų matmenys yra 300–480 x 15 nm, o apvalios formos - 25–30 nm. [...]
Tai yra mikroorganizmai, neturintys ląstelių struktūros. Virusų (virionų) struktūrinių vienetų dydžiai svyruoja nuo 10 iki 300 nm. Virionai apima ribonukleino (RNR) arba dezoksiribonukleino (DNR) molekules, apsuptas baltymų apvalkalo. Virusai yra įvairių formų: kubiniai, sferiniai, lazdelės formos ir kt. Virusų dauginimasis atliekamas paprastu dalijimu arba kompleksiškiau tik gyvo organizmo ląstelių viduje. Virusai turi veikimo specifiškumą, tai yra tam tikros virusų grupės užkrėsti tam tikrus gyvus organizmus.
Virusai, kurie yra mažesni ir mažiau sudėtingi nei ląstelės, negali gyventi savarankiškai. Tai tik labai savotiškai supakuoti genetinės informacijos gabalai, kurie gali gyventi ir daugintis tik užkrėtę ląstelę. Tokiu atveju vienoje ląstelėje gali susidaryti tūkstančiai virusinių dalelių. Daroma prielaida, kad virusai kažkaip pajungia ląstelių gyvenimo mechanizmą ir naudoja jį savo tikslams. Evoliucinė virusų kilmė nėra visiškai aiški. Jie gali būti laikomi labai sugeneruotomis ląstelėmis ar jų fragmentais. Virusų genai yra panašūs į kitų formų genus ir taip pat gali būti mutuoti. [...]
Šiame viruse yra apie 20% RNR, o jo dalelės yra daugiakampės. Elektroninių mikrografų dalelių skersmuo, gautas neigiamai kontrastuojant, yra nuo 26 iki 30 nm, o tai priklauso nuo preparato paruošimo savybių. Išsami šio viruso struktūra nėra aiški, tačiau jis kelia didelį susidomėjimą, nes kai kuriuose izoliatuose yra su šiuo virusu susijęs palydovinis virusas, aprašytas toliau, taip pat [...]
Kita priklausomybės forma būdinga tabako nekrozės viruso palydoviniam virusui. Tai mažiausias žinomas virusas. Jo RNR yra informacijos kiekis, pakankamas jo paties kapsidės baltymui ir, galbūt, specifinei RIK polimorazei koduoti. Kitoms reikšmingoms, bet iki šiol nežinomoms funkcijoms op priklauso nuo nesusijusio tabako nekrozės viruso buvimo. [...]
Viruso virionas yra srieginis, 600–700 x 12 mikronų dydžio, inaktyvuotas 60–67 ° C temperatūroje, atlaiko užšalimą. Vežėjas nežinomas. [...]
Liucernos mozaikos viruso (ALV) dalelės nuo kitų augalų virusų skiriasi savo baciliformos forma. Šių virusų struktūra turi keletą bruožų, būdingų ir lazdelės formos, ir izometriniams virusams. Iš VML viruso preparato buvo išskirti penki komponentai (b0, 1a, bb, M ir B). Nustatyta, kad mažiausiai keturi iš jų yra būtini infekcijos atsiradimui (sk. [...]
Mikrobų dydis ir forma. Bakterijų dydis svyruoja nuo dešimtųjų mikronų iki kelių mikronų. Vidutiniškai daugumos bakterijų kūno skersmuo yra 0,5-1 mikrono ribos, o vidutinis lazdelės formos bakterijų ilgis yra 1-5 mikronai. Šiuolaikinių bakteriologinių mikroskopų skiriamoji geba yra 0,2 mikronai. Todėl norint pamatyti ultramikrobus (virusus, bakteriofagus), reikia naudoti elektroninį mikroskopą, kuris gali padidinti tūrį milijonais kartų ir kurio skiriamoji geba yra 0,4 mmq. . [...]
Pavieniai vienos ar kitos rūšies virusai yra įvairios formos (apvalios, lazdelės ar kitokios formos) dariniai, kurių viduje yra baltymų apvalkale (kapsidėje) uždaryta nukleino rūgštis (DNR arba RNR). [...]
IN bendras vaizdas virusai yra submikroskopiniai dariniai, susidedantys iš baltymų ir nukleino rūgšties, organizuoti viruso dalelių pavidalu, dažnai vadinamais viruso dalelėmis, virionais, virosporomis ar nukleokapsidais. [...]
Kai kuriose elektronų mikrografijose jie pastebėjo beveik tokio paties skersmens diskoidines daleles kaip ir nepažeistas virusas. Šiose dalelėse buvo matomas centrinis kanalas, kurio skersmuo buvo įvairus, apsuptas 10 radialiai išsidėsčiusių subvienetų. [...]
Filtruojamos bakterijų formos skiriasi nuo filtruojamų virusų tuo, kad gali išsivystyti ir dirbtinėse maistinėse terpėse. [...]
Bakterijose yra 1–4% riebalų, 8–14% baltymų ir 80–85% vandens. Pėdsakuose yra fosforo, kalio, kalcio, magnio, geležies ir kitų elementų. Virusai neturi ląstelių struktūros ir jų dydis yra 10-100 nm. [...]
Ligos sukėlėjas yra Beapo geltonosios mozaikos virusas (Phaseolus virus 2 Smith). Virusas inaktyvuojamas 70 ° C temperatūroje. Tai veikia visus ankštinius augalus, negali būti perduodama su sėklomis. [...]
Sukėlėjai yra ara-bis mozaikos virusas ir aviečių žiedų polio virusas. Abu virusai priklauso tai pačiai grupei, turi izometrines daleles, kurių skersmuo yra maždaug: Yu im. Jie pernešami kontaktinėmis-mechaninėmis priemonėmis, dirvožemio nematodais ir skiepijant. Ant lapų yra šviesiai žalios arba gelsvos dėmės, skirtingo dydžio ir formos neaiškiais kraštais. Lapai yra maži, deformuoti, augalai yra prislėgti. Esant didelei žalai, imlių veislių augalai miršta per metus. [...]
NOLOGIJA bakterijų, virusų ir cheminių teršalų ekstrahavimui iš vandens ", kuri susideda iš to, kad per celiuliozės sorbentą praeinantys mikroorganizmai dėl elektrostatinės sąveikos„ įstringa "į sorbento struktūrą". Dėl to „vanduo 100% dezinfekuojamas nuo virusų, 100% beveik visų bakterijų ir 95–100% nuo E. coli bakterijų. Priemaišos iš vandens pašalinamos kompleksiškai: tai įvyksta dėl dalelių mechaninio sulaikymo akytoje filtro medžiagos struktūroje, dėl molekulinės sorbcijos, elektrostatinės sąveikos ir jonų mainų ". Asmeniškai aš nematau nieko iš esmės naujo šioje technologijoje, tačiau gyvybę teikiančiame šaltinyje yra vienas originalus momentas. Cituoju: "Viršutinė filtro dalis, esanti bažnyčios kupolo pavidalu, turi teigiamą energetinį ir psichologinį poveikį žmonėms, geriantiems išgrynintą vandenį". Tada seka lentelė, kurioje „Gyvybės dovanojimas“ lyginamas su visokiomis „akvaforomis“ ir „instapurais“ (sic), kuriomis, be abejo, jis nusišluosto nosį. [...]
Bakteriofagai ir filtruojami virusai neturi įprastos ląstelių struktūros, todėl organizuota ląstelė nėra paskutinis gyvenimo vienetas. Tai patvirtina faktai apie matomų bakterijų formų perėjimą prie „nematomų“, neląstelinės formos, vadinamos filtruojamomis matomų bakterijų formomis. [...]
Liucernos mozaika. Sukėlėjas yra liucernos mozaikos virusas (liucernos mozaikos virusas, Medicago virus 2 Smith). Jis perduodamas kontaktinėmis-mechaninėmis priemonėmis, amarais, sėklomis. Simptomai: pirmiausia ant lapų atsiranda mažos suapvalintos gelsvos dėmės, tada tarp šoninių venų yra pailgos ar netaisyklingos dėmės, išilgai gyslų - šviesiai geltonos arba balkšvos linijos. Lapai yra maži ir deformuoti. Vasarą simptomai dažnai užmaskuojami. Latentinė infekcija nėra reta. Virusas turi platų šeimininkų spektrą: jis užkrečia laukinius ir kultūrinius daugelio šeimų augalus: kandžius, Solanaceae, Asteraceae, moliūgus ir kt. [...]
Be ląstelių struktūros organizmų, yra ir neląstelinės gyvybės formos - virusai ir bakteriofagai. Beje, virusus 1892 metais atrado rusų biologas D.I. Ivanovas, o jų vardas vertime reiškia „nuodai“, kurie apskritai, daugeliui žmonių įprastai vartojami, atspindi jų poveikį sveikatai. [...]
Nėra aiškios ribos tarp gyvų ir negyvų medžiagų, tai patvirtina virusų egzistavimas. Pastarieji turi tiek gyvų, tiek negyvų požymių. Visuotinai priimtas jų apibrėžimas dar nėra suformuluotas. Paprastai manoma, kad virusai yra mažiausiai organizuotos gyvenimo formos, neturinčios savo metabolizmo ir galinčios egzistuoti tik kitų organizmų ląstelėse. Jie nesidaugina už ląstelių ribų. Tuo pačiu virusų gebėjimas daugintis, net jei jie liečiasi su kitomis ląstelėmis, yra gyvų būtybių ženklas. [...]
Dirvožemyje yra įvairių mikroorganizmų: bakterijų, aktinomicetų ar spinduliuojančių grybų, grybų, virusų ir kt. Dauguma jų apdoroja miško paklotę (humuso sluoksnį), pagerina dirvožemio struktūrą ir organinius junginius paverčia virškinamomis formomis. Padidėjus dirvožemio rūgštingumui ir formuojantis tirpioms nuodingų metalų formoms, sumažėja mikroorganizmų aktyvumas, ypač perdirbant miško paklotę. [...]
Elektrolizės produktų ir chloro inaktyvuojančiam poveikiui didelę įtaką daro chloro (laisvo arba surišto) kiekis ir forma. Ištyrus modelio poliomielito viruso, Escherichia coli ir Escherichia coli fago inaktyvavimo elektrolizės produktais ir chloro dinamiką, nustatyta, kad esant tik likusio chloro būsenai surištoje būsenoje, 30 minučių kontakto metu Escherichia coli visiškai mirė, o fago virusas - atitinkamai tik 80 ir 60%. Likus laisvam chloro likučiams, praėjus 20 minučių kontakto, E. coli ir fagą inaktyvavo daugiau nei 99%, o virusą - tik 90%. Kai laisvo likučio chloro kiekis vandenyje yra 0,1–0,3 mg / l, 10 minučių kontakto metu jis buvo visiškai dezinfekuotas E. coli ir fago atžvilgiu, o 30 minučių buvo nustatytas tik nereikšmingas aktyvių virusų kiekis. Tirtų mikroorganizmų inaktyvacijos laipsnio skirtumas visais atvejais buvo statistiškai reikšmingas. Išbandytomis sąlygomis dezinfekuojant elektrolizės produktais ir vandens chloru, kuriame yra vienodos koncentracijos mikroorganizmai, Escherichia coli buvo mažiau atsparus nei fagas, o fagas buvo mažiau atsparus nei virusas. Todėl E. coli ir fagas gali būti patikimi sanitariniai efektyvaus vandens dezinfekavimo elektrolizės produktais ir chloru prieš enterovirusus rodikliai. Iš esmės tai reiškia tuos atvejus, kai dėl nepalankių epideminių sanitarinių sąlygų enterovirusų koncentracija rezervuarų vandenyje gali žymiai padidėti ir pasiekti E. coli lygį (E. L. Lovtsevich, L. A. Sergunina, 1968). [...]
Taigi, išradus antibiotikus, pagrindinis žmogaus priešas buvo ne paprasčiausi grybai ir vienaląsčiai, o virusai. Yra pirmieji simptomai, kad retrovirusai pakeičia virusus - ikivirusinės, senoviškesnės gyvybės formos, jų organizacija kuriama ne pagal DNR molekulę, o pagal RNR. Vienas garsiausių šios gyvybės formos atstovų yra AIDS retrovirusas. [...]
Mikroskopu nematomi mikroorganizmai vadinami ultra mikrobais. Iš šios ultramikroskopinių formų grupės žmogaus praktikoje svarbiausi bakteriofagai - filtruojami virusai ir nematomos bakterijų formos. Ultramikrobus buvo galima stebėti tik elektroniniame mikroskope, o tai padidina iki 45 000 kartų. Virusai (85 pav.) Yra dalelės, susidedančios iš baltymų ir nukleino rūgšties (DNR ar RNR). Jie neturi įprastos ląstelių struktūros. Į neląstelinę gyvybės formą įeina ir bakteriofagai (86 pav.), Kurie yra pailgi dariniai sustorėjusiu galu. [...]
Infekcinis procesas yra makroorganizmo reakcijų kompleksas, atsirandantis reaguojant į jame esančių mikrobų, virusų ir kt. Įvedimą ir dauginimąsi. Tai ne visada lydi ligos požymių buvimas. Pavyzdžiui, esant mikrobinei ar besimptomei infekcijai, klinikinių požymių nėra, nors jo sukėlėjas yra organizme ir veikia įvairias jo sistemas, todėl pastaroji imunologiškai pertvarkoma. Jei infekcinį procesą lydi klinikinių požymių pasireiškimas, tai ši infekcijos forma vadinama infekcine liga. Todėl infekcinė liga yra vadinamoji akivaizdi infekcijos forma. [...]
Šiems virusams būdingas morfologinių charakteristikų panašumas, reakcijos į elektromagnetinę spinduliuotę, reprodukcija ir kt. Pagrindiniai jų komponentai yra: C, H, N, P, O, angliavandeniai ir lipidai. Yra žinoma, kad Prak-Pries ir visi onkovirusai yra termiškai nestabilūs ir sunaikinami 50–70 ° C temperatūroje, atsižvelgiant į onkoviruso tipą. [...]
Šiai grupei priklauso koloidinės (mineralinės ir organomineralinės) dirvožemio ir grunto dalelės, taip pat neatsiskyrusios ir netirpios humuso medžiagos, suteikiančios vandeniui spalvą. Pastarieji iš miško, pelkėtų ir durpinių dirvožemių išplaunami į natūralius vandens telkinius, taip pat dėl \u200b\u200bvandens augalų ir dumblių gyvenimo susidaro pačiuose vandens telkiniuose. Ši grupė taip pat gali apimti virusus ir kitus organizmus, artėjančius koloidinių dalelių dydžiui. Kadangi tarp jų yra patogeninių (patogeninių) organizmų, jų pašalinimas iš vandens yra labai atsakingas įvykis. [...]
Antroji priemaišų grupė sujungia hidrofilines ir hidrofobines mineralines ir organominerines koloidines dirvožemio ir grunto daleles, neatsiskyrusias ir netirpias didelės molekulinės masės humusinių medžiagų ir ploviklių formas. Kinetinis hidrofobinių priemaišų stabilumas apibūdinamas gravitacijos lauko jėgų ir Brauno judesio santykiu; jų visuminį stabilumą lemia sąsajos elektrostatinė būsena ir difuzinių sluoksnių susidarymas arba stabilizuojančių sluoksnių susidarymas ant dalelės paviršiaus. Šiai grupei taip pat priklauso virusai ir kiti mikroorganizmai, panašaus dydžio į koloidines daleles. [...]
Augalų virusams išskirti ir kiekybiškai įvertinti galima naudoti Bracke tankio gradiento centrifugavimo metodą. Kaip paaiškėjo, šis metodas yra kupinas daugybės galimybių ir šiuo metu yra plačiai naudojamas virusologijos ir molekulinės biologijos srityse. Atliekant tyrimus centrifuguojant tankio gradiente, centrifugos mėgintuvėlis iš dalies užpildomas tirpalu, kurio tankis mažėja nuo dugno iki menisko. Sacharozė dažniausiai naudojama augalų virusų frakcionavimo gradientui sukurti. Prieš centrifugavimą viruso dalelės gali pasiskirstyti visame tirpalo tūryje arba padengti gradiento viršų. Bracke'as pasiūlė tris skirtingus tankio gradiento centrifugavimo metodus. Izopikipiniame (pusiausvyros) centrifugavimo procese procesas tęsiasi tol, kol visos gradiento dalelės pasiekia lygį, kuriame terpės tankis yra lygus jų pačių tankiui. Taigi dalelių frakcionavimas šiuo atveju vyksta atsižvelgiant į jų tankio skirtumus. Sacharozės tirpalai nėra pakankamai tankūs izopikniškam daugelio virusų atskyrimui. Vykdant didelę spartą zonoje centrifuguojant, virusas pirmiausia taikomas anksčiau sukurtam gradientui. Kiekvieno tipo dalelės tuo pačiu metu nusodinamos per gradientą zonos arba juostos pavidalo greičiu, priklausomai nuo jų dydžio, formos ir tankio. Šiuo atveju centrifugavimas nutraukiamas, kol dalelės toliau nuosėdose. Pusiausvyros zoninis centrifugavimas yra panašus į greitą zoninį centrifugavimą, tačiau šiuo atveju centrifugavimas tęsiasi tol, kol pasiekiama izopikninė būsena. Tankio gradiento vaidmuo centrifuguojant dideliu greičiu yra slopinti konvekciją ir fiksuoti įvairių rūšių molekules specifinėse zonose. Tankio gradiento centrifugavimo teorija yra sudėtinga ir nėra gerai suprantama. Tačiau praktiškai tai yra paprastas ir elegantiškas metodas, plačiai naudojamas dirbant su augalų virusais. [...]
Pagrindinis matricoje lokalizuoto AHC CEC (kaip ir ląstelėje esančių onkovirusų) bruožas yra sąsajos tarp dviejų skirtingų laidumo terpių buvimas. Fig. 2.11 rodo elektronų mikroskopijos duomenis, rodančius adenovirusą, epstein-Barr virusas (EBV) ir TsECh ShchGK. Pav. 2.11 matyti, kad visi tos pačios skalės dariniai turi artimą sferinei formai, susidedantys iš šerdies ir apvalkalo, cheminė sudėtis kiekviename apvalkale yra elektrai aktyvių jonų, aiškiai pažymėtos virusų ir CEC ribos su jų matricomis. [...]
Anksti pavasarį jis pasirodo ant lapų kaip geltonas taškelis. Iki vasaros vidurio šis simptomas išnyksta, tačiau paveikti lapai kartais raukšlėjasi. Vaisiai yra maži, dažnai netaisyklingi ir su gumbais išilgai siūlės. Jų brendimas vėluoja. Ligos sukėlėjas - persikų mozaikos virusas perduodamas skiepijant ir pradedant pumpurais. Manoma, kad slyvų amarai neša virusą. [...]
Nepaisant daugybės atradimų, biogenezės vaizde vis dar yra daugybė tuščių vietų. Tik pagrindiniai etapai gali būti laikomi neginčijamais. Taigi dabar nėra jokių abejonių, kad biosferos atsiradimas buvo išskirtinis, vienintelis įvykis. Mažas virusas ir milžiniškas jūros monstras, vienaląsčiai dumbliai ir medinis papartis, kurie dingo prieš milijonus metų, visi yra tik to paties filogenetinio medžio šakos ir lapai. Gyvybės formos visada ir visur atskleidžia, galima sakyti, „kraujo giminystę“, o visi jos vaikai yra genetiškai susiję. Nuo tos dienos, kai Žemėje atsirado pirmasis padaras, gyvenimas ateina tik iš gyvenimo. [...]
Ląstelė yra pagrindinis visų gyvųjų organizmų struktūrinis ir funkcinis vienetas, elementari gyvoji sistema. Jis gali egzistuoti kaip atskiras organizmas (bakterijos, pirmuonys, kai kurie dumbliai ir grybai) ir kaip daugialąsčių organizmų audinių dalis. Tik virusai yra neląstelinės gyvybės formos. [...]
Pagal siūlomą schemą, pirmame proceso etape susidaro restrikcijos endonukleazės EcoR I fermento-substrato kompleksas EI ir dvigubos grandinės plazmidės DNR. Pagrindinis schemos taškas yra restrikcijos endonukleazės „EcoR I“ kompleksinio E-II susidarymas su žiedine DNR forma, turinčia vienos grandinės pertrauką, gautą hidrolizuojant fosfodiesterio ryšį vienoje iš DNR grandinių. Be to, priklausomai nuo sąlygų (substrato pobūdžio, temperatūros ir kt.) ) arba antrosios DNR grandinės skilimas tame pačiame E - II komplekse gali atsirasti, susidarant E-III fermento kompleksui su linijine DNR forma, arba E-P kompleksui atsiribojus, susidarant laisvam fermentui ir žiedinei DNR, turinčiai vienos grandinės pertrauką, kuri sukelia II formos kaupimasis tirpale. Ši schema leido paaiškinti, viena vertus, SV 40 viruso DNR ir ColE I bei bakteriofago G4 DNR hidrolizės mechanizmų skirtumus. SV 40 viruso DNR atveju vyksta E-11 fermento-substrato komplekso disociacija, dėl ko tirpale kaupiasi žiedinė DNR forma. Buvo pasiūlyta, kad šių DNR molekulių (virusas SV 40; DNR ColE I ir bakteriofagas G4) hidrolizės mechanizmo skirtumai yra „EcoR I“ restrikcijos fermento sąveikos su įvairiomis nukleotidų sekomis, esančios šalia „EcoR I“ restrikcijos fermento atpažinimo vietos, rezultatas. Tačiau ši prielaida neleidžia paaiškinti skirtumų. žiedinės DNR ColE I hidrolizės mechanizme, priklausomai nuo temperatūros (žr. aukščiau). [...]
Ši liga yra žinoma daugelyje pasaulio šalių. SSRS jis buvo rastas Ukrainoje, Moldovoje, Estijoje ir Gruzijoje ir jam taikomas vidinis karantinas. Pažeistos slyvos, vyšnių slyvos, mirabelė, abrikosai ir persikai. Ligos sukėlėjas yra slyvų raupai (\u003d Prunus virus 7 Smith). Viruso forma yra srieginė, dydis 760X20 pavadintas [...]
Tolesnis plazmidinės DNR hidrolizės mechanizmo tyrimo restrikcijos endonukleazėmis tyrimo, gauto Halfordo ir kt. ... Taigi reakcijos mechanizmas yra panašus į tą, kuris siūlomas restrikcijos fermentu „EcoI I.“ hidrolizuojant ES 40 viruso DNR. [...]
Be pirmiau minėtų „bendrųjų“ funkcijų, kūno homeostazės buvimo, yra dar vienas labai svarbus bruožas: gyvoji medžiaga tarsi sukuria dar vieną buveinę, būtent galimybę nusodinti kūną su kitomis gyvomis būtybėmis nuolatiniam ar laikinam gyvenimui. Tai yra nauja gyvenimo sukelta biotinė buveinė. Daugelis ekspertų virusus laiko būtybėmis, gyvenančiomis šioje aplinkoje. Taigi, I.A. Shilov (2000) mano, kad išskirtinis jų struktūros paprastumas yra antraeilis reiškinys, dar labiau tikėtina, kad tai naujai atsiradusi gyvų būtybių forma, visiškai įsisavinusi kitų lygių organizmų tarpląstelinę aplinką. Antrasis šios tezės patvirtinimas yra tas, kad virusai turi didelį genetinės sistemos sudėtingumą ir įvairovę. Struktūros supaprastinimas, kuris tapo įmanomas dėl privalomų besąlyginių virusų ryšių su organizmu-šeimininku, užtikrinančiu stabilias gyvenimo sąlygas, paveikė net pagrindines savybes, būdingas didžiajai daugumai gyvybės formų: virusai neturi dirglumo ir neturi savo baltymų sintezės aparato. Virusai negali savarankiškai egzistuoti, o jų ryšys su ląstele yra ne tik erdvinis, bet ir standus funkcinis ryšys, su kuriuo ląstelė ir virusas reiškia tam tikrą vienybę. [...]
Trumpalaikis šarminis HPLC apdorojimas 30 ° C temperatūroje ir esant didelei jonų stipriai in situ sukelia pertraukas, dėl kurių susidaro gana vienodo dydžio RNR fragmentai, kuriuose s2 [...]
Maliarija, hepatitu, ŽIV ir daugeliu kitų ligų paveiktų žmonių skaičius yra didžiulis. Daugelis gydytojų mano, kad reikia kalbėti ne apie „pergalę“, o tik apie laikiną sėkmę kovojant su šiomis ligomis. Kovos istorija užkrečiamos ligos yra labai trumpas, o aplinkos pokyčių nenuspėjamumas (ypač mieste) gali paneigti šiuos laimėjimus. Dėl šios priežasties infekcinių agentų „grįžimas“ fiksuojamas tarp virusų. Daugelis virusų „atitrūksta“ nuo natūralios bazės ir pereina į naują etapą, kuris gali gyventi žmogaus aplinkoje - jie tampa gripo, virusinio vėžio ir kitų ligų sukėlėjais. Galbūt ši forma yra ŽIV. [...]
Svorio vidutinės molekulinės masės ir sukimosi spindulio pokytis buvo vertinamas remiantis šviesos sklaidos duomenimis. Taikant HPMC RNR, naudojant abu skaidymo metodus, jie nustatė, kad sukimosi spindulys padidėjo prieš prasidedant intensyviam molekulės skaidymui, tuo tarpu sukimosi spindulys ir vidutinė TMV RNR molekulinė masė sumažėjo nuo pat šio proceso pradžios. Strazielli ir kt. Paaiškino šią išvadą siūlydami, kad HPMC RNR yra uždaros kilpos pavidalu. Tačiau šiuos rezultatus galima interpretuoti kitaip. Pavyzdžiui, Hazelkornas parodė, kad TMV RNR ir HPMT RNR kartu nusėdo pH ir jonų stiprumo sąlygomis, panašiomis į tas, kurias naudojo Strazielli ir kt. Priešingai, fago cpX174 žiedinės ir plačios formos DNR formos yra lengvai atskiriamos dėl jų sedimentacinių savybių [1 515]. Remdamasis sedimentacijos duomenimis įvairiomis sąlygomis, Kuiperis pasiūlė, kad izoliuota agurkų mozaikos viruso (Y padermės) RNR gali egzistuoti dviem pavidalais: atvira grandinė ir žiedinė struktūra. Tačiau šiuos duomenis, kaip ir aukščiau aprašytą atvejį, galima paaiškinti įvairiai. [...]
Bakterijų DNR yra labai polimerinis junginys, susidedantis iš daugybės nukleotidų - polinukleotidų, kurių molekulinė masė yra apie 4 milijonai.DNR molekulė yra nukleotidų grandinė, kur jų vieta turi specifinę seką. Azoto bazių išsidėstymo sekoje yra užkoduota kiekvienos rūšies genetinė informacija. Pažeisti šią seką galima esant natūralioms mutacijoms arba veikiant mutageniniams veiksniams. Tokiu atveju mikroorganizmas įgyja ar praranda bet kokią savybę. Jis paveldėjo bruožų pokyčius, tai yra, atsiranda nauja mikroorganizmų forma. Visuose mikroorganizmuose - prokariotuose ir eukariotuose - genetinės informacijos nešėjai yra nukleorūgštys - DNR ir RNR. Tik keli virusai yra išimtis: jie neturi DNR, o paveldima informacija įrašoma arba atspindima tik RNR.