Les maladies virales sont apparues dans les temps anciens, mais la virologie en tant que science a commencé à se développer à la fin du 19ème siècle.
En 1892, le scientifique-botaniste russe D.I.Ivanovsky, étudiant la maladie de la mosaïque des feuilles de tabac, a découvert que cette maladie est causée par les plus petits micro-organismes qui passent à travers des filtres bactériens à pores fins. Ces micro-organismes sont appelés virus filtrables (du latin virus - poison). Plus tard, il a été montré qu'il y avait d'autres micro-organismes passant à travers des filtres bactériens, de sorte que les virus filtrés ont commencé à être appelés simplement virus.
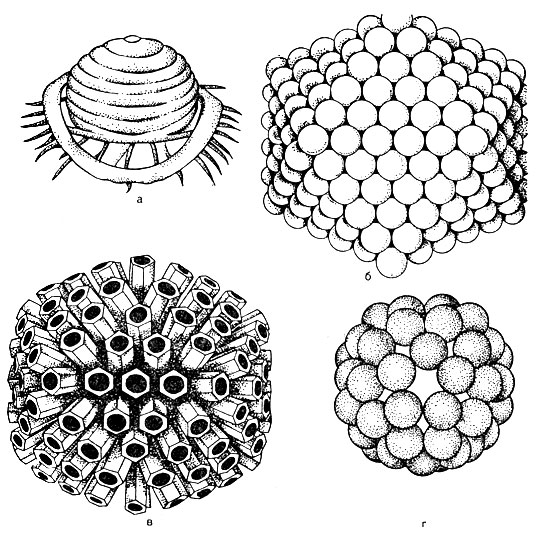
En général, la forme du virion et la présence ou l'absence d'une enveloppe en disent peu sur les maladies que les virus peuvent provoquer ou sur les espèces qu'ils peuvent infecter, mais restent un moyen utile de commencer la classification virale. Figure 5 Les virus peuvent être de forme complexe ou relativement simples.
Laquelle des affirmations suivantes concernant la structure du virus est correcte? Le noyau du virus contient le génome ou le contenu génétique général du virus. Les génomes viraux sont généralement petits par rapport aux bactéries ou aux eucaryotes, qui ne contiennent que des gènes codant pour des protéines que le virus ne peut pas obtenir de la cellule hôte. Ce matériel génétique peut être simple brin ou double brin. Il peut également être linéaire ou circulaire. Alors que la plupart des virus contiennent un segment d'acide nucléique, d'autres ont des génomes composés de plusieurs segments.
Une grande contribution à l'étude des virus a été apportée par les virologues soviétiques: M.A.Morozov, N.F. Gamaleya, L.A. Zilber, M.P. Chumakov, A.A. Smorodintsev, V.M. Zhdanov et d'autres.
Les virus sont forme non cellulaire existence de matière vivante. Ils sont très petits. Selon l'expression figurative de VM Zhdanov, «leur taille par rapport à la taille moyenne des bactéries peut être comparée à la taille d'une souris par rapport à un éléphant». Il n'est devenu possible de voir des virus qu'après l'invention du microscope électronique.
Les virus peuvent être considérés comme obligatoires parasites intracellulaires... Le virus doit s'attacher à une cellule vivante, l'ingérer, produire ses protéines et copier son génome, et trouver un moyen de s'échapper de la cellule afin que le virus puisse infecter d'autres cellules et finalement d'autres personnes. Les virus ne peuvent infecter que certains types d'hôtes et seulement certaines cellules de cet hôte. La base moléculaire de cette spécificité est qu'une molécule de surface spécifique connue sous le nom de récepteur viral doit être trouvée à la surface de la cellule hôte pour que le virus se fixe.
Actuellement, de nombreuses méthodes sont utilisées pour étudier les virus: chimiques, physiques, biologiques moléculaires, immunobiologiques et génétiques.
Tous les virus sont subdivisés en infectant les humains, les animaux, les insectes, les bactéries et les plantes.
Les virus ont une grande variété de formes et de propriétés biologiques, mais ils ont tous des caractéristiques structurelles communes. Les particules virales matures sont appelées virions.
En outre, les différences métaboliques observées dans différents types de cellules basées sur l'expression différentielle des gènes sont un facteur probable dans lequel les cellules peuvent utiliser le virus pour la réplication. La cellule doit constituer les substances nécessaires au virus, telles que les enzymes, dans lesquelles le génome viral lui-même ne contient pas de gènes, ou le virus ne peut pas se répliquer en utilisant cette cellule.
Le virus doit "prendre en charge" la cellule pour la réplication. Le cycle de réplication virale peut conduire à des changements biochimiques et structurels dramatiques dans la cellule hôte qui peuvent causer des dommages cellulaires. Ces changements, appelés effets cytopathiques, peuvent altérer la fonction cellulaire ou même détruire la cellule. Certaines cellules infectées, telles que celles infectées par le virus du rhume commun, meurent par lyse ou apoptose, libérant simultanément tous les virions de la descendance. Symptômes maladies virales sont le résultat d'une réponse immunitaire à un virus qui tente de contrôler et d'éliminer le virus de l'organisme, ainsi que des dommages cellulaires causés par le virus.
Contrairement à d'autres micro-organismes contenant à la fois de l'ADN et de l'ARN, le virion ne contient qu'un seul des acides nucléiques - soit l'ADN ou l'ARN.
L'acide nucléique des virus peut être simple brin et double brin. Presque tous les virus contenant de l'ARN ont un ARN simple brin dans leur génome, et ceux contenant de l'ADN ont un ADN double brin. Conformément à deux types de substances génétiques, les virus sont divisés en ARN et ADN. Les familles contenant de l'ADN comprennent 5 familles, celles contenant de l'ARN - 10 familles.
Pendant le processus de bourgeonnement, la cellule n'est pas lysée ou tuée immédiatement. La plupart des infections virales productives suivent des étapes similaires dans le cycle de réplication virale: nidification, pénétration, non-couverture, réplication, assemblage et libération. Le virus se fixe à un site récepteur spécifique sur la membrane de la cellule hôte via des protéines de fixation dans la capside ou des protéines intégrées dans son enveloppe. L'attachement est spécifique, et habituellement le virus ne s'attachera qu'aux cellules d'une ou plusieurs espèces et seulement à certains types de cellules au sein de ces espèces avec les récepteurs correspondants.
* (Voici des données sur quelques-uns seulement des virus pathogènes pour l'homme.)
Structure du virion... Au centre du virion se trouve un acide nucléique, qui est entouré d'une capside (du grec kanca - boîte). La capside est composée de sous-unités protéiques appelées capsomères. Un virus mature est une nucléocapside de structure chimique. Le nombre de capsomères et la manière dont ils sont conditionnés (Fig. 52) sont strictement constants pour chaque type de virus. Par exemple, le virus de la poliomyélite contient 32 capsomères, tandis que l'adénovirus contient 252 capsomères. Les capsomères peuvent être empilés sous la forme d'un polyèdre avec des bords symétriques uniformes - une forme cuboïdale (par exemple, un adénovirus). Le style sphérique est typique des virus grippaux. Il peut y avoir un type de symétrie dans lequel l'acide nucléique a la forme d'un ressort autour duquel des capsomères sont déposés, auquel cas le virus a une forme en forme de bâtonnet - un virus qui provoque la maladie des feuilles du tabac.
Contrairement aux virus animaux, l'acide nucléique du bactériophage est introduit dans la cellule hôte nue, laissant la capside à l'extérieur de la cellule. Les virus végétaux et animaux peuvent pénétrer dans les cellules par endocytose, dans laquelle la membrane cellulaire entoure et engloutit tout le virus. Certains virus enveloppés pénètrent dans la cellule lorsque l'enveloppe virale fusionne directement avec la membrane cellulaire. La capside virale se décompose ensuite à l'intérieur de la cellule et l'acide nucléique viral est libéré, qui devient alors disponible pour la réplication et la transcription.
Le mécanisme de réplication dépend du génome viral. Il y a, bien sûr, des exceptions à ce modèle. Si la cellule hôte ne fournit pas les enzymes nécessaires à la réplication virale, les gènes viraux fournissent des informations pour la synthèse directe des protéines manquantes.
Le phage a un type complexe de symétrie: la tête est cuboïde et le processus est en forme de bâtonnet (en forme de sperme) (voir Fig. 21, 22).
Ainsi, selon la méthode d'emballage, les virus sont subdivisés en formes cuboïde, sphérique, en forme de bâtonnet et spermatozoïde.
La dernière étape de la réplication virale est la libération de nouveaux virions dans l'organisme hôte, où ils sont capables d'infecter les cellules voisines et de répéter le cycle de réplication. Certains virus sont libérés lorsque la cellule hôte meurt, tandis que d'autres virus peuvent quitter les cellules infectées en s'échappant à travers la membrane sans tuer la cellule directement.
Figure 6 Lorsqu'elles sont infectées par le virus de la grippe, les glycoprotéines se fixent à la cellule épithéliale hôte. En conséquence, le virus est absorbé. Figure 7 Les virus causent une variété de maladies chez les humains, allant des maladies bénignes aux maladies graves.
Certains virus aux structures plus complexes ont un shell appelé peplos. Il se forme lorsque le virus quitte la cellule hôte. Dans ce cas, la capside virale est enveloppée par la surface interne de la membrane cytoplasmique de la cellule hôte et une ou plusieurs couches de l'enveloppe de la supercapside sont formées. Seuls certains virus ont une telle enveloppe, par exemple les virus de la rage, de l'herpès, de l'encéphalite. Cette coquille contient des phospholipides, qui sont dégradés par l'éther. Ainsi, en agissant sur l'éther, il est possible de distinguer un virus à péplos d'un virus à "capside nue".
Le vaccin peut être préparé en utilisant des virus vivants atténués, des virus tués ou des sous-unités moléculaires du virus. En général, les virus vivants conduisent à une immunité accrue, mais ont la capacité de provoquer des maladies à une fréquence faible. Le vaccin viral tué et les virus sous-unitaires sont incapables de provoquer la maladie, mais entraînent généralement une immunité moins efficace ou à long terme.
Les vaccins à virus vivants atténués sont conçus en laboratoire pour provoquer peu de symptômes chez les receveurs tout en leur procurant une immunité contre les infections futures. La poliomyélite est l'une des maladies qui a marqué une étape importante dans l'utilisation des vaccins. Le succès du vaccin contre la polio a ouvert la voie à la distribution systématique de vaccins infantiles contre la rougeole, les oreillons, la rubéole, la varicelle et d'autres maladies. Les vaccins vivants sont généralement produits en affaiblissant le virus de type sauvage en le cultivant en laboratoire dans des tissus ou à des températures différentes de celles auxquelles le virus est habitué chez l'hôte.
Dans certains virus, des capsomères sous forme d'épines (ces épines sont émoussées) dépassent de la couche lipidique externe de l'enveloppe. Ces virus sont appelés peplomers (par exemple virus de la grippe, voir Fig. 52).
L'acide nucléique du virus est porteur de propriétés héréditaires, et la capside et l'enveloppe externe ont des fonctions protectrices, comme pour protéger l'acide nucléique. De plus, ils facilitent l'entrée du virus dans la cellule.
Par exemple, le virus peut être cultivé dans des cellules in vitro, des embryons d'oiseaux ou d'animaux vivants. L'adaptation à ces nouvelles cellules ou à la température provoque des mutations dans les génomes du virus, leur permettant de mieux se développer en laboratoire tout en interférant avec leur capacité à provoquer des maladies lorsqu'elles sont réintroduites dans une condition trouvée chez l'hôte. Ces virus atténués provoquent donc toujours des infections, mais ils ne se développent pas très bien, ce qui permet aux réponses immunitaires d'évoluer dans le temps pour éviter des maladies graves.
Le risque d'utiliser des vaccins vivants, qui sont généralement plus efficaces que les vaccins tués, est un risque faible mais significatif que ces virus reviennent à leur forme pathogène par des mutations inverses. Des mutations inverses se produisent lorsqu'un vaccin mute chez un hôte de sorte qu'il se lit à l'hôte et peut provoquer à nouveau une maladie, qui peut ensuite se propager à d'autres personnes lors d'une épidémie.
Tailles de virus... Les virus sont mesurés en nanomètres. Leur taille fluctue dans une large gamme de 15-20 à 350-400 nm.
Méthodes de mesure des virus: 1) filtration à travers des filtres bactériens avec une taille de pore connue; 2) ultracentrifugation - les gros virus précipitent plus rapidement; 3) photographier des virus au microscope électronique.
Dans la grippe, une mutation dans les gènes des molécules de surface aide le virus à échapper à l'immunité protectrice qu'il aurait pu recevoir lors de la saison grippale précédente, ce qui oblige les gens à se faire vacciner chaque année. D'autres virus, tels que ceux qui causent des maladies infantiles, la rougeole, les oreillons et la rubéole, mutent si peu que le même vaccin est utilisé année après année.
Vaccins et médicaments antiviraux pour le traitement
Dans certains cas, les vaccins peuvent être utilisés pour traiter une infection virale active. Dans le cas de la rage, maladie neurologique mortelle transmise dans la salive de virus contaminés par la rage, la progression de la maladie entre le moment de la morsure de l'animal et le moment où il pénètre dans le système nerveux central peut prendre deux semaines ou plus. C'est assez de temps pour vacciner une personne soupçonnée d'avoir été mordue par un animal enragé, et la réponse immunitaire accrue de la vaccination est suffisante pour empêcher le virus de pénétrer dans les tissus nerveux.
La composition chimique des virus... La quantité et le contenu des virus à ADN et à ARN ne sont pas les mêmes. Dans l'ADN, le poids moléculaire varie de 1 · 10 6 à 1,6 · 10 8, et dans l'ARN - de 2 · 10 6 à 9,0 · 10 6.
Les protéines des virions se trouvent en nombre insignifiant, elles se composent de 16 à 20 acides aminés. En plus des protéines de capside, il existe également des protéines internes associées à l'acide nucléique. Les protéines déterminent les propriétés antigéniques des virus et, en raison de l'empilement dense des chaînes polypeptidiques, protègent le virus de l'action des enzymes de la cellule hôte.
De cette manière, les conséquences neurologiques fatales de la maladie sont évitées et l'individu n'a plus qu'à se remettre de la morsure infectée. Cette approche est également utilisée pour traiter Ebola, l'un des virus les plus rapides et les plus meurtriers qui infectent les humains, bien qu'il infecte généralement des populations limitées. Ebola est également la principale cause de mortalité chez les gorilles. En utilisant des vaccins nouvellement développés qui améliorent la réponse immunitaire, on espère que le système immunitaire des personnes touchées contrôlera mieux le virus, réduisant potentiellement la mortalité.
Les lipides et les glucides se trouvent dans l'enveloppe externe des virions complexes. La membrane de la cellule hôte est la source de lipides et de glucides. Les polysaccharides qui composent certains virus déterminent leur capacité à provoquer une agglutination des érythrocytes.
Enzymes virales... Les virus n'ont pas leur propre métabolisme, ils n'ont donc pas besoin d'enzymes métaboliques. Cependant, certains virus se sont avérés avoir des enzymes qui facilitent leur pénétration dans la cellule hôte. Par exemple, dans le virus de la grippe A, la neuraminidase a été trouvée, qui clive l'acide neuraminique contenu dans les membranes des cellules animales (érythrocytes, etc.). Les phages ont du lysozyme, qui détruit la membrane cellulaire, la phosphatase, etc.
Une autre façon de traiter les infections virales consiste à utiliser des médicaments antiviraux. Ces médicaments ont souvent une capacité limitée à guérir les maladies virales, mais ont été utilisés pour contrôler et réduire les symptômes d'un large éventail de maladies virales. Pour la plupart des virus, ces médicaments inhibent le virus en bloquant les actions d'une ou plusieurs de ses protéines. Il est important que les protéines cibles soient codées par des gènes viraux et que ces molécules ne soient pas présentes dans une cellule hôte saine.
Ainsi, la croissance virale est inhibée sans endommager l'hôte. Il y a beaucoup de médicaments antivirauxdisponibles pour traiter les infections, certaines spécifiques à un virus particulier et d'autres pouvant affecter plusieurs virus. Les médicaments antiviraux sont conçus pour traiter l'herpès génital et la grippe. Pour l'herpès génital, des médicaments tels que l'acyclovir peuvent réduire le nombre et la durée des épisodes de maladie virale active au cours desquels les patients développent des lésions virales dans les cellules de la peau.
Détection des antigènes viraux... Les antigènes viraux dans les cellules hôtes infectées peuvent être détectés en utilisant des techniques d'immunofluorescence. Préparations contenant des cellules virus infectésont traités avec des sérums immuns luminescents spécifiques. Lorsqu'il est observé au microscope fluorescent, une lueur caractéristique est observée dans les endroits où les particules virales s'accumulent. Le type de virus est déterminé par la correspondance du sérum luminescent spécifique qui a provoqué la luminescence.
Parce que le virus reste caché dans les tissus nerveux du corps pour la vie, ce médicament n'est pas un remède, mais il peut rendre les symptômes de la maladie plus gérables. Pour la grippe, des médicaments comme le Tamiflu peuvent réduire la durée des symptômes de «grippe» d'un ou deux jours, mais le médicament ne prévient pas complètement les symptômes. Autres médicaments antivirauxtelles que la ribavirine ont été utilisées pour traiter diverses infections virales.
Lorsque l'un de ces médicaments est utilisé seul, le taux de mutation élevé du virus permet au virus de développer rapidement une résistance au médicament. Les virus sont des entités dépourvues de cellules qui ne peuvent généralement être observées qu'au microscope électronique. Les virus sont divers et infectent les archées, les bactéries, les champignons, les plantes et les animaux. Les virus sont constitués d'un noyau d'acide nucléique entouré d'une capside protéique avec ou sans enveloppe lipidique externe.
L'introduction du virus dans la cellule, son interaction avec la cellule hôte et la reproduction (reproduction) se composent d'une série d'étapes successives.
Étape 1. Commence par le processus d'adsorption aux dépens du virion et des récepteurs cellulaires. Dans les virions complexes, les récepteurs sont situés à la surface de la membrane sous forme d'excroissances styloïdes (virus de la grippe), dans des virions simples, à la surface de la capside.
Étape 2. La pénétration du virus dans la cellule hôte se déroule différemment pour différents virus. Par exemple, certains phages percent la membrane avec leur processus et injectent de l'acide nucléique dans la cellule hôte (voir chapitre 8). D'autres virus pénètrent dans la cellule en aspirant une particule virale à l'aide d'une vacuole, c'est-à-dire qu'une dépression se forme au site d'introduction dans la membrane cellulaire, puis ses bords sont fermés et le virus apparaît dans la cellule. Cette rétraction est appelée viropexis.
Étape 3. «Décapage du virus» (désintégration). Pour sa reproduction, l'acide nucléique viral est libéré des enveloppes protéiques (enveloppe et capside) qui le protègent. Le processus de déshabillage peut commencer pendant l'adsorption, ou il peut se produire lorsque le virus est déjà à l'intérieur de la cellule.
Stade 4. A ce stade, la réplication (reproduction) des acides nucléiques et la synthèse des protéines virales se produisent. Cette étape se produit avec la participation de l'ADN ou de l'ARN de la cellule hôte.
Étape 5. Assemblage du virion. Ce processus est facilité par l'auto-assemblage de particules protéiques autour de l'acide nucléique viral. La synthèse des protéines peut commencer immédiatement après la synthèse des acides nucléiques viraux, ou après un intervalle de plusieurs minutes ou plusieurs heures. Dans certains virus, l'auto-assemblage se produit dans le cytoplasme. D'autres ont des cellules hôtes dans le noyau. La formation de la coque externe (peplos) se produit toujours dans le cytoplasme.
Stade 6. La libération du virion de la cellule hôte se produit par percolation du virus à travers la membrane cellulaire ou à travers un trou formé dans la cellule hôte (dans ce cas, la cellule hôte meurt).
Types d'interaction virus-cellule... Le premier type, l'infection productive, se caractérise par la formation de nouveaux virions dans la cellule hôte.
Le deuxième type - infection abortive - est que la réplication de l'acide nucléique est interrompue.
Le troisième type est caractérisé par l'incorporation d'acide nucléique viral dans l'ADN de la cellule hôte; il existe une forme de coexistence du virus et de la cellule hôte (virogénie). Dans ce cas, la synchronicité de la réplication de l'ADN viral et cellulaire est assurée. Dans les phages, cela s'appelle la lysogénie.
Examen microscopique... Avec des infections virales individuelles dans le cytoplasme ou les noyaux des cellules de l'hôte, des corps intracellulaires spécifiques sont observés - des inclusions qui ont une valeur diagnostique (corps de Babesh-Negri en cas de rage, corps de Guarnieri dans la variole, etc.). Les tailles des particules virales et des corps-inclusions peuvent être artificiellement augmentées par des méthodes spéciales de traitement des préparations avec mordant et imprégnation (par exemple, la méthode d'argenture selon Morozov) et observées au microscope à immersion. Les virions plus petits qui se trouvent en dehors de la portée de vue d'un microscope optique ne sont détectés que par microscopie électronique. Il existe différents points de vue concernant les inclusions intracellulaires. Certains auteurs pensent qu'ils représentent une collection de virus. D'autres pensent qu'ils résultent de la réaction de la cellule à l'introduction de virus.
Génétique des virus... La modification (changements non héréditaires) des virus est due aux caractéristiques de la cellule hôte dans laquelle le virus se reproduit. Les virus modifiés acquièrent la capacité d'infecter des cellules similaires à celles dans lesquelles ils ont été modifiés. Différents virus se manifestent de différentes manières. Par exemple, dans les phages, la forme des «points négatifs» (colonies de phages) change.
Mutation - dans les virus, elle se produit sous l'influence des mêmes mutagènes qui provoquent une mutation chez les bactéries (facteurs physiques et chimiques). Une mutation se produit lors de la réplication de l'acide nucléique. Les mutations affectent diverses propriétés des virus, par exemple, la sensibilité à la température, etc.
La recombinaison génétique dans les virus peut résulter de l'infection simultanée d'une cellule hôte avec deux virus, tandis que des gènes individuels peuvent être échangés entre deux virus et des recombinants sont formés contenant les gènes de deux parents.
La réactivation génétique des gènes se produit parfois lorsqu'un virus inactivé est croisé avec un virus à part entière, ce qui conduit au salut du virus inactivé.
La génétique spontanée et dirigée des virus est d'une grande importance dans le développement du processus infectieux.
Résistant aux facteurs environnementaux... La plupart des virus sont inactivés par l'action hautes températures... Cependant, il existe des exceptions, par exemple le virus de l'hépatite résiste à la chaleur.
À basses températures les virus ne sont pas sensibles, les rayons ultraviolets du soleil ont un effet inactivant sur les virus. La lumière du soleil dispersée agit moins activement sur eux. Les virus sont résistants au glycérol, ce qui permet de les conserver longtemps dans la glycérine. Ils sont résistants aux antibiotiques (lors de la culture de virus, le matériel d'essai est traité avec des antibiotiques pour supprimer la flore bactérienne).
Les acides, les alcalis, les désinfectants inactivent les virus. Cependant, certains virus inactivés au formol conservent leurs propriétés immunogènes, ce qui permet d'utiliser le formol pour obtenir des vaccins (vaccin antirabique).
Susceptibilité des animaux... L'éventail des animaux sensibles à certains virus est très large, par exemple, de nombreux animaux sont sensibles aux virus de la rage. Certains virus n'affectent qu'une seule espèce animale, par exemple le virus de la peste canine n'affecte que les chiens. Il existe des virus auxquels les animaux ne sont pas sensibles - par exemple, le virus de la rougeole, etc.
Organotropie des virus... Les virus ont la capacité d'infecter certains organes, tissus et systèmes. Par exemple, le virus de la rage attaque le système nerveux. Le virus de la variole est dermatrope, etc.
Libération de virus dans l'environnement... À partir d'un corps malade, des virus peuvent être excrétés dans les matières fécales, par exemple le virus de la poliomyélite et d'autres entérovirus. Le virus de la rage est excrété dans la salive, le virus de la grippe est excrété par l'écoulement de la muqueuse nasopharyngée, etc.
Les principales voies de transmission des virus... Gouttelettes en suspension dans l'air (grippe, variole), nourriture (poliomyélite, hépatite A), contact et ménage (rage), transmissibles (encéphalite).
Immunité antivirale... Le corps humain a une résistance innée à certains virus. Par exemple, les humains ne sont pas sensibles au virus de la peste canine. Les animaux ne sont pas sensibles au virus de la rougeole. Dans ces cas, l'immunité antivirale repose sur l'absence de cellules capables de soutenir la reproduction des virus.
L'immunité antivirale est déterminée à la fois par des facteurs de défense cellulaires et humoraux, non spécifiques et spécifiques. Facteurs non spécifiques. Un puissant inhibiteur de la reproduction virale est une substance protéique - l'interféron. À corps sain il est contenu en quantité insignifiante et les virus contribuent à la production d'interféron et sa quantité augmente considérablement. Il n'est pas spécifique car il bloque la reproduction de divers virus. Cependant, il a une spécificité tissulaire, c'est-à-dire que les cellules de différents tissus forment différents interférons. On pense que son mécanisme d'action réside dans le fait qu'il interfère avec la synthèse des protéines dans la cellule hôte et arrête ainsi la reproduction du virus.
Les facteurs spécifiques de l'immunité antivirale comprennent les anticorps neutralisant les virus, l'hémagglutination et la précipitation.
Méthodes de culture de virus... Les virus se reproduisent uniquement dans les cellules viables. Ils sont cultivés: dans des embryons de poulet (fig. 53), des cultures de tissus humains et divers animaux, dans le corps d'animaux sensibles, des arthropodes sensibles.
Dans la première période du développement de la virologie, la principale méthode d'étude des virus était l'infection artificielle des animaux, mais cette méthode est complexe et, en plus de cela, les animaux étaient immunisés contre de nombreux virus.
L'introduction de méthodes pour la culture de virus dans des embryons de poulet et dans la culture de cellules de tissus humains et animaux a été d'une grande importance dans le développement de la virologie.
Infection d'embryons de poulet... Pour la reproduction des virus, on utilise des embryons de poulet âgés de 7 à 12 jours, incubés dans un thermostat à 37 ° C.Une condition préalable au bon développement de l'embryon est le respect d'une certaine humidité de l'air, qui peut être créée en plaçant un récipient contenant de l'eau dans le thermostat.
L'aptitude d'un embryon de poulet à l'infection est déterminée par la présence de mouvements de l'embryon et d'un réseau développé de vaisseaux sanguins sur la membrane chorion-allantoïque lors de la numérisation avec un ovoscope.
La culture de virus dans des embryons de poulet est effectuée à différents endroits de l'embryon, qui est infecté (voir Fig.53):
1) sur la membrane chorion-allantoïque,
2) dans la cavité allantoïdienne;
3) dans la cavité amniotique;
4) dans le sac vitellin.
Les embryons de poulet sont infectés dans une boîte à l'aide d'instruments stériles. Avant l'infection, les embryons de poulet sont essuyés deux fois avec un coton-tige imbibé d'alcool.
Infection sur la membrane chorion-allantoïque. Après la désinfection, les œufs sont soigneusement coupés d'un morceau de coquille de l'extrémité émoussée, la membrane de la coquille est retirée et la membrane chorion-allantoïque est trouvée. Un matériau infectieux en une quantité de 0,1 à 0,2 ml à l'aide d'une seringue ou d'une pipette Pasteur est appliqué sur la membrane chorion-allantoïque. Après l'infection, le trou est fermé avec un capuchon et l'espace entre celui-ci et l'embryon de poulet est rempli de paraffine.
De l'autre côté de l'œuf, écrivez au crayon le nom du matériel infectieux et la date de l'infection.
Infection dans la cavité amniotique. L'œuf est ovoscopé et sur le côté latéral un site est sélectionné où le chorion-allantoïde est dépourvu de gros vaisseaux sanguins. Cette zone est marquée avec un crayon. Les œufs sont placés sur un support en position horizontale, désinfectés et un trou dans la coquille est percé d'une lance stérile spéciale à une profondeur de 213 mm, à travers laquelle une aiguille contenant du matériel infectieux est insérée à la même distance directement dans la cavité amniotique. Afin d'empêcher le liquide injecté de refluer, une perforation est pratiquée au-dessus du coussin gonflable, après quoi les deux trous sont remplis de paraffine.
Infection dans la cavité allantoïdienne. L'infection est réalisée dans une boîte sombre. L'espace d'air est noté, la coquille au-dessus de l'espace d'air est désinfectée et une aiguille de seringue avec le matériau est insérée à travers le trou dans la coquille vers l'embryon. Si l'aiguille pénètre dans la cavité allantoïdienne, l'ombre de l'embryon est déplacée. Après l'infection, le trou est rempli de paraffine.
Infection dans le sac vitellin. La coque est désinfectée. L'œuf est placé sur le support avec l'extrémité émoussée vers la droite afin que le sac vitellin soit tourné vers le haut. Un trou est percé au-dessus de la chambre à air au centre. À travers le trou dans la coquille dans une direction horizontale sur une profondeur de 2-3 mm, une aiguille de seringue est insérée, qui pénètre dans le sac vitellin. Le matériau est injecté dans un volume de 0,2 à 0,3 ml. Après l'introduction du matériau, le trou est ciré.
Le régime de température et la durée d'incubation dépendent des propriétés biologiques du virus introduit.
Les œufs infectés sont contrôlés quotidiennement - ovoscopie pour vérifier la viabilité de l'embryon. Si les embryons meurent le premier jour, la cause en est généralement un traumatisme lors de l'infection. Ces œufs sont issus de l'expérience.
S'il est nécessaire d'examiner séparément chaque composant de l'embryon, le matériel est collecté dans un certain ordre: le liquide allantoïque est aspiré, puis le liquide amniotique, la membrane chorion-allantoïque est coupée, la membrane amniotique, l'embryon, le sac vitellin sont séparés, et alors seulement la membrane chorion-allantoïque est retirée, en la séparant de l'intérieur surface de la coque. La présence du virus dans l'embryon infecté est déterminée par les modifications caractéristiques de la membrane chorion-allantoïque de l'embryon de poulet infecté.
Les virus qui n'ont pas d'activité hémagglutinante sont détectés à l'aide des CSC.
Pour détecter le virus dans les liquides allantoïdiens ou amniotiques des embryons infectés, un RHA est placé (l'hémagglutination est causée par des liquides allantoïdiens ou amniotiques ou une suspension préparée à partir de la membrane chorion-allantoïque).
Culture de virus en culture cellulaire... Pour l'accumulation de virus dans des cultures de cellules sensibles, des tissus humains et divers animaux sont utilisés. La plus grande application pratique a été reçue par des cultures monocouches de lignées cellulaires primaires trypsinisées et transplantées.
Les cultures cellulaires monocouches sont cultivées dans des récipients à matelas plats en verre. La suspension cellulaire dans un milieu nutritif liquide à une température de 37 ° C permet d'obtenir une couche "in vitro" de cellules avec une certaine structure histologique. La présence de virus dans les cultures de tissus est détectée par le changement (dégénérescence) des cellules. Le type de virus est déterminé en neutralisant l'effet des virus en ajoutant des sérums spécifiques de type appropriés au matériel vacciné.
Ces méthodes permettent une intégration plus rapide des résultats de la recherche et sont plus rentables. Dans les cas où les virus ne provoquent pas d'action cytopathique (dégénérescence) et ne se développent pas dans les embryons de poulet, des méthodes d'infection des animaux sont utilisées (voir chapitre 11).
Pour la culture de virus, des cellules continues sont utilisées, qui sont le plus souvent obtenues à partir de cellules de tumeurs malignes.
Les cultures monocouches sont obtenues à partir d'embryons humains, de poulet et d'animaux.
L'avantage des cultures cellulaires monocouche est la simplicité de la technique et la facilité de comptabilité.
La capacité des cellules à se reproduire à l'extérieur du corps est liée au degré de différenciation tissulaire. Les tissus moins différenciés ont une plus grande capacité à proliférer (tissu conjonctif, épithélial).
L'essence des méthodes de préparation des cultures de tissus primaires est la destruction du tissu intercellulaire et la séparation des cellules pour la production ultérieure d'une monocouche.
La dissociation des cellules est réalisée en agissant sur le tissu des enzymes protéolytiques, le plus souvent la trypsine. La solution de trypsine favorise la séparation des cellules tout en conservant leur capacité à se reproduire. La croissance des cultures cellulaires nécessite un milieu nutritif. La composition du milieu est complexe, il comprend un certain nombre d'ingrédients: acides aminés, glucose, vitamines, sels minéraux, coenzymes, etc. La culture tissulaire est obtenue dans des conditions strictement aseptiques. Des antibiotiques (500 U de pénicilline et 250 U de streptomycine dans 1 ml) sont ajoutés au milieu pour supprimer la croissance de la flore bactérienne.
Le tissu préparé est versé avec une solution à 0,25% de trypsine chauffée et incubé dans un thermostat à 37 ° C. Pendant l'incubation, le tissu est périodiquement agité en faisant tourner le ballon. Les cellules trypsinisées sont centrifugées à 800-1000 tr / min pendant 5 minutes.
La trypsinisation et la centrifugation sont effectuées très soigneusement pour ne pas blesser les cellules. Après centrifugation, le surnageant est éliminé et le sédiment cellulaire est placé dans un petit volume de milieu de culture. Pour obtenir une masse homogène, la suspension de cellules est filtrée sur une couche de gaze dans un entonnoir (stérile). La stérilité de la suspension cellulaire est vérifiée en inoculant 0,1 ml dans 2 tubes avec du bouillon de sucre.
Le succès de la culture cellulaire dépend de la dose d'inoculation, par conséquent, après la trypsinisation, les cellules sont comptées dans la chambre de Goryaev. Après comptage, la suspension de cellules est diluée avec un milieu nutritif pour que 1 ml contienne 500 000-1000000 cellules et est versée dans des éprouvettes et des matelas. Les tubes de culture tissulaire sont incubés dans un incubateur en position inclinée.
Les cultures inoculées sont examinées quotidiennement sous un microscope à faible grossissement pour déterminer leurs modèles de croissance. Les cellules proliférantes normales sont de couleur claire et se développent en une seule couche. Si les cellules sont sombres, granuleuses et ne prolifèrent pas, ce qui peut être le résultat d'une contamination (mauvaise manipulation des plats ou contamination des ingrédients), alors ces cultures sont retirées de l'expérience.
Changer le milieu nutritif 2-3 jours après le semis améliore l'intensité de la prolifération.
Des cellules normales, bien proliférantes, sont infectées avec le matériel d'essai.
Les cultures transplantées sont principalement obtenues à partir de tumeurs malignes. Souche Hela - culture de cellules cancéreuses cervicales d'une femme nommée Helena (obtenue en 1950); la souche Hep-2 a été isolée d'un patient atteint d'un cancer du larynx. La croissance de ces cellules est maintenue dans les laboratoires par passages successifs. Leur particularité réside dans le fait qu'ils se reproduisent longtemps. À l'heure actuelle, ces cellules ont traversé des milliers de générations. Au cours du passage, ils perdent certaines propriétés morphologiques et biochimiques - ils subissent des mutations. Cependant, ils restent tout à fait adaptés à la culture de virus qu'ils contiennent. La culture de ces cellules est utilisée par des laboratoires du monde entier.
La reproduction du virus en culture cellulaire se produit à des moments différents, en fonction des propriétés du virus et du type de cellules.
La présence du virus est jugée par l'effet cytopathique. La dégénérescence cellulaire est observée au microscope. Le temps d'action cytopathique et sa nature dépendent de la dose et des propriétés du virus.
Dans certains virus, un effet cytopathique est détecté après quelques jours (virus de la variole), dans d'autres, après 1 à 2 semaines (virus de l'hépatite, etc.).
Actuellement, il existe déjà des centaines de virus connus pour infecter les humains. La lutte contre les infections virales est menée par différentes méthodes. La vaccination est la plus efficace. De cette manière, la variole a été éliminée et l'incidence de la poliomyélite a été réduite. La prévention sociale est importante dans la lutte contre les infections virales - la destruction des chiens errants (lutte contre la rage), la prévention personnelle, etc.
Cependant, ces mesures ne peuvent garantir l'élimination de toutes les maladies virales. Les scientifiques recherchent constamment des moyens par lesquels il serait possible d'infecter le virus sans endommager la cellule dans laquelle il se trouve.
Par conséquent, il est naturel que dans le programme du Parti communiste de l'Union soviétique, la virologie soit nommée l'une des principales branches des sciences naturelles, qui devrait recevoir un développement prioritaire dans les années à venir.
Méthodes de base de la recherche des virus... 1. Réaction d'hémagglutination, réaction d'hémagglutination retardée, réaction d'hémagglutination indirecte. Réaction de liaison du complément.
2. La réaction de neutralisation des virus en culture tissulaire.
3. Méthode d'immunofluorescence.
4. Méthode histologique - identification des inclusions (petits corps de Babesh - Negri - avec rage; petits corps de Pashen - avec variole, etc.).
5. Méthode biologique.
Infection virale une cellule est un complexe de processus résultant de l'interaction d'une cellule avec un génome viral. Le virion extracellulaire est inerte et cette inertie persiste jusqu'à ce que le génome viral commence à fonctionner de manière intracellulaire, en tant qu'unité génétique indépendante. La caractéristique cardinale qui sous-tend la classification donnée ci-dessous est la relation entre les génomes du virus et la cellule. Si le génome viral se réplique indépendamment de la réplication du génome cellulaire, il s'agit d'un type autonome d'infection virale. Si le génome viral est inclus dans le génome cellulaire et se réplique avec lui, il s'agit d'un type intégratif. Chacun de ces deux principaux types d'infection virale peut être subdivisé en plusieurs formes en fonction de la formation du virus (infection productive et abortive), selon l'issue du processus infectieux de la cellule (infection lytique et non lytique), selon la dynamique d'interaction entre le virus et la cellule (infection aiguë et chronique) ).
Un type autonome d'infection virale d'une cellule est caractéristique de la plupart des virus connus des animaux et des humains. Dans ce type d'infection, le génome viral se réplique indépendamment de la réplication du génome cellulaire.
Une infection virale autonome productive est caractérisée par la formation d'un virus complet. C'est la forme d'interaction la plus étudiée entre virus et cellules. Les mécanismes de base de la reproduction virale discutés au chapitre 3 ont été étudiés et décrits pour cette forme particulière d'infection.
Une infection virale avortée autonome se produit si l'interaction du virus avec la cellule n'entraîne pas la formation de virions pleinement précieux. Cela se produit généralement lorsque les cellules sont infectées par un virus défectueux; lorsque le virus pénètre dans les cellules qui lui résistent; lorsque les cellules sensibles sont infectées par un virus de grande valeur dans des conditions non permissives.
L'infection virale autonome lytique entraîne la mort cellulaire. Le virus qui cause la mort cellulaire est appelé cytopathogène, et les dommages cellulaires sont désignés par le terme «cytope, effet togénique». La nature des changements cytopathologiques dans les infections virales est très diverse. Cela pourrait être degu
Défaillance cellulaire (vacuolisation du cytoplasme, pycnose nucléaire, aberrations chromosomiques), formation d'inclusions intracellulaires virales spécifiques, émergence de cellules géantes telles que polycaryocytes ou symplastes. La lyse cellulaire est la dernière étape de l'action des virus cytopathogènes. Pour la manifestation de la cytopathogénicité, le virus ne nécessite pas la reproduction du génome et la mise en œuvre de toutes les informations génétiques.
Une infection virale autonome non lytique est caractéristique des virus contenant une membrane externe de lipoprotéine et des cellules bourgeonnantes pour émettre des cellules. Les arénavirus rétro, orthomyxo, paramyxo sont capables de provoquer une infection cellulaire non lytique productive. Cependant, dans les cas les plus typiques, un caractère non lytique est caractéristique des infections abortives, lorsque les propriétés génétiques des virus nécessaires à la mise en œuvre des fonctions cytopathogènes sont absentes ou n'apparaissent pas.
Selon la dynamique du processus infectieux, les infections virales sont divisées en aiguës et chroniques. Quand infection aiguë dans la cellule infectée, la formation de virions se produit, après quoi le processus infectieux s'arrête. Dans une infection chronique, la cellule infectée produit des virions ou des composants viraux jusqu'à sa mort naturelle (et non à partir d'une infection virale donnée), tandis que les cellules filles restent infectées. Une infection lytique productive et avortée est toujours aiguë. Une infection non lytique productive est toujours chronique. En règle générale, l'infection non lytique abortive a une évolution chronique, mais elle peut également se produire sous une forme aiguë.
Le type intégratif d'infection virale est caractérisé par l'union physique (intégration) des génomes viral et cellulaire. Dans ce cas, le génome viral se réplique et fonctionne comme une partie intégrante du génome cellulaire. Ce type d'infection virale est obligatoire pour les rétrovirus et est possible dans le cas des papovirus, adénovirus et herpès virus.
Les infections intégratives peuvent être classées selon les mêmes paramètres que les infections autonomes - productivité, dynamique et résultat; la relation d'intégration entre le virus et la cellule peut également être productive et avortée. L'infection intégrative productive est typique des rétrovirus. L'avortement, une infection rétrovirale intégrative, se produit dans les mêmes conditions que celles décrites ci-dessus pour les infections avortées autonomes. Ainsi, les rétrovirus défectueux sont dépourvus de
De nouvelles parties du génome (certaines souches du virus du sarcome de Rous, les virus du sarcome de souris, de chats, de singes, de leucémies de souris Frend et Rauscher, etc.) peuvent à elles seules provoquer une infection intégrative abortive, qui peut se transformer en infection productive en présence d'un virus assistant. L'infection est un exemple d'infection intégrative abortive causée par un rétrovirus pleinement valable dans des cellules non permissives. virus aviaire Sarcome de Rous de cellules de mammifères. Dans le même temps, un processus d'intégration avorté se produit dans les cellules, qui se caractérise par une production réduite d'ARN viral, une accumulation d'un assortiment incomplet de protéines virales, une altération de leur traitement et une absence de formation de virions.
Habituellement, les infections intégratives sont non lytiques, comme dans le cas des rétrovirus de types B, C, D, papov oncogènes, adéno, herpès virus, mais il existe également des infections lytiques causées par le rétrovirus cytopathogène de la réticuloendothéliose aviaire, ou le virus «moussant».
L'infection intégrative virogène est une infection avortée qui peut être traduite en une forme productive. Les cellules atteintes de cette forme d'infection peuvent spontanément ou sous l'influence d'influences spéciales (physiques, chimiques, biologiques) commencer la production d'un virus infectieux. Ce phénomène est appelé virogénie, et les cellules sont désignées comme virogènes, par analogie avec la lysogénie et les cellules lysogéniques dans le système phage-bactérien.
En savoir plus sur le sujet Infection des cellules virales:
- Infections virales et infections d'étiologie virale suspectée
- Infections bactériennes et virales aéroportées: grippe, parainfluenza, infection à adénovirus, infection respiratoire syncytiale. Bronchopneumonie bactérienne, pneumonie lobaire.