Bolile virale au apărut în cele mai vechi timpuri, dar virologia ca știință a început să se dezvolte la sfârșitul secolului al XIX-lea.
În 1892, omul de știință-botanist rus DI Ivanovsky, studiind boala mozaicului din frunzele de tutun, a constatat că această boală este cauzată de cele mai mici microorganisme care trec prin filtrele bacteriene cu pori fini. Aceste microorganisme sunt numite virusuri filtrabile (din virusul latin - otravă). Mai târziu s-a arătat că există și alte microorganisme care trec prin filtrele bacteriene, prin urmare, virusurile filtrate au început să fie numite pur și simplu virusuri.
În general, forma virionului și prezența sau absența unui plic spun puțin despre ce boli pot provoca virușii sau ce specii pot infecta, dar sunt totuși o modalitate utilă de a începe clasificarea virală. Figura 5 Virușii pot avea o formă complexă sau relativ simplă.
Care dintre următoarele afirmații despre structura virusului este corectă? Nucleul virusului conține genomul sau conținutul genetic general al virusului. Genomii virali sunt de obicei mici în comparație cu bacteriile sau eucariotele, care conțin doar gene care codifică proteinele pe care virusul nu le poate obține de la celula gazdă. Acest material genetic poate fi monocatenar sau bicatenar. Poate fi, de asemenea, liniar sau circular. În timp ce majoritatea virușilor conțin un segment de acid nucleic, alții au genomi care sunt alcătuite din mai multe segmente.
O mare contribuție la studiul virusurilor a fost făcută de virologii sovietici: MA Morozov, N. F. Gamaleya, L. A. Zilber, M. P. Chumakov, A. A. Smorodintsev, V. M. Zhdanov și alții.
Virușii sunt formă necelulară existența materiei vii. Sunt foarte mici. Conform expresiei figurative a lui VM Zhdanov, „dimensiunea lor în raport cu dimensiunea bacteriilor medii poate fi comparată cu dimensiunea unui șoarece în raport cu un elefant”. A devenit posibil să se vadă viruși numai după inventarea microscopului electronic.
Virușii pot fi considerați obligați paraziți intracelulari... Virusul trebuie să se atașeze de o celulă vie, să o ingereze, să-și producă proteinele și să-i copieze genomul și să găsească o modalitate de a scăpa de celulă, astfel încât virusul să poată infecta alte celule și, în final, alte persoane. Virușii pot infecta numai anumite tipuri de gazde și numai anumite celule din acea gazdă. Baza moleculară pentru această specificitate este că o moleculă specifică de suprafață cunoscută sub numele de receptor viral trebuie găsită pe suprafața celulei gazdă pentru ca virusul să se atașeze.
În prezent, multe metode sunt folosite pentru a studia virusurile: chimice, fizice, moleculare biologice, imunobiologice și genetice.
Toate virusurile sunt împărțite în oameni care infectează, animale, insecte, bacterii și plante.
Virușii au o mare varietate de forme și proprietăți biologice, dar toate au trăsături structurale comune. Particulele de virus mature se numesc virioni.
În plus, diferențele metabolice observate în diferite tipuri de celule pe baza expresiei genetice diferențiale sunt un factor probabil în care celulele pot utiliza virusul pentru replicare. Celula trebuie să alcătuiască substanțele necesare virusului, cum ar fi enzimele, în care genomul viral în sine nu conține gene sau virusul nu se poate reproduce folosind această celulă.
Virusul trebuie să „preia” celula pentru replicare. Ciclul de replicare virală poate duce la modificări biochimice și structurale dramatice în celula gazdă care pot provoca leziuni celulare. Aceste modificări, numite efecte citopatice, pot modifica funcția celulei sau chiar distruge celula. Unele celule infectate, cum ar fi cele infectate cu virusul răcii obișnuite, mor prin liză sau apoptoză, eliberând simultan toți virionii descendenților. Simptome boli virale sunt rezultatul unui răspuns imun la un virus care încearcă să controleze și să elimine virusul din organism, precum și din deteriorarea celulelor cauzate de virus.
Spre deosebire de alte microorganisme care conțin atât ADN, cât și ARN, virionul conține doar unul dintre acizii nucleici - fie ADN, fie ARN.
Acidul nucleic al virușilor poate fi monocatenar și bicatenar. Aproape toți virusii care conțin ARN au ARN monocatenar în genomul lor, iar cei care conțin ADN au ADN bicatenar. În conformitate cu două tipuri de substanță genetică, virușii sunt împărțiți în ARN și ADN. Familiile care conțin ADN includ 5 familii, cele care conțin ARN - 10 familii.
În timpul procesului de înmugurire, celula nu este lizată sau ucisă imediat. Majoritatea infecțiilor virale productive urmează pași similari în ciclul de replicare a virusului: cuibărire, infiltrare, dezvelire, replicare, asamblare și eliberare. Virusul se atașează la un site specific al receptorului de pe membrana celulei gazdă prin intermediul proteinelor de atașament din capsidă sau proteinelor încorporate în anvelopa sa. Atașamentul este specific și, de obicei, virusul se va atașa numai la celulele uneia sau mai multor specii și numai la anumite tipuri de celule din acele specii cu receptorii corespunzători.
* (Iată date despre unele dintre virusurile patogene pentru oameni.)
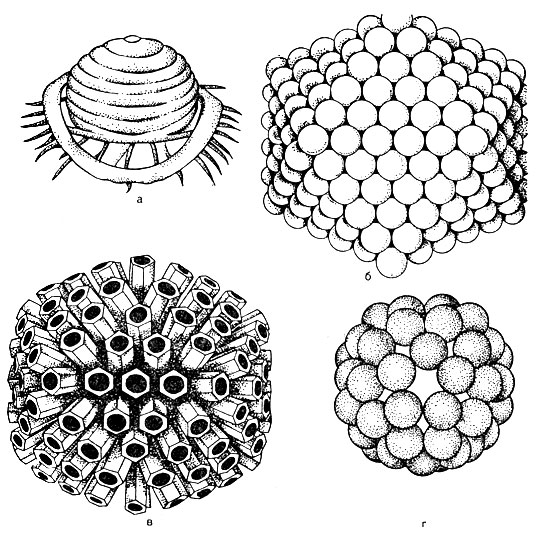
Structura virionului... În centrul virionului există un acid nucleic, care este înconjurat de o capsidă (din greaca kanca - cutie). Capsidul este alcătuit din subunități proteice numite capsomere. Un virus matur este o nucleocapsidă în structura chimică. Numărul capsomerilor și modul în care sunt ambalate (Fig. 52) sunt strict constante pentru fiecare tip de virus. De exemplu, virusul poliomielitei conține 32 capsomere, în timp ce adenovirusul conține 252 capsomere. Capsomerii pot fi stivuiti sub forma unui poliedru cu margini simetrice uniforme - forma cuboidala (de exemplu, adenovirus). Stilul sferic este tipic pentru virusurile gripale. Poate exista un tip de simetrie în care acidul nucleic are forma unui izvor în jurul căruia sunt așezate capsomere, caz în care virusul are o formă în formă de tijă - un virus care provoacă boala frunzelor de tutun.
Spre deosebire de virusurile animale, acidul nucleic bacteriofagic este introdus în celula gazdă gol, lăsând capsida în afara celulei. Virusurile vegetale și animale pot pătrunde în celule prin endocitoză, în care membrana celulară înconjoară și înghite întregul virus. Unii viruși înveliți intră în celulă atunci când învelișul viral fuzionează direct cu membrana celulei. Capsidul viral se descompune apoi în interiorul celulei și acidul nucleic viral este eliberat, care apoi devine disponibil pentru replicare și transcriere.
Mecanismul de replicare depinde de genomul viral. Există, desigur, excepții de la acest model. Dacă celula gazdă nu furnizează enzimele necesare pentru replicarea virală, genele virale oferă informații pentru sinteza directă a proteinelor lipsă.
Fagul are un tip complex de simetrie: capul este cuboidal, iar procesul este în formă de tijă (în formă de spermă) (vezi Fig. 21, 22).
Astfel, în funcție de metoda de ambalare, virușii sunt împărțiți în forme cuboidale, sferice, în formă de tijă și spermatozoizi.
Ultimul pas în replicarea virală este eliberarea de noi virioni în gazdă, unde aceștia sunt capabili să infecteze celulele vecine și să repete ciclul de replicare. Unii viruși sunt eliberați atunci când celula gazdă moare, în timp ce alți viruși pot părăsi celulele infectate prin scurgerea prin membrană fără a ucide celula direct.
Figura 6 Când sunt infectate cu virusul gripal, glicoproteinele se atașează la celula epitelială gazdă. Ca urmare, virusul este absorbit. Figura 7 Virușii cauzează o varietate de boli la om, variind de la boli minore la boli grave.
Unele viruși, care sunt mai complexe, au o coajă numită peplos. Se formează atunci când virusul părăsește celula gazdă. În acest caz, capsida virală este învelită de suprafața interioară a membranei citoplasmatice a celulei gazdă și se formează unul sau mai multe straturi ale anvelopei supercapsidelor. Doar unii viruși au un astfel de plic, de exemplu, virusurile rabiei, herpesului, encefalitei. Această coajă conține fosfolipide, care sunt degradate de eter. Astfel, acționând asupra eterului, este posibil să se distingă un virus cu peplos de un virus cu „capsidă goală”.
Vaccinul poate fi preparat folosind viruși vii atenuați, viruși uciși sau subunități moleculare ale virusului. În general, virusurile vii duc la creșterea imunității, dar au capacitatea de a provoca boli la o frecvență scăzută. Vaccinul viral ucis și virusul subunității nu sunt capabili să provoace boli, dar în general duc la o imunitate mai puțin eficientă sau pe termen lung.
Vaccinurile cu virus viu atenuat sunt concepute în laborator pentru a provoca puține simptome la destinatari, oferindu-le în același timp imunitate împotriva viitoarelor infecții. Poliomielita a fost una dintre bolile care au marcat o etapă importantă în utilizarea vaccinurilor. Succesul vaccinului împotriva poliomielitei a deschis calea pentru distribuția de rutină a vaccinurilor din copilărie împotriva rujeolei, oreionului, rubeolei, varicelei și a altor boli. Vaccinurile vii sunt de obicei produse prin slăbirea virusului de tip sălbatic prin creșterea acestuia în laborator în țesuturi sau la temperaturi diferite de cele cu care este obișnuit virusul în gazdă.
La unii viruși, capsomerii sub formă de spini (acești spini sunt contondenți) ies din stratul lipidic exterior al plicului. Acești viruși sunt numiți peplomeri (de exemplu, virusul gripal, a se vedea Figura 52).
Acidul nucleic al virusului este purtător de proprietăți ereditare, iar capsida și învelișul exterior au funcții de protecție, ca și cum ar proteja acidul nucleic. În plus, acestea facilitează intrarea virusului în celulă.
De exemplu, virusul poate fi crescut în celule in vitro, embrioni de la păsări sau animale vii. Adaptarea la aceste noi celule sau temperatură determină mutații în genomul virusului, permițându-le să crească mai bine în laborator, în timp ce interferează cu capacitatea lor de a provoca boli atunci când sunt reintroduse într-o afecțiune găsită în gazdă. Astfel, virușii atenuați provoacă în continuare infecții, dar nu cresc foarte bine, permițând răspunsurilor imune să evolueze în timp pentru a preveni bolile grave.
Pericolul utilizării vaccinurilor vii, care sunt în general mai eficiente decât vaccinurile ucise, este un risc scăzut, dar semnificativ, ca acești viruși să revină la forma lor cauzatoare de boli prin mutații inverse. Mutațiile inverse apar atunci când un vaccin mutați într-o gazdă, astfel încât să citească gazda și poate provoca din nou boala, care poate fi apoi răspândită la alte persoane într-o epidemie.
Dimensiunile virușilor... Virușii sunt măsurați în nanometri. Mărimea lor fluctuează într-o gamă largă de la 15-20 la 350-400 nm.
Metode de măsurare a virusului: 1) filtrare prin filtre bacteriene cu o dimensiune a porilor cunoscută; 2) ultracentrifugare - virusurile mari precipită mai repede; 3) fotografierea virușilor la microscopul electronic.
În gripă, o mutație a genelor pentru moleculele de suprafață ajută virusul să scape de imunitatea de protecție pe care ar fi putut-o primi în sezonul gripal anterior, făcând imperativ ca oamenii să fie vaccinați în fiecare an. Alte virusuri, cum ar fi cele care cauzează boli în copilărie, rujeola, oreion și rubeolă, mută atât de puțin încât același vaccin este utilizat an de an.
Vaccinuri și medicamente antivirale pentru tratament
În unele cazuri, vaccinurile pot fi utilizate pentru a trata o infecție virală activă. În cazul rabiei, o boală neurologică fatală transmisă în saliva virușilor infectați cu rabia, progresia bolii de la momentul mușcăturii animalului până la momentul în care intră în sistemul nervos central poate dura două săptămâni sau mai mult. Acesta este timpul suficient pentru a vaccina o persoană suspectată că a fost mușcată de un animal rabid, iar răspunsul imun crescut din vaccinare este suficient pentru a preveni pătrunderea virusului în țesutul nervos.
Compoziția chimică a virușilor... Cantitatea și conținutul virusurilor ADN și ARN nu sunt aceleași. În ADN, greutatea moleculară variază de la 1 · 10 6 la 1,6 · 10 8, iar în ARN - de la 2 · 10 6 la 9,0 · 10 6.
Proteinele din virioni se găsesc într-un număr nesemnificativ, acestea constând din 16-20 aminoacizi. În plus față de proteinele capsidei, există și proteine \u200b\u200binterne asociate cu acidul nucleic. Proteinele determină proprietățile antigenice ale virușilor și, de asemenea, datorită împachetării dense a lanțurilor polipeptidice, protejează virusul de acțiunea enzimelor celulei gazdă.
În acest fel, se previn consecințele neurologice fatale ale bolii și individul trebuie doar să se recupereze după mușcătura infectată. Această abordare este utilizată și pentru tratarea Ebola, unul dintre cei mai rapizi și mai letali viruși care infectează oamenii, deși infectează de obicei populații limitate. Ebola este, de asemenea, principala cauză de deces la gorile. Prin utilizarea vaccinurilor nou dezvoltate care stimulează răspunsul imun, se speră că sistemele imune ale persoanelor afectate vor controla mai bine virusul, reducând potențial mortalitatea.
Lipidele și carbohidrații se găsesc în învelișul exterior al virionilor complexi. Membrana celulei gazdă este sursa de lipide și carbohidrați. Polizaharidele, care fac parte din unele virusuri, determină capacitatea lor de a provoca aglutinarea eritrocitelor.
Enzime virale... Virușii nu au propriul lor metabolism, deci nu au nevoie de enzime metabolice. Cu toate acestea, s-a constatat că unii viruși au enzime care facilitează penetrarea lor în celula gazdă. De exemplu, în virusul gripal A, s-a găsit neuraminidaza, care scinde acidul neuraminic conținut în membranele celulelor animale (eritrocite etc.). Fagii au lizozimă, care distruge membrana celulară, fosfataza etc.
O altă modalitate de tratare a infecțiilor virale este utilizarea medicamentelor antivirale. Aceste medicamente au adesea capacitate limitată de a vindeca bolile virale, dar au fost utilizate pentru a controla și reduce simptomele pentru o gamă largă de boli virale. Pentru majoritatea virușilor, aceste medicamente inhibă virusul blocând acțiunile uneia sau mai multora dintre proteinele sale. Este important ca proteinele țintă să fie codificate de gene virale și ca aceste molecule să nu fie prezente într-o celulă gazdă sănătoasă.
Astfel, creșterea virală este inhibată fără a afecta gazda. Sunt multi medicamente antiviraledisponibile pentru tratarea infecțiilor, unele specifice unui anumit virus și altele care pot afecta mai mulți viruși. Medicamentele antivirale sunt concepute pentru a trata herpesul genital și gripa. Pentru herpesul genital, medicamente precum aciclovirul pot reduce numărul și durata episoadelor de boli virale active în care pacienții dezvoltă leziuni virale în celulele pielii.
Detectarea antigenelor virale... Antigenele virale din celulele gazdă infectate pot fi detectate folosind tehnici de imunofluorescență. Preparate care conțin celule virus infectatsunt tratate cu seruri luminoase imune specifice. Când este privit la microscop fluorescent, se observă o strălucire caracteristică în locurile în care se acumulează particule virale. Tipul de virus este determinat de corespondența serului luminescent specific care a provocat luminescența.
Deoarece virusul rămâne ascuns în țesutul nervos al corpului pe viață, acest medicament nu este un remediu, dar poate face simptomele bolii mai ușor de gestionat. Pentru gripă, medicamente precum Tamiflu pot reduce durata simptomelor gripei cu una sau două zile, dar medicamentul nu previne complet simptomele. Alții medicamente antiviralecum ar fi ribavirina au fost utilizate pentru a trata o varietate de infecții virale.
Când oricare dintre aceste medicamente este utilizat singur, rata mare de mutație a virusului permite virusului să dezvolte rapid rezistență la medicament. Virușii sunt entități fără celule care, de obicei, pot fi văzute doar cu microscopul electronic. Virușii sunt diverse, infectând arhee, bacterii, ciuperci, plante și animale. Virușii constau dintr-un miez de acid nucleic înconjurat de o capsidă proteică cu sau fără un înveliș lipidic exterior.
Introducerea virusului în celulă, interacțiunea sa cu celula gazdă și reproducere (reproducerea) sunt compuse dintr-o serie de etape succesive.
Etapa 1. Începe cu procesul de adsorbție în detrimentul receptorilor virionali și celulari. La virioni complexi, receptorii sunt localizați pe suprafața plicului sub formă de excrescențe stiloide (virusul gripal), în virioni simpli, pe suprafața capsidei.
Etapa 2. Penetrarea virusului în celula gazdă are loc diferit pentru diferiți viruși. De exemplu, unii fagi străpung membrana cu procesul lor și injectează acid nucleic în celula gazdă (vezi Capitolul 8). Alți viruși intră în celulă atrăgând o particulă virală cu ajutorul unui vacuol, adică se formează o depresiune la locul de introducere în membrana celulară, apoi marginile sale se închid și virusul intră în celulă. Această retragere se numește viropexis.
Etapa 3. „Dezactivarea virusului” (dezintegrare). Pentru reproducerea sa, acidul nucleic viral este eliberat de învelișurile proteice (plic și capsidă) care îl protejează. Procesul de dezbrăcare poate începe în timpul adsorbției sau poate apărea atunci când virusul este deja în interiorul celulei.
Etapa 4. În acest stadiu apare replicarea (reproducerea) acizilor nucleici și sinteza proteinelor virale. Această etapă are loc cu participarea ADN-ului sau ARN-ului celulei gazdă.
Etapa 5. Asamblarea virionului. Acest proces este facilitat de auto-asamblarea particulelor de proteine \u200b\u200bîn jurul acidului nucleic viral. Sinteza proteinelor poate începe imediat după sinteza acidului nucleic viral sau după un interval de câteva minute sau câteva ore. La unele virusuri, auto-asamblarea are loc în citoplasmă. Alții au celule gazdă în nucleu. Formarea învelișului exterior (peplos) are loc întotdeauna în citoplasmă.
Etapa 6. Eliberarea virionului din celula gazdă are loc prin percolarea virusului prin membrana celulară sau printr-o gaură formată în celula gazdă (în acest caz, celula gazdă moare).
Tipuri de interacțiune virus-celulă... Primul tip, infecția productivă, se caracterizează prin formarea de noi virioni în celula gazdă.
Al doilea tip - infecția avortată - este că replicarea acidului nucleic este întreruptă.
Al treilea tip se caracterizează prin încorporarea acidului nucleic viral în ADN-ul celulei gazdă; există o formă de coexistență a virusului și a celulei gazdă (virogenie). În acest caz, se asigură sincronicitatea replicării ADN-ului viral și celular. La fagi, aceasta se numește lizogenie.
Examinare microscopica... Cu infecții virale individuale, corpurile intracelulare specifice sunt observate în citoplasma sau nucleele celulelor gazdei - incluziuni care au valoare diagnostic (corpuri Babesh-Negri în caz de rabie, corpuri Guarnieri în variolă etc.). Dimensiunile particulelor virale și ale corpurilor-incluziuni pot fi crescute artificial prin metode speciale de prelucrare a preparatelor cu mordant și impregnare (de exemplu, metoda de argintare conform lui Morozov) și observate cu microscopie prin imersie. Virioni mai mici care se află în afara vederii unui microscop optic sunt detectați numai cu microscopie electronică. Există diferite puncte de vedere cu privire la incluziunile intracelulare. Unii autori cred că reprezintă o colecție de viruși. Alții cred că apar ca urmare a reacției celulei la introducerea virușilor.
Genetica virusului... Modificarea (modificări ne-moștenite) a virușilor se datorează caracteristicilor celulei gazdă în care se reproduce virusul. Virușii modificați dobândesc capacitatea de a infecta celule similare cu cele în care au fost modificați. Diferitele viruși se manifestă în moduri diferite. De exemplu, forma „petelor negative” (colonii de fagi) se modifică în fagi.
Mutația - la viruși, apare sub influența acelorași mutageni care provoacă mutația bacteriilor (factori fizici și chimici). O mutație apare în timpul replicării acidului nucleic. Mutațiile afectează diferite proprietăți ale virușilor, de exemplu, sensibilitatea la temperatură etc.
Recombinarea genetică în viruși poate apărea ca urmare a infecției simultane a unei celule gazdă cu doi viruși, în timp ce genele individuale pot fi schimbate între doi viruși și se formează recombinați care conțin genele a doi părinți.
Reactivarea genetică a genelor apare uneori atunci când un virus inactivat este încrucișat cu unul complet, ceea ce duce la salvarea virusului inactivat.
Genetica spontană și direcționată a virușilor are o mare importanță în dezvoltarea procesului infecțios.
Rezistent la factorii de mediu... Majoritatea virușilor sunt inactivați prin acțiune temperaturi mari... Cu toate acestea, există excepții, de exemplu, virusul hepatitei este rezistent la căldură.
LA temperaturi scăzute virușii nu sunt sensibili, razele ultraviolete ale soarelui au un efect inactivant asupra virușilor. Lumina soarelui împrăștiată acționează asupra lor mai puțin activ. Virușii sunt rezistenți la glicerol, ceea ce face posibilă menținerea lor în glicerină pentru o lungă perioadă de timp. Sunt rezistente la antibiotice (în timpul cultivării virusurilor, materialul testat este tratat cu antibiotice pentru a suprima flora bacteriană).
Acizii, alcalii, dezinfectanții inactivează virusurile. Cu toate acestea, unele virusuri inactivate cu formalină își păstrează proprietățile imunogene, ceea ce face posibilă utilizarea formalinei pentru a produce vaccinuri (vaccin antirabic).
Susceptibilitatea animalelor... Gama de animale sensibile pentru unele virusuri este foarte largă, de exemplu, multe animale sunt sensibile la virusurile rabiei. Unele virusuri afectează doar o specie de animal, de exemplu virusul ciumei canine afectează numai câinii. Există viruși la care animalele nu sunt sensibile - de exemplu, virusul rujeolei etc.
Organotropia virușilor... Virușii au capacitatea de a infecta anumite organe, țesuturi și sisteme. De exemplu, virusul rabiei atacă sistemul nervos. Virusul variolei este dermatropic etc.
Eliberarea de viruși în mediu... Dintr-un corp bolnav, virușii pot fi excretați în fecale, de exemplu, virusul poliomielitei și alte enterovirusuri. Virusul rabiei este excretat în salivă, virusul gripal este eliminat din descărcarea mucoasei nazofaringiene etc.
Principalele căi de transmitere a virușilor... Picături în aer (gripă, variolă), alimente (poliomielită, hepatită A), contact de uz casnic (rabie), transmisibil (encefalită).
Imunitatea antivirală... Corpul uman are o rezistență înnăscută la anumiți viruși. De exemplu, oamenii nu sunt sensibili la virusul ciumei canine. Animalele nu sunt sensibile la virusul rujeolic. În aceste cazuri, imunitatea antivirală se bazează pe absența celulelor capabile să susțină reproducerea virușilor.
Imunitatea antivirală este determinată atât de factorii de apărare celulari, cât și de cei umorali, nespecifici și specifici. Factori nespecifici. Un inhibitor puternic al reproducerii virale este o substanță proteică - interferon. ÎN corp sanatos este conținut într-o cantitate nesemnificativă, iar virusurile contribuie la producerea de interferon și cantitatea sa crește semnificativ. Este nespecific, deoarece blochează reproducerea diferiților viruși. Cu toate acestea, are specificitate tisulară, adică celulele din diferite țesuturi formează interferon diferit. Se crede că mecanismul său de acțiune constă în faptul că interferează cu sinteza proteinelor din celula gazdă și, prin urmare, oprește reproducerea virusului.
Factorii specifici ai imunității antivirale includ anticorpi neutralizanți împotriva virușilor, anticorpi hemaglutinați și precipitați.
Metode de cultivare a virusului... Virușii se reproduc numai în celule viabile. Sunt cultivate: la embrioni de pui (Fig. 53), culturi de țesuturi ale oamenilor și ale diferitelor animale, în corpul animalelor sensibile, artropode sensibile.
În prima perioadă a dezvoltării virologiei, principala metodă pentru studierea virușilor a fost infecția artificială a animalelor, dar această metodă este complexă și, pe lângă aceasta, animalele erau imune la mulți viruși.
De o mare importanță în dezvoltarea virologiei a fost introducerea metodelor de cultivare a virușilor în embrioni de pui și în cultura celulelor țesuturilor umane și animale.
Infecția embrionului de pui... Pentru reproducerea virușilor se utilizează embrioni de pui cu vârsta cuprinsă între 7-12 zile, incubați într-un termostat la 37 ° C. O condiție prealabilă pentru dezvoltarea corectă a embrionului este respectarea unei anumite umidități a aerului, care poate fi creată prin plasarea unui vas cu apă în termostat.
Adecvarea unui embrion de pui pentru infecție este determinată de prezența mișcărilor embrionului și a unei rețele dezvoltate de vase de sânge pe membrana corion-alantoică atunci când se scanează cu un ovoscop.
Cultivarea virușilor în embrionii de pui se efectuează în diferite locuri ale embrionului, care este infectat (vezi Fig. 53):
1) pe membrana corion-alantoică,
2) în cavitatea alantoică;
3) în cavitatea amniotică;
4) în sacul gălbenușului.
Embrionii de pui sunt infectați într-o cutie folosind instrumente sterile. Înainte de infecție, embrionii de pui sunt șterși de două ori cu un tampon de bumbac umezit cu alcool.
Infecție pe membrana corion-alantoică. După dezinfectare, ouăle sunt tăiate cu atenție o bucată din coajă de la capătul contondent, membrana cojii este îndepărtată și membrana corion-alantoică este găsită. Materialul infecțios în cantitate de 0,1-0,2 ml folosind o seringă sau o pipetă Pasteur se aplică pe membrana corion-alantoică. După infecție, orificiul este închis cu un capac și spațiul dintre acesta și embrionul puilor este umplut cu parafină.
Pe cealaltă parte a oului, scrieți în creion numele materialului infecțios și data infecției.
Infecție în cavitatea amniotică. Oul este ovoscopat și pe partea laterală este selectat un loc unde corion-alantoida este lipsită de vase de sânge mari. Această zonă este marcată cu un creion. Ouăle sunt așezate pe un suport în poziție orizontală, dezinfectate și o gaură din coajă este străpunsă cu o suliță sterilă specială la o adâncime de 213 mm, prin care un ac cu material infecțios este introdus la aceeași distanță direct în cavitatea amniotică. Pentru a preveni scurgerea lichidului injectat înapoi, se face o puncție deasupra airbag-ului, după care ambele găuri sunt umplute cu parafină.
Infecție în cavitatea alantoică. Infecția se efectuează într-o cutie întunecată. Se observă spațiul aerian, coaja peste spațiul aerian este dezinfectată și un ac de seringă cu materialul este introdus prin orificiul din coajă spre embrion. Dacă acul pătrunde în cavitatea alantoică, atunci umbra embrionului este deplasată. După infecție, gaura este umplută cu parafină.
Infecție în sacul gălbenușului. Coaja este dezinfectată. Oul este așezat pe suport cu capătul tocit în dreapta, astfel încât sacul gălbenușului să fie orientat în sus. O gaură este perforată deasupra camerei de aer din centru. Prin orificiul din coajă într-o direcție orizontală până la o adâncime de 2-3 mm, se introduce un ac de seringă, care intră în sacul gălbenușului. Materialul este injectat într-un volum de 0,2-0,3 ml. După introducerea materialului, gaura este ceruită.
Regimul de temperatură și durata incubației depind de proprietățile biologice ale virusului introdus.
Ouăle infectate sunt verificate zilnic - ovoscopie pentru a verifica viabilitatea embrionului. Dacă embrionii mor în prima zi, atunci cauza este de obicei trauma în timpul infecției. Astfel de ouă sunt derivate din experiență.
Dacă este necesar să se examineze separat fiecare componentă a embrionului, materialul este colectat într-o anumită ordine: lichidul alantoic este aspirat, apoi lichidul amniotic, membrana corion-alantoică este tăiată, membrana amniotică, embrionul, sacul gălbenușului sunt separate și numai atunci membrana corion-alantoică este îndepărtată, separându-l de interior suprafața cochiliei. Prezența virusului în embrionul infectat este determinată de modificările caracteristice ale membranei corion-alantoice a embrionului de pui infectat.
Virușii care nu au activitate de hemaglutinare sunt detectați utilizând RSK.
Pentru a detecta virusul în fluidele alantoice sau amniotice ale embrionilor infectați, se plasează RHA (hemaglutinarea este cauzată de fluide alantoice sau amniotice sau de o suspensie preparată din membrana corion-alantoică).
Cultivarea virușilor în cultura celulară... Pentru acumularea de viruși în culturi celulare sensibile, se utilizează țesuturi ale oamenilor și ale diferitelor animale. Cea mai mare aplicație practică a fost primită de culturile cu un singur strat de linii celulare primare tripsinizate și transplantate.
Culturile celulare cu un singur strat sunt cultivate în vase de saltea plate din sticlă. Suspensia celulară într-un mediu nutritiv lichid la o temperatură de 37 ° C face posibilă obținerea unui strat "in vitro" de celule cu o anumită structură histologică. Prezența virușilor în culturile de țesuturi este detectată prin schimbarea (degenerarea) celulelor. Tipul de viruși este determinat prin neutralizarea efectului virușilor prin adăugarea de seruri specifice de tip adecvate materialului vaccinat.
Aceste metode permit încorporarea mai rapidă a rezultatelor cercetării și sunt mai rentabile. În cazurile în care virușii nu provoacă acțiune citopatică (degenerare) și nu se dezvoltă la embrioni de pui, se utilizează metode de infectare a animalelor (vezi capitolul 11).
Pentru cultivarea virușilor se utilizează celule continue, care se obțin mai des din celule de tumori maligne.
Culturile cu un singur strat se obțin din embrioni umani, de pui, animale.
Avantajul culturilor de celule cu un singur strat este simplitatea tehnicii și ușurința contabilității.
Capacitatea celulelor de a se reproduce în afara corpului este legată de gradul de diferențiere a țesuturilor. Țesuturile mai puțin diferențiate au o capacitate mai mare de proliferare (țesut conjunctiv, epitelial).
Esența metodelor în prepararea culturilor de țesut primar este distrugerea țesutului intercelular și separarea celulelor pentru producerea ulterioară a unui monostrat.
Disocierea celulelor se efectuează acționând asupra țesutului enzimelor proteolitice, cel mai adesea tripsină. Soluția de tripsină promovează separarea celulelor, menținându-și în același timp capacitatea de reproducere. Cultivarea celulelor în creștere necesită un mediu nutritiv. Compoziția mediului este complexă, include o serie de ingrediente: aminoacizi, glucoză, vitamine, săruri minerale, coenzime etc. Cultura țesuturilor se obține în condiții strict aseptice. Antibioticele (500 U de penicilină și 250 U de streptomicină în 1 ml) sunt adăugate la mediu pentru a suprima creșterea florei bacteriene.
Țesutul preparat este turnat cu o soluție 0,25% de tripsină încălzită și incubat într-un termostat la 37 ° C. În timpul incubației, țesutul este agitat periodic prin rotirea balonului. Celulele tripsinizate sunt centrifugate la 800-1000 rpm timp de 5 minute.
Tripsinizarea și centrifugarea se efectuează foarte atent pentru a nu răni celulele. După centrifugare, supernatantul este îndepărtat și sedimentul celular este plasat într-un volum mic de mediu de cultură. Pentru a obține o masă omogenă, suspensia de celule este filtrată printr-un strat de tifon într-o pâlnie (sterilă). Suspensia celulară este verificată pentru sterilitate prin inocularea a 0,1 ml în 2 tuburi cu bulion de zahăr.
Succesul cultivării celulelor depinde de doza de inoculare, prin urmare, după tripsinizare, celulele sunt numărate în camera Goryaev. După numărare, suspensia de celule este diluată cu un mediu nutritiv în așa fel încât 1 ml să conțină 500.000-1.000.000 de celule și să fie turnat în eprubete și saltele. Tuburile de cultură a țesuturilor sunt incubate într-un incubator într-o poziție înclinată.
Culturile inoculate sunt examinate zilnic la un microscop cu mărire redusă pentru a determina natura creșterii lor. Celulele normale care proliferează sunt de culoare deschisă și cresc într-un singur strat. Dacă celulele sunt întunecate, granuloase și nu proliferează, ceea ce poate fi rezultatul contaminării (manipularea slabă a vaselor sau a contaminării ingredientelor), atunci astfel de culturi sunt eliminate din experiment.
Schimbarea mediului nutritiv la 2-3 zile după însămânțare îmbunătățește intensitatea proliferării.
Celulele normale, bine proliferante, sunt infectate cu materialul testat.
Culturile transplantate se obțin în principal din tumori maligne. Tulpina Hela - cultura celulelor cancerului de col uterin ale unei femei pe nume Helena (obținută în 1950); tulpina Hep-2 a fost izolată de la un pacient cu cancer laringian. Creșterea acestor celule este menținută în laboratoare prin pasaje succesive. Particularitatea lor constă în faptul că se reproduc mult timp. În prezent, aceste celule au trecut prin mii de generații. În timpul pasajului, ei pierd unele proprietăți morfologice și biochimice - suferă mutații. Cu toate acestea, ele rămân destul de potrivite pentru cultivarea virusurilor din ele. Cultura acestor celule este utilizată de laboratoare din întreaga lume.
Reproducerea virusului în cultura celulară are loc în momente diferite, în funcție de proprietățile virusului și de tipul de celule.
Prezența virusului este judecată de efectul citopatic. Degenerescența celulară este observată la microscop. Momentul acțiunii citopatice și natura sa depind de doza și proprietățile virusului.
La unele virusuri, un efect citopatic este detectat după câteva zile (virusul variolei), la altele, după 1-2 săptămâni (virusul hepatitei etc.).
În prezent, există deja sute de viruși despre care se știe că infectează oamenii. Lupta împotriva infecțiilor virale se desfășoară prin diferite metode. Imunizarea este cea mai eficientă. În acest fel, variola a fost eliminată și incidența poliomielitei a fost redusă. Prevenirea socială este importantă în lupta împotriva infecțiilor virale - distrugerea câinilor fără stăpân (lupta împotriva rabiei), prevenirea personală etc.
Cu toate acestea, aceste măsuri nu pot asigura eliminarea tuturor bolilor virale. Oamenii de știință caută în mod constant modalități prin care ar fi posibil să se infecteze virusul fără a deteriora celula în care se află.
Prin urmare, este firesc ca în programul Partidului Comunist al Uniunii Sovietice virologia să fie numită una dintre ramurile principale ale științelor naturii, care ar trebui să primească o dezvoltare prioritară în următorii ani.
Metode de bază de cercetare a virușilor... 1. Reacția hemaglutinării, reacția hemaglutinării întârziate, reacția hemaglutinării indirecte. Reacție de legare a complementului.
2. Reacția de neutralizare a virusurilor în cultura țesuturilor.
3. Metoda imunofluorescenței.
4. Metoda histologică - identificarea incluziunilor (corpurile mici ale lui Babesh - Negri - cu rabie; Corpurile mici ale lui Pașen - cu variolă etc.).
5. Metoda biologică.
Infectie virala o celulă este un complex de procese care decurg din interacțiunea unei celule cu un genom viral. Virionul extracelular este inert și această inertitate persistă până când genomul viral începe să funcționeze intracelular, ca unitate genetică independentă. Caracteristica cardinală care stă la baza clasificării date mai jos este relația dintre genomii virusului și celula. Dacă genomul viral se reproduce independent de replicarea genomului celular, acesta este un tip autonom de infecție virală. Dacă genomul viral este inclus în genomul celular și se reproduce împreună cu acesta, acesta este un tip integrativ. Fiecare dintre aceste două tipuri principale de infecție virală poate fi împărțit în mai multe forme în funcție de formarea virusului (infecție productivă și avortă), în funcție de rezultatul procesului infecțios pentru celulă (infecție litică și non-litică), în funcție de dinamica interacțiunii dintre virus și celulă (infecție acută și cronică) ).
Un tip autonom de infecție virală a unei celule este caracteristic majorității virusurilor cunoscute ale animalelor și oamenilor. În acest tip de infecție, genomul viral se reproduce independent de replicarea genomului celular.
O infecție virală autonomă productivă se caracterizează prin formarea unui virus complet. Aceasta este cea mai studiată formă de interacțiune între viruși și celule. Mecanismele de bază ale reproducerii virale, care au fost discutate în capitolul 3, au fost investigate și descrise pentru această formă specială de infecție.
O infecție virală autonomă abortivă apare dacă interacțiunea virusului cu celula nu are ca rezultat formarea de virioni pe deplin valoroși. Acest lucru apare de obicei atunci când celulele sunt infectate cu un virus defect; când virusul pătrunde în celule rezistente la acesta; când celulele sensibile sunt infectate cu un virus pe deplin valoros în condiții non-permisive.
Infecția virală autonomă litică duce la moartea celulară. Virusul care provoacă moartea celulelor se numește citopatogen, iar leziunile celulare sunt denumite prin termenul „citop, efect togenic”. Natura modificărilor citopatologice în infecțiile virale este foarte diversă. Ar putea fi degu
Eșec celular (vacuolizarea citoplasmei, picnoza nucleară, aberații cromozomiale), formarea incluziunilor intracelulare virale specifice, apariția celulelor gigantice, cum ar fi policariocitele sau simplastele. Liza celulară este etapa finală a acțiunii virusurilor citopatogene. Pentru manifestarea citopatogenității, virusul nu necesită reproducerea genomului și implementarea tuturor informațiilor genetice.
O infecție virală autonomă non-litică este caracteristică virușilor care conțin un înveliș exterior lipoproteic și care trimit celule prin înmugurire. Retro, orthomyxo, paramyxo, arenavirusurile sunt capabile să provoace infecții cu celule non-litice productive. Cu toate acestea, în cele mai tipice cazuri, o natură non-litică este caracteristică infecțiilor avortate, când proprietățile genetice ale virușilor necesare implementării funcțiilor citopatogene sunt absente sau nu sunt manifestate.
Conform dinamicii procesului infecțios, infecțiile virale sunt împărțite în acute și cronice. Cand infecție acută în celula infectată are loc formarea virionilor, după care procesul infecțios se oprește. În infecția cronică, celula infectată produce virioni sau componente virale până la moartea sa naturală (nu din cauza unei infecții virale date), în timp ce celulele fiice rămân infectate. Infecția litică productivă și avortă este întotdeauna acută. O infecție non-litică productivă este întotdeauna cronică. Infecția non-litică abortivă, de regulă, are o evoluție cronică, dar poate apărea și sub formă acută.
Tipul integrativ de infecție virală se caracterizează prin uniunea fizică (integrarea) genomului viral și celular. În acest caz, genomul viral se replică și funcționează ca parte integrantă a genomului celular. Acest tip de infecție virală este obligatoriu pentru retrovirusuri și este posibil în cazul virusurilor papa, ado și herpes.
Infecțiile integrative pot fi clasificate în funcție de aceiași parametri ca și cei autonomi - productivitate, dinamică și rezultat; relația de integrare dintre virus și celulă poate fi, de asemenea, productivă și avortă. Infecția integrativă productivă este tipică retrovirusurilor. Avortul, o infecție retrovirală integrativă, are loc în aceleași condiții ca cele descrise mai sus pentru infecțiile autonome avortate. Astfel, retrovirusurile defecte sunt lipsite de
Părți noi ale genomului (unele tulpini ale virusului sarcomului Rous, virușilor sarcomului șoarecilor, pisicilor, maimuțelor, șoarecilor Frend și Rauscher virușii leucemici etc.) pot provoca singuri doar o infecție integrativă abortivă, care se poate transforma într-o infecție productivă în prezența unui virus asistent. Un exemplu de infecție integrativă abortivă cauzată de un retrovirus complet valoros în celulele non-permisive este infecția virusul aviar Sarcomul Rous al celulelor de mamifere. În același timp, are loc un proces de integrare abortivă în celule, care se caracterizează prin scăderea producției de ARN viral, acumularea unui sortiment incomplet de proteine \u200b\u200bvirale, afectarea procesării acestora și lipsa formării virionilor.
De obicei, infecțiile integrative sunt nelitice, ca în cazul retrovirusurilor de tip B, C, D, papov oncogen, adeno, virusuri herpetice, dar există și infecții litice cauzate de retrovirusul citopatogen al reticuloendoteliozei aviare sau virusul „spumant”.
Infecția integrativă virogenă este o infecție avortă care poate fi tradusă într-o formă productivă. Celulele cu această formă de infecție pot spontan sau sub influența unor influențe speciale (fizice, chimice, biologice) să înceapă să producă un virus infecțios. Acest fenomen se numește virogenie, iar celulele sunt desemnate ca virogene, prin analogie cu lizogenia și celulele lizogene din sistemul fago-bacterian.
Mai multe despre subiectul Infecție cu celule virale:
- Infecții virale și infecții cu etiologie virală suspectată
- Infecții bacteriene și virale în aer: gripă, parainfluenză, infecție cu adenovirus, infecție respiratorie sincițială. Bronhopneumonie bacteriană, pneumonie lobară.