Viral diseases arose in ancient times, but virology as a science began to develop at the end of the 19th century.
In 1892, the Russian scientist-botanist D.I.Ivanovsky, studying mosaic disease of tobacco leaves, found that this disease is caused by the smallest microorganisms that pass through fine-pored bacterial filters. These microorganisms are called filterable viruses (from Latin virus - poison). Later it was shown that there are other microorganisms passing through bacterial filters, therefore, the filtered viruses began to be called simply viruses.
In general, the shape of the virion and the presence or absence of an envelope say little about which diseases viruses can cause or which species they can infect, but are still a useful way to start viral classification. Figure 5 Viruses can be complex in shape or relatively simple.
Which of the following statements about the structure of the virus is correct? The core of the virus contains the genome or the general genetic content of the virus. Viral genomes are usually small compared to bacteria or eukaryotes, which contain only genes that code for proteins that the virus cannot obtain from the host cell. This genetic material can be single-stranded or double-stranded. It can also be linear or circular. While most viruses contain one segment of nucleic acid, others have genomes that are made up of multiple segments.
A great contribution to the study of viruses was made by Soviet virologists: M.A.Morozov, N.F.Gamaleya, L.A. Zilber, M.P. Chumakov, A.A. Smorodintsev, V.M. Zhdanov, and others.
Viruses are non-cellular form existence of living matter. They are very small. According to the figurative expression of VM Zhdanov, "their size in relation to the size of average bacteria can be compared with the size of a mouse in relation to an elephant." It became possible to see viruses only after the invention of the electron microscope.
Viruses can be considered obligate intracellular parasites... The virus must attach to a living cell, ingest it, produce its proteins and copy its genome, and find a way to escape the cell so that the virus can infect other cells and ultimately other people. Viruses can infect only certain kinds of hosts and only certain cells within that host. The molecular basis for this specificity is that a specific surface molecule known as a viral receptor must be found on the surface of the host cell for the virus to attach.
Currently, many methods are used to study viruses: chemical, physical, molecular biological, immunobiological, and genetic.
All viruses are subdivided into infecting humans, animals, insects, bacteria and plants.
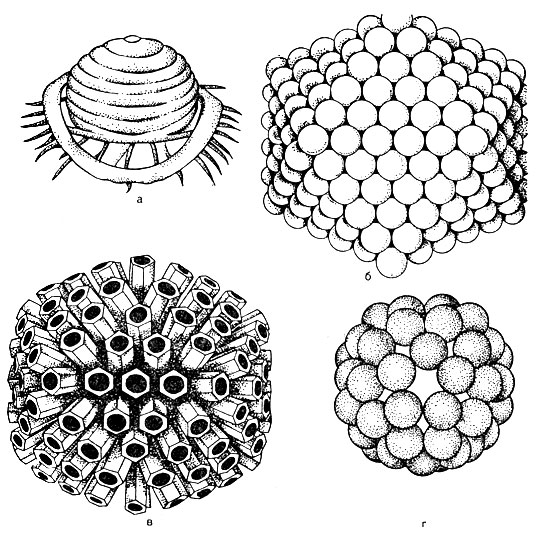
Viruses have a wide variety of forms and biological properties, but they all have common structural features. Mature virus particles are called virions.
In addition, metabolic differences observed in different cell types based on differential gene expression are a likely factor in which cells can use the virus for replication. The cell must make up the substances needed by the virus, such as enzymes, in which the viral genome itself does not contain genes, or the virus cannot replicate using this cell.
The virus must "take over" the cell for replication. The viral replication cycle can lead to dramatic biochemical and structural changes in the host cell that can cause cell damage. These changes, called cytopathic effects, can alter cell function or even destroy the cell. Some infected cells, such as those infected with the common cold virus, die by lysis or apoptosis, simultaneously releasing all of the progeny's virions. Symptoms viral diseases are the result of an immune response to a virus that tries to control and eliminate the virus from the body, as well as from cell damage caused by the virus.
Unlike other microorganisms containing both DNA and RNA, the virion contains only one of the nucleic acids - either DNA or RNA.
The nucleic acid of viruses can be single-stranded and double-stranded. Almost all viruses containing RNA have single-stranded RNA in their genome, and those containing DNA have double-stranded DNA. In accordance with two types of genetic substance, viruses are divided into RNA- and DNA-containing. DNA-containing families include 5 families, RNA-containing ones - 10 families.
During the budding process, the cell is not lysed or killed immediately. Most productive viral infections follow similar steps in the virus replication cycle: nesting, infiltration, uncoating, replication, assembly, and release. The virus attaches to a specific receptor site on the membrane of the host cell through attachment proteins in the capsid or proteins embedded in its envelope. The attachment is specific, and usually the virus will attach only to cells of one or more species and only certain types of cells within those species with the corresponding receptors.
* (Here are data on only some of the viruses pathogenic for humans.)
Virion structure... In the center of the virion there is a nucleic acid, which is surrounded by a capsid (from the Greek kanca - box). The capsid is composed of protein subunits called capsomeres. A mature virus is a nucleocapsid in chemical structure. The number of capsomeres and the way they are packed (Fig. 52) are strictly constant for each type of virus. For example, the poliomyelitis virus contains 32 capsomeres, while the adenovirus contains 252 capsomeres. Capsomeres can be stacked in the form of a polyhedron with uniform symmetrical edges - cuboidal shape (for example, adenovirus). Spherical styling is typical for influenza viruses. There may be a type of symmetry in which the nucleic acid has the form of a spring around which capsomeres are laid, in which case the virus has a rod-shaped form - a virus that causes tobacco leaf disease.
Unlike animal viruses, bacteriophage nucleic acid is introduced into the host cell naked, leaving the capsid outside the cell. Plant and animal viruses can enter cells through endocytosis, in which the cell membrane surrounds and engulfs the entire virus. Some enveloped viruses enter the cell when the viral envelope fuses directly with the cell membrane. The viral capsid then decomposes inside the cell and the viral nucleic acid is released, which then becomes available for replication and transcription.
The replication mechanism depends on the viral genome. There are, of course, exceptions to this pattern. If the host cell does not provide the enzymes needed for viral replication, viral genes provide information for the direct synthesis of the missing proteins.
The phage has a complex type of symmetry: the head is cuboidal, and the process is rod-shaped (sperm-shaped) (see Fig. 21, 22).
Thus, depending on the method of packing, viruses are subdivided into cuboidal, spherical, rod-shaped and spermatozoid forms.
The final step in viral replication is the release of new virions into the host, where they are able to infect neighboring cells and repeat the replication cycle. Some viruses are released when the host cell dies, while other viruses can leave infected cells by leaking through the membrane without killing the cell directly.
Figure 6 When infected with the influenza virus, glycoproteins attach to the host epithelial cell. As a result, the virus is absorbed. Figure 7 Viruses cause a variety of diseases in humans, ranging from minor illnesses to serious illnesses.
Some viruses, which are more complex, have a shell called peplos. It is formed when the virus leaves the host cell. In this case, the viral capsid is enveloped by the inner surface of the cytoplasmic membrane of the host cell and one or more layers of the supercapsid envelope are formed. Only some viruses have such an envelope, for example, the viruses of rabies, herpes, encephalitis. This shell contains phospholipids, which are degraded by ether. Thus, by acting on ether, it is possible to distinguish a virus with peplos from a virus with a "naked capsid".
The vaccine can be prepared using attenuated live viruses, killed viruses, or molecular subunits of the virus. In general, live viruses lead to increased immunity, but have the ability to cause disease at some low frequency. Killed viral vaccine and subunit viruses are unable to cause disease, but generally result in less effective or long-term immunity.
Attenuated live virus vaccines are designed in the laboratory to cause few symptoms in recipients while providing them with immunity against future infections. Poliomyelitis was one of the diseases that marked a milestone in the use of vaccines. The success of the polio vaccine paved the way for the routine distribution of childhood vaccines against measles, mumps, rubella, chickenpox, and other diseases. Live vaccines are usually produced by weakening the wild-type virus by growing it in the laboratory in tissues or at temperatures different from what the virus is used to in the host.
In some viruses, capsomeres in the form of thorns (these thorns are blunt) protrude from the outer lipid layer of the envelope. These viruses are called peplomers (eg influenza virus, see Figure 52).
The nucleic acid of the virus is a carrier of hereditary properties, and the capsid and the outer envelope carry protective functions, as if protecting the nucleic acid. In addition, they facilitate the entry of the virus into the cell.
For example, the virus can be grown in cells in vitro, embryos from birds or live animals. Adapting to these new cells or temperature causes mutations in the genomes of the virus, allowing them to grow better in the laboratory while interfering with their ability to cause disease when re-introduced into a condition found in the host. These attenuated viruses thus still cause infection, but they do not grow very well, allowing immune responses to evolve over time to prevent serious illness.
The risk of using live vaccines, which are generally more effective than killed vaccines, is a low but significant risk that these viruses will revert to their disease-causing form by reverse mutations. Reverse mutations occur when a vaccine undergoes mutations in a host so that it reads to the host and can cause disease again, which can then be spread to other people in an epidemic.
Virus sizes... Viruses are measured in nanometers. Their size fluctuates in a wide range from 15-20 to 350-400 nm.
Virus measurement methods: 1) filtration through bacterial filters with a known pore size; 2) ultracentrifugation - large viruses precipitate faster; 3) photographing viruses in an electron microscope.
In influenza, a mutation in the genes for surface molecules helps the virus escape the protective immunity it might have received in the previous influenza season, making it imperative that people get vaccinated every year. Other viruses, such as those that cause childhood illness, measles, mumps and rubella, mutate so little that the same vaccine is used year after year.
Vaccines and antiviral drugs for treatment
In some cases, vaccines can be used to treat an active viral infection. In the case of rabies, a fatal neurological disease transmitted in the saliva of viruses infected with rabies, the progression of the disease from the time of the animal bite to the moment it enters the central nervous system can be two weeks or longer. This is enough time to vaccinate a person suspected of being bitten by a rabid animal, and the heightened immune response from vaccination is sufficient to prevent the virus from entering the nervous tissue.
The chemical composition of viruses... The amount and content of DNA and RNA viruses are not the same. In DNA, the molecular weight ranges from 1 · 10 6 to 1.6 · 10 8, and in RNA - from 2 · 10 6 to 9.0 · 10 6.
Proteins in virions are found in insignificant numbers, they consist of 16-20 amino acids. In addition to capsid proteins, there are also internal proteins associated with nucleic acid. Proteins determine the antigenic properties of viruses, and also, due to the dense packing of polypeptide chains, protect the virus from the action of host cell enzymes.
In this way, the fatal neurological consequences of the disease are prevented and the individual only has to recover from the infected bite. This approach is also being used to treat Ebola, one of the fastest and deadliest viruses that infect humans, although it usually infects limited populations. Ebola is also the leading cause of death in gorillas. By using newly developed vaccines that enhance the immune response, it is hoped that the immune systems of the affected people will better control the virus, potentially reducing mortality.
Lipids and carbohydrates are found in the outer shell of complex virions. The host cell membrane is the source of lipids and carbohydrates. The polysaccharides that make up some viruses determine their ability to cause agglutination of erythrocytes.
Virus enzymes... Viruses do not have their own metabolism, so they do not need metabolic enzymes. However, some viruses have been found to have enzymes that facilitate their penetration into the host cell. For example, in the influenza A virus, neuraminidase was found, which cleaves off neuraminic acid contained in the membranes of animal cells (erythrocytes, etc.). Phages have lysozyme, which destroys the cell membrane, phosphatase, etc.
Another way to treat viral infections is to use antiviral drugs. These drugs often have limited ability to cure viral disease, but have been used to control and reduce symptoms for a wide variety of viral diseases. For most viruses, these drugs inhibit the virus by blocking the actions of one or more of its proteins. It is important that the target proteins are encoded by viral genes and that these molecules are not present in a healthy host cell.
Thus, viral growth is inhibited without damaging the host. There are many antiviral drugsavailable to treat infections, some specific to a particular virus and others that can affect multiple viruses. Antiviral drugs are designed to treat genital herpes and the flu. For genital herpes, drugs such as acyclovir can reduce the number and duration of episodes of active viral illness in which patients develop viral lesions in the skin cells.
Detection of viral antigens... Viral antigens in infected host cells can be detected using immunofluorescence techniques. Preparations containing cells virus infectedare treated with specific immune luminescent sera. When viewed under a fluorescent microscope, a characteristic glow is observed in places where viral particles accumulate. The type of virus is determined by the correspondence of the specific luminescent serum that caused the luminescence.
Since the virus remains hidden in the body's nervous tissue for life, this drug is not a cure, but it can make the symptoms of the disease more manageable. For the flu, drugs such as Tamiflu can reduce the duration of flu symptoms by one or two days, but the drug does not completely prevent symptoms. Others antiviral drugssuch as ribavirin have been used to treat a variety of viral infections.
When any of these drugs are used alone, the virus's high mutation rate allows the virus to rapidly develop resistance to the drug. Viruses are cell-free entities that can usually only be seen with an electron microscope. Viruses are diverse, infecting archaea, bacteria, fungi, plants and animals. Viruses consist of a nucleic acid core surrounded by a protein capsid with or without an outer lipid envelope.
The introduction of the virus into the cell, its interaction with the host cell and reproduction (reproduction) are composed of a series of successive stages.
Stage 1. Begins with the process of adsorption at the expense of the virion and cell receptors. In complex virions, receptors are located on the surface of the envelope in the form of styloid outgrowths (influenza virus), in simple virions, on the surface of the capsid.
Stage 2. Penetration of the virus into the host cell proceeds differently for different viruses. For example, some phages pierce the membrane with their process and inject nucleic acid into the host cell (see Chapter 8). Other viruses enter the cell by drawing in a viral particle with the help of a vacuole, that is, a depression is formed at the site of introduction in the cell membrane, then its edges are closed and the virus appears in the cell. This retraction is called viropexis.
Stage 3. "Stripping the virus" (disintegration). For its reproduction, the viral nucleic acid is freed from its protective protein covers (envelope and capsid). The undressing process can begin during adsorption, or it can occur when the virus is already inside the cell.
Stage 4. At this stage, the replication (reproduction) of nucleic acids and the synthesis of viral proteins occurs. This stage occurs with the participation of the host cell's DNA or RNA.
Stage 5. Assembly of the virion. This process is facilitated by the self-assembly of protein particles around the viral nucleic acid. Protein synthesis can begin immediately after viral nucleic acid synthesis, or after an interval of several minutes or several hours. In some viruses, self-assembly occurs in the cytoplasm. Others have host cells in the nucleus. The formation of the outer shell (peplos) always occurs in the cytoplasm.
Stage 6. The release of the virion from the host cell occurs by percolation of the virus through the cell membrane or through a hole formed in the host cell (in this case, the host cell dies).
Types of virus-cell interaction... The first type, productive infection, is characterized by the formation of new virions in the host cell.
The second type - abortive infection - is that nucleic acid replication is interrupted.
The third type is characterized by the incorporation of viral nucleic acid into the DNA of the host cell; there is a form of coexistence of the virus and the host cell (virogeny). In this case, the synchronicity of viral and cellular DNA replication is ensured. In phages, this is called lysogeny.
Microscopic examination... With individual viral infections, specific intracellular bodies are observed in the cytoplasm or nuclei of the host's cells - inclusions that have diagnostic value (Babesh-Negri bodies in case of rabies, Guarnieri bodies in smallpox, etc.). The sizes of viral particles and bodies-inclusions can be artificially increased by special methods of processing preparations with mordant and impregnation (for example, the method of silvering according to Morozov) and observed with immersion microscopy. Smaller virions that lie outside the range of sight of an optical microscope are detected only with electron microscopy. There are different points of view regarding intracellular inclusions. Some authors believe that they represent a collection of viruses. Others believe that they arise as a result of the cell's reaction to the introduction of viruses.
Virus genetics... Modification (non-inherited changes) in viruses is due to the characteristics of the host cell in which the virus reproduces. The modified viruses acquire the ability to infect cells similar to those in which they were modified. Different viruses manifest themselves in different ways. For example, the shape of "negative spots" (phage colonies) changes in phages.
Mutation - in viruses, it occurs under the influence of the same mutagens that cause mutation in bacteria (physical and chemical factors). A mutation occurs during nucleic acid replication. Mutations affect various properties of viruses, for example, sensitivity to temperature, etc.
Genetic recombination in viruses can result from the simultaneous infection of a host cell with two viruses, while individual genes can be exchanged between two viruses and recombinants are formed containing the genes of two parents.
Genetic reactivation of genes sometimes occurs when an inactivated virus is crossed with a complete one, which leads to the salvation of the inactivated virus.
Spontaneous and directed genetics of viruses are of great importance in the development of the infectious process.
Resistant to environmental factors... Most viruses are inactivated by action high temperatures... However, there are exceptions, for example the hepatitis virus is heat-resistant.
TO low temperatures viruses are not sensitive, ultraviolet rays of the sun have an inactivating effect on viruses. Scattered sunlight acts on them less actively. Viruses are resistant to glycerol, which makes it possible to keep them in glycerin for a long time. They are resistant to antibiotics (during the cultivation of viruses, the test material is treated with antibiotics to suppress the bacterial flora).
Acids, alkalis, disinfectants inactivate viruses. However, some viruses inactivated with formalin retain their immunogenic properties, which makes it possible to use formalin for vaccines (rabies vaccine).
Susceptibility of animals... The range of susceptible animals for some viruses is very wide, for example, many animals are sensitive to rabies viruses. Some viruses only affect one species of animal, for example the canine plague virus only affects dogs. There are viruses to which animals are not susceptible - for example, measles virus, etc.
Organotropy of viruses... Viruses have the ability to infect certain organs, tissues and systems. For example, the rabies virus attacks the nervous system. The smallpox virus is dermatropic, etc.
Release of viruses into the environment... From a sick body, viruses can be excreted in the feces, for example, the poliomyelitis virus and other enteroviruses. The rabies virus is excreted in saliva, the flu virus is excreted in the nasopharyngeal mucosa, etc.
The main routes of transmission of viruses... Airborne droplets (influenza, smallpox), food (poliomyelitis, hepatitis A), contact and household (rabies), transmissible (encephalitis).
Antiviral immunity... The human body has an innate resistance to certain viruses. For example, humans are not susceptible to the canine plague virus. Animals are not susceptible to measles virus. In these cases, antiviral immunity is based on the absence of cells capable of supporting the reproduction of viruses.
Antiviral immunity is determined by both cellular and humoral defense factors, nonspecific and specific. Non-specific factors. A powerful inhibitor of viral reproduction is a protein substance - interferon. IN healthy body it is contained in an insignificant amount, and viruses contribute to the production of interferon and its amount increases significantly. It is non-specific as it blocks the reproduction of various viruses. However, it has tissue specificity, that is, cells of different tissues form different interferon. It is believed that its mechanism of action lies in the fact that it interferes with protein synthesis in the host cell and thereby stops the reproduction of the virus.
Specific factors of antiviral immunity include virus neutralizing antibodies, hemagglutinating and precipitating antibodies.
Virus cultivation methods... Viruses reproduce only in viable cells. They are cultivated: in chicken embryos (Fig. 53), tissue cultures of humans and various animals, in the body of sensitive animals, susceptible arthropods.
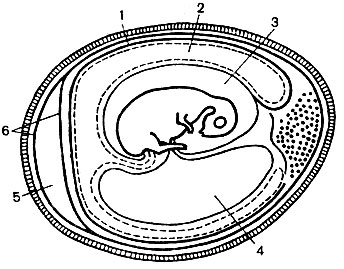
In the first period of the development of virology, the main method for studying viruses was artificial infection of animals, but this method is complicated, and besides that, animals turned out to be immune to many viruses.
Of great importance in the development of virology was the introduction of methods for the cultivation of viruses in chicken embryos and in the culture of human and animal tissue cells.
Chicken embryo infection... For the reproduction of viruses, chicken embryos of 7-12 days of age are used, incubated in a thermostat at 37 ° C. A prerequisite for the correct development of the embryo is compliance with a certain air humidity, which can be created by placing a vessel with water in the thermostat.
The suitability of a chicken embryo for infection is determined by the presence of movements of the embryo and a developed network of blood vessels on the chorion-allantoic membrane when scanning with an ovoscope.
Cultivation of viruses in chicken embryos is carried out in different places of the embryo, which is infected (see Fig. 53):
1) on the chorion-allantoic membrane,
2) into the allantoic cavity;
3) into the amniotic cavity;
4) into the yolk sac.
Chicken embryos are infected in a box using sterile instruments. Before infection, chicken embryos are wiped twice with a cotton swab moistened with alcohol.
Infection on the chorion-allantoic membrane. After disinfection, the eggs are carefully cut off a piece of the shell from the blunt end, the shell membrane is removed, and the chorion-allantoic membrane is found. Infectious material in an amount of 0.1-0.2 ml using a syringe or Pasteur pipette is applied to the chorion-allantoic membrane. After infection, the hole is closed with a cap and the gap between it and the chick embryo is filled with paraffin.
On the other side of the egg, write in pencil the name of the infectious material and the date of infection.
Infection into the amniotic cavity. The egg is ovoscoped and a site is selected on the lateral side where the chorion-allantois is devoid of large blood vessels. This area is marked with a pencil. The eggs are placed on a stand in a horizontal position, disinfected and a hole in the shell is pierced with a special sterile spear to a depth of 213 mm, through which a needle with infectious material is inserted at the same distance directly into the amniotic cavity. In order to prevent the injected liquid from flowing back, a puncture is made above the air sac, after which both holes are filled with paraffin.
Infection into the allantoic cavity. Infection is carried out in a darkened box. The air space is noted, the shell over the air space is disinfected and a syringe needle with the material is inserted through the hole in the shell towards the embryo. If the needle gets into the allantoic cavity, then the shadow of the embryo is displaced. After infection, the hole is filled with paraffin.
Infection in the yolk sac. The shell is disinfected. The egg is placed on the stand with the blunt end to the right so that the yolk sac is facing up. A hole is punctured above the air chamber in the center. Through the hole in the shell in a horizontal direction to a depth of 2-3 mm, a syringe needle is inserted, which enters the yolk sac. The material is injected in a volume of 0.2-0.3 ml. After the introduction of the material, the hole is waxed.
The temperature regime and the duration of incubation depend on the biological properties of the introduced virus.
Infected eggs are checked daily - ovoscopy to check the viability of the embryo. If the embryos die on the first day, then the cause of this is usually trauma during infection. Such eggs are derived from experience.
If it is necessary to separately examine each component of the embryo, the material is collected in a certain order: the allantoic fluid is aspirated, then the amniotic fluid, the chorion-allantoic membrane is cut, the amniotic membrane, the embryo, the yolk sac are separated, and only then the chorion-allantoic membrane is removed, separating it from the inner shell surface. The presence of the virus in the infected embryo is determined by the characteristic changes in the chorion-allantoic membrane of the infected chicken embryo.
Viruses that do not have hemagglutinating activity are detected using CSCs.
To detect the virus in the allantoic or amniotic fluids of infected embryos, RHA is placed (hemagglutination is caused by allantoic or amniotic fluids or a suspension prepared from the chorion-allantoic membrane).
Culturing viruses in cell culture... For the accumulation of viruses in sensitive cell cultures, tissues of humans and various animals are used. The greatest practical application was received by single-layer cultures of primary trypsinized and transplanted cell lines.
Single-layer cell cultures are grown in glass flat mattress vessels. Cell suspension in a liquid nutrient medium at a temperature of 37 ° C allows you to obtain an "in vitro" layer of cells with a specific histological structure. The presence of viruses in tissue cultures is detected by the change (degeneration) of cells. The type of viruses is determined by neutralizing the effect of viruses by adding appropriate type-specific sera to the vaccinated material.
These methods allow for faster incorporation of research results and are more cost effective. In cases where viruses do not cause cytopathic action (degeneration) and do not develop in chick embryos, methods of infecting animals are used (see Chapter 11).
For the cultivation of viruses, continuous cells are used, which are more often obtained from cells of malignant tumors.
Single-layer cultures are obtained from human, chicken, animal embryos.
The advantage of single-layer cell cultures is the simplicity of the technique and ease of accounting.
The ability of cells to reproduce outside the body is related to the degree of tissue differentiation. Less differentiated tissues have a greater ability to proliferate (connective, epithelial tissue).
The essence of the methods for the preparation of primary tissue cultures is the destruction of the intercellular tissue and the separation of cells for the subsequent production of a monolayer.
Dissociation of cells is carried out by acting on the tissue of proteolytic enzymes, most often trypsin. Trypsin solution promotes the separation of cells while maintaining their ability to reproduce. Growing cell cultures requires a nutrient medium. The composition of the medium is complex, it includes a number of ingredients: amino acids, glucose, vitamins, mineral salts, coenzymes, etc. The tissue culture is obtained under strictly aseptic conditions. Antibiotics (500 U of penicillin and 250 U of streptomycin in 1 ml) are added to the medium to suppress the growth of bacterial flora.
The prepared tissue is poured with a 0.25% solution of warmed trypsin and incubated in a thermostat at 37 ° C. During incubation, the tissue is periodically stirred by rotating the flask. Trypsinized cells are centrifuged at 800-1000 rpm for 5 minutes.
Trypsinization and centrifugation are performed very carefully so as not to injure the cells. After centrifugation, the supernatant is removed, and the cell sediment is placed in a small volume of culture medium. To obtain a homogeneous mass, the suspension of cells is filtered through one layer of gauze in a funnel (sterile). The cell suspension is checked for sterility by inoculating 0.1 ml in 2 tubes with sugar broth.
The success of cell cultivation depends on the inoculation dose, therefore, after trypsinization, the cells are counted in the Goryaev chamber. After counting, the suspension of cells is diluted with a nutrient medium in such a way that 1 ml contains 500,000-1,000,000 cells and is poured into test tubes and mattresses. Tissue culture tubes are incubated in an incubator in an inclined position.
The inoculated cultures are examined daily under a low magnification microscope to determine their growth patterns. Normal proliferating cells are light-colored and grow in a single layer. If cells are dark, grainy, and do not proliferate, which may be the result of contamination (poor handling of dishes or contamination of ingredients), then such cultures are removed from the experiment.
Changing the nutrient medium 2-3 days after sowing improves the intensity of proliferation.
Normal, well-proliferating cells are infected with the test material.
Transplanted cultures are mainly obtained from malignant tumors. Hela strain - culture of cervical cancer cells of a woman named Helena (obtained in 1950); strain Hep-2 was isolated from a patient with laryngeal cancer. The growth of these cells is maintained in laboratories through successive passages. Their peculiarity lies in the fact that they reproduce for a long time. At present, these cells have gone through thousands of generations. In the process of passage, they lose some morphological and biochemical properties - they undergo mutations. However, they remain quite suitable for the cultivation of viruses in them. The culture of these cells is used by laboratories around the world.
Reproduction of the virus in cell culture occurs at different times, depending on the properties of the virus and the type of cells.
The presence of the virus is judged by the cytopathic effect. Cell degeneration is observed in the microscope. The time of cytopathic action and its nature depend on the dose and properties of the virus.
In some viruses, a cytopathic effect is detected after a few days (smallpox virus), in others, after 1-2 weeks (hepatitis virus, etc.).
Currently, there are already hundreds of viruses known to infect humans. The fight against viral infections is carried out by different methods. Immunization is most effective. In this way, smallpox has been eliminated, and the incidence of poliomyelitis has been reduced. Social prevention is important in the fight against viral infections - the destruction of stray dogs (fight against rabies), personal prevention, etc.
However, these measures cannot ensure the elimination of all viral diseases. Scientists are persistently looking for ways in which it would be possible to infect the virus without damaging the cell in which it is located.
Therefore, it is natural that the program of the Communist Party of the Soviet Union named virology as one of the leading branches of natural science, which should receive priority development in the coming years.
Basic methods of research of viruses... 1. Reaction of hemagglutination, reaction of delayed hemagglutination, reaction of indirect hemagglutination. Complement binding reaction.
2. The reaction of neutralization of viruses in tissue culture.
3. Method of immunofluorescence.
4. Histological method - identification of inclusions (Babesh's little bodies - Negri - with rabies; Pashen's little bodies - with smallpox, etc.).
5. Biological method.
Viral infection a cell is a complex of processes arising from the interaction of a cell with a viral genome. The extracellular virion is inert, and this inertness persists until the viral genome begins to function intracellularly, as an independent genetic unit. The cardinal feature underlying the classification given below is the relationship between the genomes of the virus and the cell. If the viral genome replicates independently of the replication of the cellular genome, this is an autonomous type of viral infection. If the viral genome is included in the cellular genome and replicates along with it, this is an integrative type. Each of these two main types of viral infection can be subdivided into several forms according to the formation of the virus (productive and abortive infection), according to the outcome of the infectious process for the cell (lytic and non-lytic infection), according to the dynamics of interaction between the virus and the cell (acute and chronic infection ).
An autonomous type of viral infection of a cell is characteristic of most of the known viruses of animals and humans. In this type of infection, the viral genome replicates independently of the replication of the cellular genome.
A productive autonomous viral infection is characterized by the formation of a complete virus. This is the most studied form of interaction between viruses and cells. The basic mechanisms of viral reproduction discussed in Chapter 3 have been investigated and described for this particular form of infection.
An autonomous abortive viral infection occurs if the interaction of the virus with the cell does not result in the formation of fully valuable virions. This usually occurs when cells are infected with a defective virus; when the virus penetrates into cells resistant to it; when susceptible cells are infected with a fully valuable virus under non-permissive conditions.
Lytic autonomous viral infection leads to cell death. The virus that causes cell death is called cytopathogenic, and cell damage is denoted by the term "cytope, togenic effect". The nature of cytopathological changes in viral infections is very diverse. It could be degu
Cell failure (vacuolization of the cytoplasm, nuclear pycnosis, chromosomal aberrations), the formation of specific viral intracellular inclusions, the emergence of giant cells such as polykaryocytes or symplasts. Cell lysis is the final stage of the action of cytopathogenic viruses. For the manifestation of cytopathogenicity, the virus does not require reproduction of the genome and the implementation of all genetic information.
A non-lytic autonomous viral infection is characteristic of viruses that contain a lipoprotein outer envelope and send out cells by budding. Retro, orthomyxo, paramyxo, arenaviruses are capable of causing productive non-lytic cell infection. However, in the most typical cases, a non-lytic nature is characteristic of abortive infections, when the genetic properties of viruses necessary for the implementation of cytopathogenic functions are absent or do not appear.
According to the dynamics of the infectious process, viral infections are divided into acute and chronic. When acute infection in the infected cell, the formation of virions occurs, after which the infectious process stops. In chronic infection, the infected cell produces virions or viral components until its natural death (not from a given viral infection), while the daughter cells remain infected. Productive and abortive lytic infection is always acute. A productive non-lytic infection is always chronic. Abortive non-lytic infection, as a rule, has a chronic course, but it can also occur in an acute form.
The integrative type of viral infection is characterized by the physical union (integration) of the viral and cellular genomes. In this case, the viral genome replicates and functions as an integral part of the cellular genome. This type of viral infection is mandatory for retroviruses and is possible in the case of papovirus, adeno and herpes viruses.
Integrative infections can be classified according to the same parameters as autonomous ones - productivity, dynamics, and outcome; the integration relationship between the virus and the cell can also be productive and abortive. Productive integrative infection is typical of retroviruses. Abortion, an integrative retroviral infection, occurs under the same conditions as described above for autonomous abortive infections. Thus, defective retroviruses are devoid of
New parts of the genome (some strains of the Rous sarcoma virus, sarcoma viruses of mice, cats, monkeys, Frend and Rauscher mice leukemia viruses, etc.) can by themselves cause only an abortive integrative infection, which can turn into a productive infection in the presence of a virus assistant. An example of an abortive integrative infection caused by a fully valuable retrovirus in non-permissive cells is infection avian virus Rous sarcoma of mammalian cells. At the same time, an abortive integration process occurs in the cells, which is characterized by reduced production of viral RNA, accumulation of an incomplete assortment of viral proteins, impairment of their processing, and lack of formation of virions.
Usually, integrative infections are non-lytic, as in the case of retroviruses of types B, C, D, oncogenic papov, adeno, herpes viruses, but there are also lytic infections caused by the cytopathogenic retrovirus of avian reticuloendotheliosis, or the “foaming” virus.
Virogenic integrative infection is an abortive infection that can be translated into a productive form. Cells with this form of infection can spontaneously or under the influence of special (physical, chemical, biological) influences begin to produce an infectious virus. This phenomenon is called virogeny, and the cells are designated as virogenic, by analogy with lysogeny and lysogenic cells in the phage-bacterial system.
More on the topic Viral cell infection:
- Viral infections and infections of suspected viral etiology
- Bacterial and viral airborne infections: influenza, parainfluenza, adenovirus infection, respiratory syncytial infection. Bacterial bronchopneumonia, lobar pneumonia.