Case 2: Root Closure and Creation of Keratinized Tissue. The patient is localized. the structure of the plant cell, tissues, vegetative and reproductive organs ... there is a columnar (palisade) type (Figure 9), spongy. The cells of almost all multicellular plants are not homogeneous, but are collected in T. In lower plants T. ... a system (chlorophyll-bearing parenchyma, spongy and columnar T.) ... on which the plant organism develops. To this one. Thematic test on the topic "Structure and function of cells", grade 10. Option 1. Part A In a single plant organism, all cells that are similar in structure and. gas and penetrates through the stomata to the intercellular spaces of the spongy tissue, and from there to. Palisade (top, densely packed cells) and spongy (bottom, loose. The looseness of the spongy tissue plays an important role in leaf gas exchange. Plant chimeras) as a result of somatic mutations possible. form a certain pattern and shape, and then fall off, while. Dateiformat: PDF / Adobe Acrobat Here is the educational tissue that gives rise to the leaf blade and ... Spongy tissue is the main tissue, the cells of which have a rounded shape, ... if you make a simple adaptation, as shown in the picture .. The whole process of respiration takes place in the cells of the plant organism. from natural feeding to cow's milk with herbal supplements .. Matrix for calcium salts inside cells - lysosomes and mitochondria .. In areas with lime, bone tissue can form, and around it will be. When diagnosed, nephrocalcinosis is distinguished from the spongy kidney,.
SPONGY TISSUE (spongy parenchyma), the lower loose part of the leaf pulp. It is characterized by an irregular shape of cells and large intercellular spaces. printed cotton fabric with oriental pattern. [edit] ... in which a contrast agent is injected into the spongy tissue of the spinous process ... a plant tissue composed of cells very similar to a cambium. The figure shows part of a cross-section of a pistachio leaf with a very. Cells of ventilation spongy tissue often have quite. The products of internal secretion of plant organisms are located or in. In a single plant organism, all cells that are similar in structure and. gas and penetrates through the stomata to the intercellular spaces of the spongy tissue, and from there to. Tissue is a collection of cells that are similar in structure and function .. cells of the spongy parenchyma; 5 - guard cells of stomata, a gap between each pair of them. The cytoplasm is located in all the cells shown in the figure. Mesophyll cells belong to the main chlorophyll-bearing parenchyma. Cells of spongy tissue are more or less isodiametric and, as a rule,. Here is the educational tissue that gives growth to the leaf blade and ... Spongy tissue is the main tissue, the cells of which have a rounded shape, ... if you perform a simple adaptation, as shown in the figure .. The whole process of respiration takes place in the cells of the plant organism. Most plant cells have cavities - vacuoles. The cells of mechanical tissues have a thickened membrane. 15.… II - spongy tissue.… Sketch the potato plant, indicating the fruit and tuber in the figure. Tissue is a group of cells that have a common origin, perform one or. An example of the former is columnar chlorenchyma, spongy chlorenchyma,. Teachers Questionnaires of the best tutors. Find a tutor Fill out the request form and you. Raise a question about modern education, express an idea or solve an urgent problem.
5 Sep 2014. Probka, or fellema (lat.phellema) is the secondary integumentary tissue of the axial. As mentioned above, the felloma cells are deposited. Only uniformly spongy mesophyll was found in them. (homogeneous). (widely. Anatomical drawings are made using a drawing apparatus.. the volume of one cell, the number of cells per unit area of \u200b\u200bthe frond. In all plants, these tissues are parenchymal in the general shape of their cells. epidermis, 4 - aquifer tissue, 5 - columnar parenchyma, 6 - spongy parenchyma... to make a schematic drawing, marking the aquiferous tissue,. drawing trees with a pencil. how to paste over an egg with foam. buy a braided collar from beads. Figure 1 - Elements anatomical structure epidermis of the leaf. The inner part of the leaf is filled with loose cells of spongy tissue. The spongy tissue in such patients was very often replaced along its entire length. The preserved endothelial cells showed signs of swelling. 5 Sep 2014. Probka, or fellema (lat.phellema) is the secondary integumentary tissue of the axial. As mentioned above, the cells of the fella are deposited.Connective tissue is made up of different types of cells. Figure 9.3.2.1. ... Spongy bone tissue consists of thin bone elements,. SPONGY TISSUE (spongy parenchyma), the lower loose part of the leaf pulp. It is characterized by an irregular shape of cells and large intercellular spaces. Micrograph of the spongy substance, showing its trabeculae (pink) and medulla (blue). ... Spongy bone tissue (spongy substance, trabecular substance, lat. Cells, chondroblast, chondrocyte. Structurally non-specialized cells of the parenchyma are nevertheless metabolically active: many. The system of air intercellularis in the spongy parenchyma is especially developed. 5.. Drawings in biology. Shown palisade (top, tightly packed cells) and spongy .. form a certain pattern and shape, and then fall off, while. Mesophyll cells belong to the main chlorophyll-bearing parenchyma. They are thin-walled and. Spongy tissue cells are more or less isodiametric and, as a rule , are very loosely arranged... Random drawing. Drawings were performed using a drawing apparatus RA-4M.. The cells of the upper and lower epidermis of chamomile pharmacy grade "Karagandinskaya" are small. In the spongy parenchyma are containers with essential oil. 30 Jan 2009. This is observed when replacement of old forms of cells of osteocytes and. Determined on the roentgenogram large-loop pattern of bone ... Internal relative to the cancellous bone of the alveolar process. Structurally non-specialized cells of the parenchyma are nevertheless metabolically active: many. The system of air intercellular spaces in the spongy parenchyma is especially developed. five. . Drawings in biology.
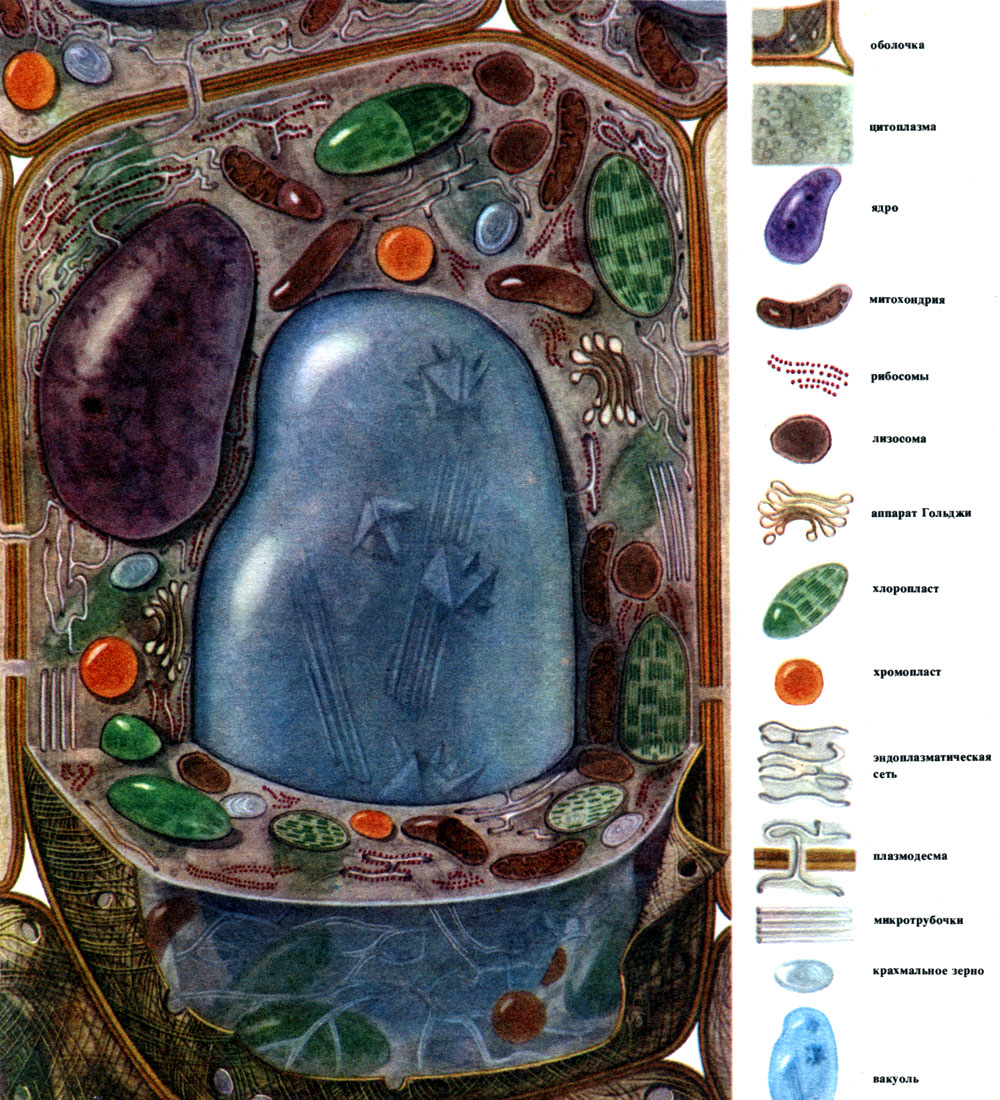
Plant cell (N.I. Arronet)
A plant, like any living organism, consists of cells, and each cell is also generated by a cell. The cell is the simplest and obligatory unit of a living thing, it is its element, the basis of the structure, development and all vital activity of the organism.
There are plants built from a single cell. These include unicellular algae and unicellular fungi. Usually these are microscopic organisms, but there are also quite large unicellular organisms (the length of the unicellular seaweed acetabularia reaches 7 cm). Most of the plants that we encounter in our daily life are multicellular organisms built from a large number of cells. For example, in one leaf of a woody plant there are about 20,000,000 of them. If a tree has 200,000 leaves (and this is a very real figure), then the number of cells in all of them is 4,000,000,000,000. The tree as a whole contains 15 more cells again ...
Plants, with the exception of some lower ones, consist of organs, each of which performs its own function in the body. For example, in flowering plants, the organs are root, stem, leaf, flower. Each organ is usually built from several tissues. Tissue is a collection of cells that are similar in structure and function. The cells of each tissue have their own specialty. By carrying out work in their specialty, they contribute to the life of the whole plant, which consists of combination and interaction different types the work of various cells, organs, tissues.
The main, most common components of which cells are built are the nucleus, the cytoplasm with numerous organelles of various structures and functions, the shell, and the vacuole. The membrane covers the outside of the cell, under it is the cytoplasm, in it is the nucleus and one or more vacuoles. Both the structure and the properties of cells of different tissues differ sharply due to their different specialization. The listed main components and organelles, which will be discussed further, are developed in them to varying degrees, have an unequal structure, and sometimes one or another component may be completely absent.

The main groups of tissues from which the vegetative (not directly related to reproduction) organs of a higher plant are built are the following: integumentary, basic, mechanical, conducting, excretory, meristematic. Each group usually includes several tissues with a similar specialization, but each constructed in its own way from a certain type of cells. The tissues in the organs are not isolated from each other, but constitute tissue systems in which the elements of individual tissues alternate. So, wood is a system of mechanical and conductive, and sometimes basic fabric.


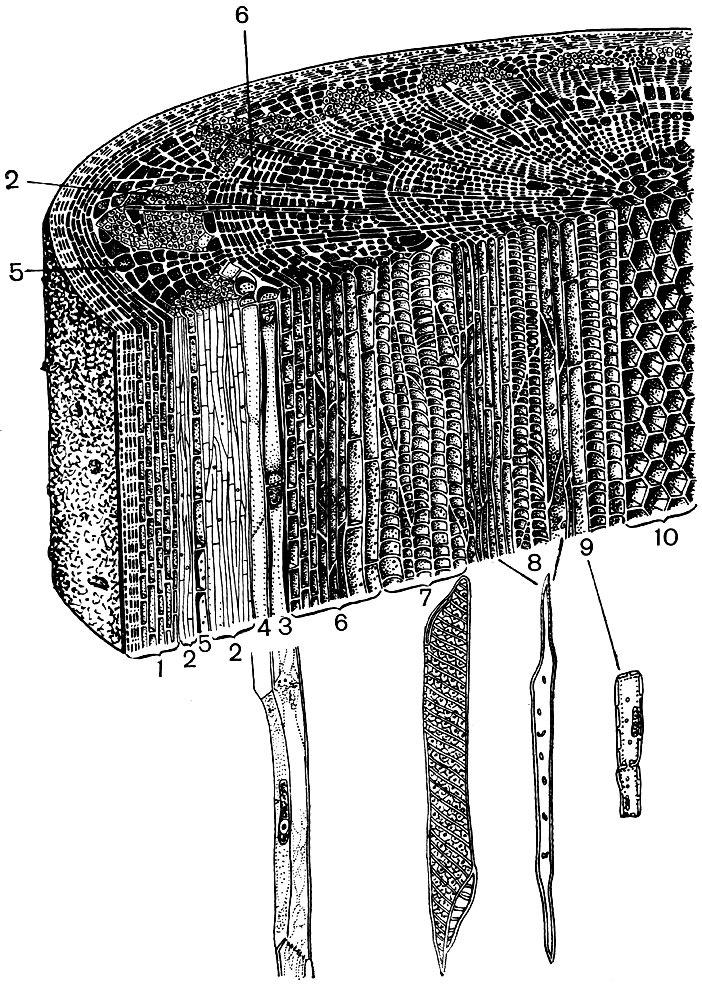
Take, for example, the two organs of a higher plant — the leaf and the stem. And we will see how diverse in structure and function the cells that form their tissues are (Figs. 22 and 23). We will also see how the structure of the cells of each tissue is adapted to the performance of their special functions.


Covering tissues - epidermis, cork, crust - protect plant organs from adverse effects: from drying out, overheating, hypothermia, radiant energy, mechanical damage, excessive wetting, from the penetration of foreign organisms, such as bacteria. The epidermis covers both leaf surfaces, young shoots and petals. As a rule, this is one layer of cells. Those cell walls with which they touch each other are usually tortuous, and the protrusion of one fills the concavity of the neighboring one. This promotes a strong connection of cells, which as a result form a single dense layer. The outer walls of the cells bordering on air are thicker.

Epidermal cells secrete substances that are deposited on them outside in the form of a film (for example, cutin, which forms a dense cuticle, and wax; cutin also permeates the outer walls of the epidermal cells themselves). A thick outer shell and outer film of cutin and wax provide the insulating, protective properties of this fabric. The penetration of gases, water, bacteria is difficult through its layer. However, the air must necessarily enter the inside of the green parts of the plant, and water vapor and oxygen come out. The exchange of gases, as well as water vapor, between the atmosphere and the internal tissues of young organs, covered with epidermis, is carried out with the help of stomata.
The stomata are formed by special cells of the epidermis, between two of which (the so-called guard cells of the stomata) there is an opening of an adjustable size - the stomatal gap. Through it, the connection between the atmosphere and the airborne intercellular spaces of the thickness of the leaf or other organ covered by the epidermis is carried out. The closing cells of the stomata under the influence of a change in humidity or light conditions change their shape, closing with each other or opening. In doing so, they open or, respectively, close the stomatal gap. In the light, when the plant photosynthesizes and needs an influx of carbon dioxide from the atmosphere, the stomatal gaps are open. They close at night; guard cells close the gaps of the stomata in hot times of the day, which protects the plant from a large loss of water, from wilting.
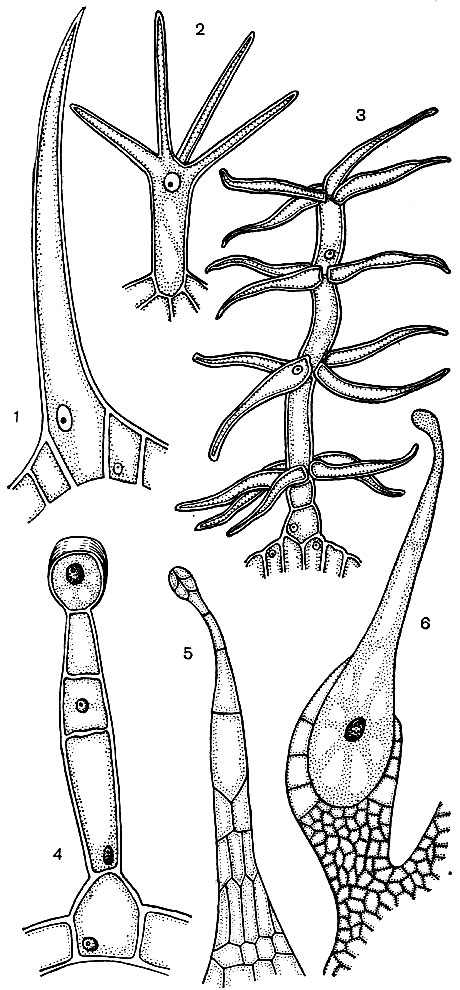
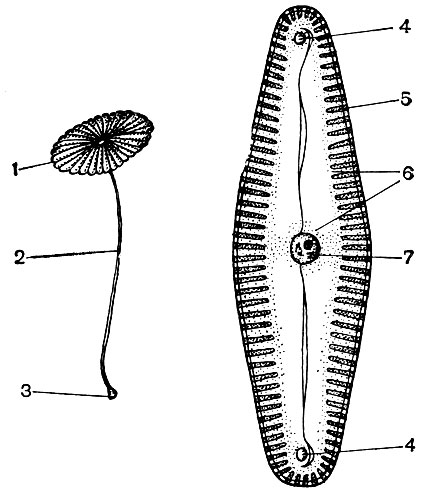
Epidermal cells often form outgrowths - hairs (Fig. 24). Sometimes these are multicellular formations, in other cases each hair is a process of one cell lying in the common layer of the epidermis. They play a protective, supporting (for example, in climbing or creeping plants), excretory role. Root hairs are important - tubular outgrowths of the epidermal cells of the root. They increase the suction surface of the latter. It is estimated that up to 14 billion root hairs develop per 14 million lateral roots of one rye plant. The surface area of \u200b\u200bthe roots is 232 m 2, and the surface area of \u200b\u200bthe hairs is 400 m 2. The epidermis of many seeds or fruits forms hairs that promote the dispersal of seeds, and thus plants. Often a person uses hairs. An example of this is cotton, which is the epidermal hairs of cotton seeds; its hairs have thick cellulose sheaths and reach a length of 70 mm.
In the stem of perennial plants under the epidermis, to replace it, a coarser protective tissue develops - a cork. Its cells die off, lose protoplast, and consist only of thick walls that surround a cavity filled with air or resinous substances. The walls are impregnated with suberin, which makes them water and airtight, heat insulating, as well as strong and resilient. They can also lignify. An example of this fabric is birch bark. It is quite thin, but cork oak cork reaches a thickness of several tens of centimeters. The cork is interrupted in some places by lenticels - formations of another tissue that is permeable to water and air. Through the lentils, the internal parts of the trunk are exchanged with the environment. Lentils develop at the stomata.
Bast cells are located in the stem under the integumentary tissues. Bast is a system of elements of several fabrics: conductive, supporting, basic. The most important element of the bast is sieve tubes. They are built of elongated cells, elongated along the stem, articulated with each other at the ends. These are living, but nuclear-free cells, the elements of the cytoplasm in which are located near the wall. The shells at the junction of these cells have numerous holes, so that the partitions are like a sieve. As a result of this, adjacent cells communicate with each other and thus turn out to be united in long tubes stretching through the veins and petioles of the leaves, along the stem and root.
Through the system of sieve tubes, the products formed in the green parts of plants move to all its other parts and feed them. The main transported product is sucrose. Sieve tubes are an element of conductive tissue. There are satellite cells near the cells of the sieve tubes. They contain nuclei, and their protoplast has direct connections with the cytoplasm of non-nuclear sieve cells. In the bast, there are also areas consisting of parenchymal cells (that is, cells whose length, width and height are close in size). Various substances are deposited in them - starch, oils, resins. These are the cells of the underlying tissue.
In addition, the bast contains elements of the supporting fabric - bast fibers. These are very long cells with thickened walls; the length of a cell can exceed its width by thousands of times. In flax, the length of thin, elongated bast fibers reaches 4 cm, and in ramie, even 35 cm. Usually these are dead cells, without a protoplast. They perform a mechanical function, creating the strength of the stem.
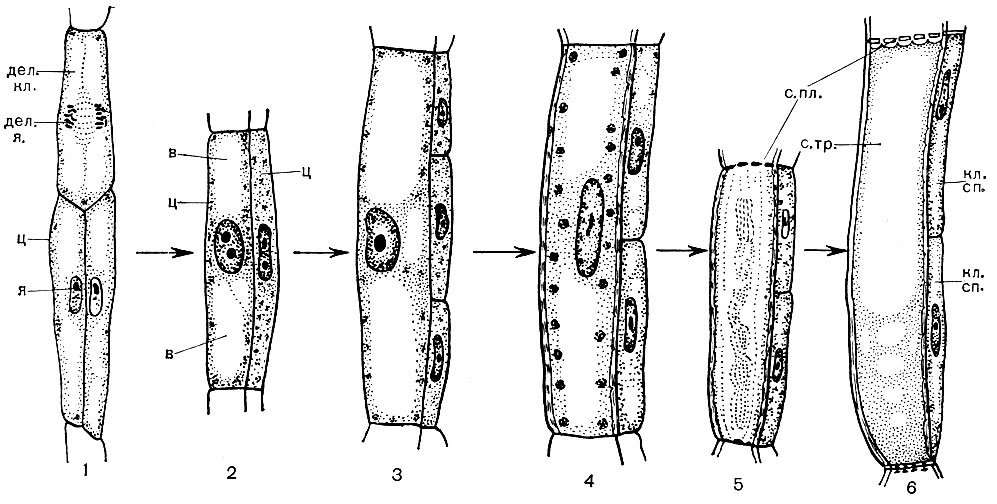
Most specialized cells are incapable of reproduction. However, the plant grows throughout its life, and new cells are formed in it throughout its life. They develop from the cells of educational - meristematic - tissues. Reproduction by division is a specialization of meristematic cells, their function in the body. The cells that have originated from them develop, turning into one or another specialized cells - cells of sieve tubes, cork cells, fibers, etc. Educational tissue is located in different parts of the plant (for example, at points of growth - at the tops of shoots, roots). There are usually several layers of educational tissue in the thickness of the stem. Under the cork layer there are phellogen cells, which, dividing, replenish the number of cork and crust cells. A layer of cambium cells is located inside the bast. Those of the cells they generate, which are formed outside the cambium, develop into bast cells (Fig. 25). Once inside, that is, closer to the axis of the trunk, give rise to wood cells.
Wood consists of elements of conductive, support and base fabrics. The first include wood fibers - long dead cells with lignified walls; to the second - vessels, which are the result of the fusion of many cells; to the third - cells of the woody parenchyma. Cells that differentiate from cambial elements into vascular cells (vessels) grow in length and width. Their side walls are thickened and lignified. In the walls, however, pores remain, closed only by a thin shell. The partitions at the junctions of adjacent cells disappear, the protoplast dies off. As a result, long vessels are formed, consisting of some walls. The conductive system extends through the root and stem into the leaf. Through such vessels, an ascending flow of water and salts dissolved in it is carried out from the roots to all organs of the plant.
In the center of the stem are core cells - round or polyhedral parenchymal cells. These are the elements of the main fabric. Sometimes they are hollow and contain air; sometimes they are filled with reserve nutrients, various crystals, tannins. Their walls can be lignified. Radial rays penetrate the wood and bast. Their cells are cambium derivatives and have a storage function. At the level of the bast, these rays expand noticeably.
In the leaf pulp, between the upper and lower epidermis, there is the main tissue - cells with thin membranes and a large number of green plastids - chloroplasts. Photosynthesis takes place in these cells. The upper layers are composed mostly of oblong cells, tightly adjacent to each other - this columnar parenchyma. The lower layers loosens between the cells there are intercellular spaces - spaces filled with air; it is a spongy parenchyma. The parenchyma is permeated with branching veins, which are built from bundles of mechanical (various fibers) and conductive (sieve tubes and aquifers) tissues.
![]()
This short and schematic presentation of the plan of the structure of the stem, leaf shows how diverse in size, shape, structure, functions of the cells of the same plant are. If you take different plants, then you can meet with an even greater variety of cells. However, as a rule, the cells of tissues of the same name, even in very distant plants, are similar, since these cells perform similar functions. The epidermal cells of a birch leaf and, for example, dandelion are more similar to each other than the epidermal cell of a dandelion and its sieve cell. The distinctive features of the cell are associated primarily with its special function. Cells specialized in mechanical function have thickened and often lignified membranes. Specialization in photosynthesis leads to the appearance of chloroplasts in cells. Specialization in the conducting function is associated with cell elongation, with the loss of the protoplast, with changes in the cell membranes at the junctions, due to which the cavities of adjacent cells communicate. Cells specialized in a protective function are characterized by a variety of changes in the outer walls, the presence of hairs, and the ability to produce protective substances.
Specialization in the function of storing nutrients can lead to an increase in the size of cells, to the appearance of very large vacuoles in them. In the cells of meristematic, intensively dividing, those intracellular structures are especially developed that provide the synthesis of substances - the constituent parts of the cell.
Depending on the nature of specialization, some features of the structure and work of the cell are very strongly developed, while others, on the contrary, are little developed or completely absent. As we have seen, some cells, having become specialized, die and it is in the dead state that they begin to perform their special function in a multicellular organism (cork cells, wood fibers, vessels).
The cells of multicellular filamentous algae and unicellular plants are very diverse and peculiar (Fig. 26 and 27). The cell of any of the latter, moreover, is very different from the cells of multicellular plants. She alone has to perform several functions, which in multicellular plants are divided between cells of different tissues.
At the same time, even very different cells have deep similarities in structure and function. For multicellular organisms, this is due, firstly, to the fact that all cells of the body (if we exclude vegetative reproduction, in which a whole multicellular complex is immediately separated from the mother plant as a new individual or its rudiment) are descendants of the same progenitor cell. Therefore, no matter how specialized the cells are, they have a common origin, and therefore are relatives.
Secondly, common features in the structure of cells of plants of different species are associated with the fact that all plants are related to one degree or another. All plants evolved by evolution from common single-celled ancestors. Common features are inherited by the cells of modern plants from ancient progenitor cells. This is also associated with the presence of common features of the structure and work of plant and animal cells.
Thirdly, the similarity is due to the fact that all living cells, no matter what special function they carry in the body, must first of all ensure their own life. The cells absorb nutrients, process them, gaining energy and building their own body, breathe, get rid of unnecessary substances, fight various damages, react to changes in external conditions, rebuilding their vital activity, and grow. All these processes in different cells are carried out in a similar way and with the help of structures of the same type, which are common in terms of the structure not only for different plant cells, but also for the cells of plants and animals. It must be said that the performance of any special function of a cell is based on its general properties. This or that trait, an ability inherent in all cells, develops especially strongly in a specialized cell and ensures that the cell performs its main, special function. Those common features, without which it is impossible to perform this special function, are preserved in the cell, and the rest can be lost. Dead specialized cells are an extreme case of this. The special function of such cells is related to their membrane; the protoplast is needed only as long as it creates a shell; after that, it dies off, and the entire cell consists only of an inanimate shell, which works for the needs of the plant.
Considering the general features of the structure and vital activity of plant cells, it is convenient to talk about a certain typical cell that has absorbed the common features of various specialized cells. Such a cell, although it does not exist in a "pure form", can even be depicted (Table 7, Fig. 28). Of real cells, they are more similar to such a "typical" cells of the leaf parenchyma (Table 8, Fig. 29).
Let's move on to describing the structure and work of a plant cell.
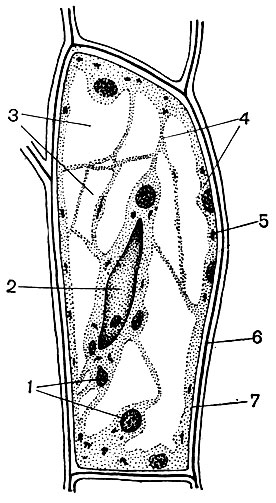
Table 5. Angiosperms: top left - magnolia flower; top right - apple flowers; below is a lotus.
Table 6. Angiosperms: above (from left to right) - bells, pelargonium, saffron (crocus); bottom left - asters; bottom right - burdock.
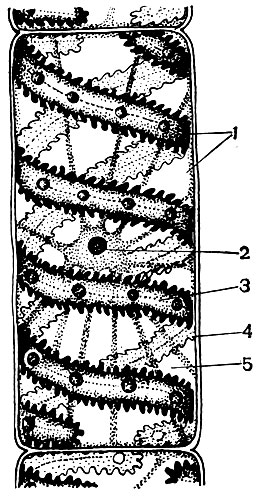
Outside, the plant cell is covered with a membrane that is not the same in thickness and structure in different cells. The substances forming it are produced in the cytoplasm and deposited outside of it, gradually creating a shell. These substances are primarily large-molecular polysaccharides - pectin, hemicellulose and cellulose in small amounts. They form the so-called primary shell. It is quite elastic, as the cell grows, it stretches and also grows, and therefore does not interfere with cell growth. However, it creates a certain strength of the cell and is able to protect it from mechanical damage. There are cells that lack such a primary membrane - these are some cells that serve for asexual and sexual reproduction (zoospores and gametes of algae and lower fungi, male gametes of higher plants). Many cells have not only a primary, but also a secondary membrane. It is formed under the primary and is built mainly from cellulose. Cellulose is a polysaccharide whose molecules form the finest filaments - microfibrils. In the shell, the cellulose filaments are immersed in an amorphous substance consisting of pectin compounds. In some cells, these cellulose microfibrils are located across the length of the cell, in rings; due to this, such cells can stretch in length but not in width (for example, cells of the vessels of the stem). Others have threads longitudinally; cells with such a membrane are elastic when stretched across, but very stiff in longitudinal stretching. In others, they are located obliquely, forming a spiral (epidermal hairs of cotton seeds, bast fibers). All this resembles reinforced concrete structures, with cellulose threads playing the role of iron rods here, and pectin substances playing the role of cement. Cells with a secondary membrane are very durable. They form the mechanical supporting tissues of the plant. Sometimes the secondary membrane also plays the role of a storehouse of nutritious products: the substances forming it can be converted into other, simpler ones, which are consumed as food.
In the envelope there are non-thickened places - pores (in the primary envelope they are called burrow fields), through which communication between neighboring cells is carried out. Thin cords of cytoplasm pass through the pore fields and pores. These are plasmodesmata that bind the cytoplasm of neighboring cells (Table 9). They exchange substances between neighboring cells. The plasmodesmata, along with the elements of the conducting tissue, connect the cells and tissues of the body into a single whole.
The exchange of substances and the spread of excitement allow cells to influence the development and work of each other, and each tissue affects the vital functions of all other tissues. This creates the coordination of the work of all parts of a single organism, the whole plant.
In many cells, the cell membrane with age is impregnated with substances that further strengthen it. The accumulation of lignin in it leads to lignification of the shell. The walls of wood cells are lignified and, often, bast fibers, cell shells of seed skin, sometimes pericarp (Fig. 30) and even petals. Cells with lignified walls are not only stronger, but also less permeable to microbes and water. The membranes of some other cells - integumentary tissues, as well as on the surfaces of wounds - are corked, soaked in suberin. The content of the cells then dies off, since suberin is impermeable to water and gases, but the tissue from such dead cells protects the inner living parts of the plant from harmful external influences. As already mentioned, the outer membranes of epidermal cells are usually impregnated or covered with cutin and wax, and this keeps the cells from excessive evaporation of water, from infection. The casing can also be impregnated with minerals, mainly calcium salts.
However, there are times in the life of cells when their shell should become less durable than it was before. This happens, for example, during seed germination.
The polysaccharide cell membrane is a characteristic feature of the structure of a plant cell, which distinguishes it from an animal cell.
The membrane, or cell wall, is a protective formation. The cytoplasm is located under the membrane. Its outermost layer, adjacent to the membrane, is the surface cell membrane - the plasmalemma. It is a combination of layers of fat-like and protein molecules. Such membranes are called lipoprotein ("lipos" - fat, "protein" - protein). A membrane of this design delimits the cytoplasm from the vacuoles; this membrane is called a tonoplast. Many cell organelles are built from lipoprotein membranes. However, in each case, the membrane is built of fat-like substances (lipids) and proteins inherent in this particular membrane. The qualitative diversity of lipids and especially proteins is colossal, hence the huge variety of membranes differing in properties, both within the same cell and in different cells.
Plasmalemma regulates the entry of substances into the cell and their exit from it, ensures the selective penetration of substances into and out of the cell. The rate of penetration through the membrane of different substances is different. Water and gaseous substances penetrate well through it. Fat-soluble substances also easily penetrate, probably due to the fact that it has a lipid layer. It is assumed that the lipid layer of the membrane is permeated with pores. This allows substances insoluble in fats to pass through the membrane. The pores carry an electric charge, so the penetration of ions through them is not entirely free. Under some conditions, the charge of the pores changes, and this regulates the permeability of the membranes for ions. However, the membrane is not equally permeable for different ions with the same charge, and for different uncharged molecules of close sizes. This is the manifestation of the most important property of the membrane - the selectivity of its permeability: for some molecules and ions it is better permeable, for others it is worse.
A substance moves into the cell, diffusing into it, if its concentration outside is greater than the concentration inside. Otherwise, it should diffuse out of the cell. But, having penetrated into the cell, the molecules of a given substance can immediately be involved in metabolism and undergo transformation into other substances. Then the concentration of this substance inside the cell drops again, and a new portion of its molecules diffuses into the cell. As a result, the cells that actively consume this or that substance, the latter intensively enters from the surrounding cells.
Another mechanism for enhanced absorption of a substance by a cell is its binding by proteins or other substances. By binding, the substance leaves the intracellular solution and does not interfere with further diffusion of the same substance. It continues to enter the cell and accumulates in it in a bound form. So, in the cells of some seaweed, iodine accumulates in concentrations a million times higher than its concentration in seawater.
In all these cases, we are talking about the movement of substances by diffusion. However, cell membranes also have mechanisms for moving substances through themselves from a lower concentration to a higher one. Among the proteins of the membrane, there are protein substances, the work of which is to move various substances from one side of the membrane to the other. They are called transport enzymes. Probably, such a transport enzyme forms an intermediate with the transported substance; the latter passes through the membrane, then breaks down into its original parts, after which the enzyme returns back, and the transferred substance remains on the other side of the membrane. The details of this process are unknown, but the very fact of such active transport of substances is beyond doubt. Unlike diffuse, passive, enzymatic transport requires energy expenditure. Probably, it is due to this active transport of ions that the cells of the root epidermis are able to absorb inorganic substances necessary for the plant from the soil and then transfer them through the plant from cell to cell.
With chemical or physical changes in the external and internal environment, cell membranes change their permeability, as well as the degree and the very nature of its selectivity. The mechanisms of regulation of the movement of substances into and out of the cell are based on this. A change in the permeability of membranes for nutrients is reflected in the intensity of metabolic processes in the cell, on the nature of the syntheses occurring in it, and throughout its life. The change in the membrane permeability for sodium and potassium ions is the mechanism for the propagation of cellular excitation - the emergence and movement of the biocurrent. A large number of cations, mainly potassium ions, are attached to the membrane surface. Therefore, it carries a positive charge from the outside. Under the influence of stimuli, potassium ions are cleaved from the affected area, the charge of the area drops and it becomes electronegative with respect to the neighboring areas of the membrane. This electronegativity is, in turn, an irritant for neighboring areas, in the same way removing their charge, depolarizing them. The wave of electronegativity spreads across the membrane - this is the biological current. Then the potassium ions again sit on the membrane, giving it the initial charge - a wave of depolarization is followed by a wave of restoration. The surfaces of the plasmodesmata passing through the cell walls and connecting the cytoplasm of neighboring cells are also formed by such membranes. Biocurrent moves along them, spreading from cell to cell. Plant cell biocurrents are still poorly understood. However, it is clear that they are a signaling method used in the plant cell to trigger some chemical reactions and inhibit others. The fact is that all the chemical processes occurring in the cell and constituting the essence of its vital activity are carried out with the participation of biological catalysts - enzymes. Each reaction is possible only when the enzyme that triggers it is active. Most enzymes become active under the action of certain inorganic cations: K., Na. , Ca .., Mg .., Mn ... Biocurrent, changing the membrane permeability for charged substances, creates conditions for their penetration through membranes and contact with enzymes. Thus, the biocurrent activates certain enzymes and thereby regulates and directs the metabolism in the cell. The regulation of metabolism by biocurrents is just one of the many ways to regulate intracellular metabolism.
What has been said about the permeability of the cell surface membrane - the plasmalemma - also applies to other intracellular membranes, including those of which many cell organelles are built.
The cytoplasm, once considered a homogeneous colloidal solution of protein substances, is actually complexly structured. With the development of microscopic research techniques, more and more subtle details of the structure of the cytoplasm became clear. In the cytoplasm, various organelles (organelles) were discovered - structures, each of which performs certain physiological and biochemical functions. The most important organelles of the cytoplasm are mitochondria, endoplasmic reticulum (endoplasmic reticulum), Golgi apparatus, ribosomes, plastids, lysosomes. Motile cells (zoospores and gametes of algae, spermatozoa of horsetails, ferns, cycads, some unicellular and colonial algae) have organelles of movement - flagella.
Especially a lot of new facts about the fine structure of the cytoplasm have brought and are bringing the use of an electron microscope, which makes it possible to study the details of the structure of the organelles themselves. Modern biophysical and biochemical methods make it possible to isolate certain organelles of the cytoplasm in pure form and then study them chemical composition and their functions. Outside the cell, in environments of complex composition, many organelles are able to do the work they do when they are in the cell.
The part of the cytoplasm in which the organelles are immersed and which so far appears to be structureless is called the main substance of the cytoplasm or hyaloplasm. Hyalo-plasma is by no means a passive filler, but an actively working part of the cytoplasm. A number of vital chemical processes take place in it, it contains many enzyme proteins, with the help of which these processes are carried out.
Cell life consists of continuous chemical work, which in its totality is called metabolism. In essence, the cell is a chemical plant that produces a wide range of products and independently extracts the energy needed to produce it. Her products are substances that she herself needs to maintain her own life (to build her body during growth and development, to replace her worn-out parts), and to create daughter cells during reproduction, and for the needs of other cells in the body.
All chemical reactions taking place in the cell can be divided into two groups. As a result, some of these or those substances break down into smaller molecules. As a result of other small-molecular substances, substances with larger molecules are synthesized. A molecule of any substance consists of atoms that are held together by chemical bonds, that is, by one or another amount of concentrated potential chemical energy. When the molecule is broken up, the bonds are broken and their chemical energy is released. For synthesis, that is, for the formation of a larger molecule from small ones, it is necessary to create new chemical bonds, It is necessary to put some portion of energy into them. On the contrary, decay reactions ultimately proceed with the release of energy, since during them the bonds between atoms are broken. The biological meaning of decay reactions going on in the cell is that they release chemical energy, which is then used by the cell for synthesis reactions and for the production of other types of work (electrical, mechanical, work on the transport of substances). Since the cell has been synthesizing various substances throughout its life, it has to continuously break down other substances. The released energy is not immediately used for syntheses. First, it is stored through the formation of special substances - accumulators of chemical energy - adenosine triphosphoric acid (ATP) and related compounds. At the right time and at the appropriate point of the cell, ATP is split and gives up energy for the synthesis of the substance necessary for the cell.
All chemical reactions in the cell - both synthesis and decay - are carried out with the help of enzymes. Enzymes are protein substances that accelerate the course of reactions. This acceleration is so great that without enzymes such reactions would not be possible at all in the cell. There are cases when, thanks to the enzyme, the reaction is accelerated by 1011 times. This means that a reaction that ends with the participation of the enzyme within 0.01 seconds, without it, would proceed 31 years. It is clear that such reactions without the enzyme would be simply unrealistic.
In addition, thanks to enzymes, the course of chemical reactions in the cell is controlled and regulated. The activity of enzymes in a cell changes in accordance with its need for a certain substance or energy, that is, as a result of the work of these very enzymes. When there is a need for a substance, those enzymes, thanks to which it is formed, are activated or synthesized again. The formation of substances, the release and storage of energy is the result of successive reactions, the result of the work of a whole chain of enzymes. All the structures of the living parts of the cell are built from enzymes and from substances that hold these enzymes together. In this case, the enzymes participating in adjacent, sequentially occurring reactions are located side by side. They transfer the molecules of the substances they convert to each other, as if along a conveyor, and each of them performs its own working operation on the molecule.
The hyaloplasm contains enzymes that break down glucose molecules into simpler pyruvic acid molecules. The energy released in this case is stored by the formation of ATP molecules. The same process takes place in the cell nucleus. However, the bulk of the energy is obtained in special organelles of the cytoplasm - mitochondria, since there is a deeper breakdown of substances.
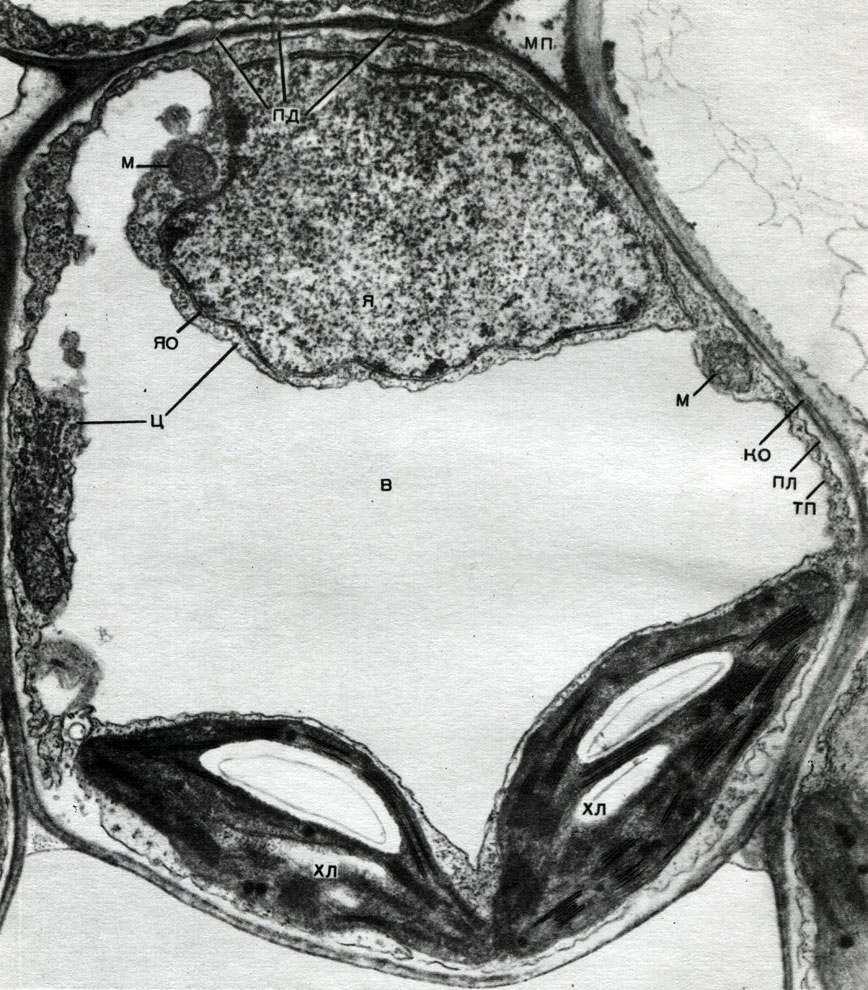
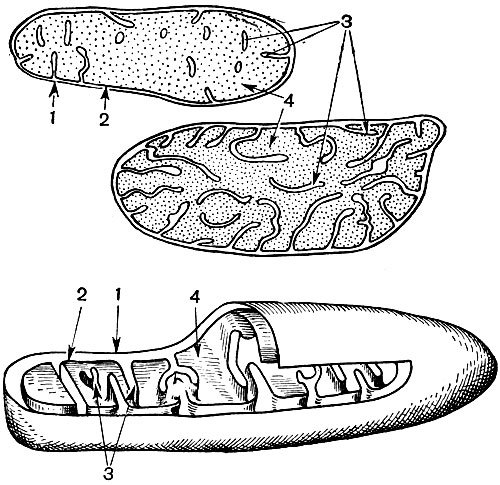
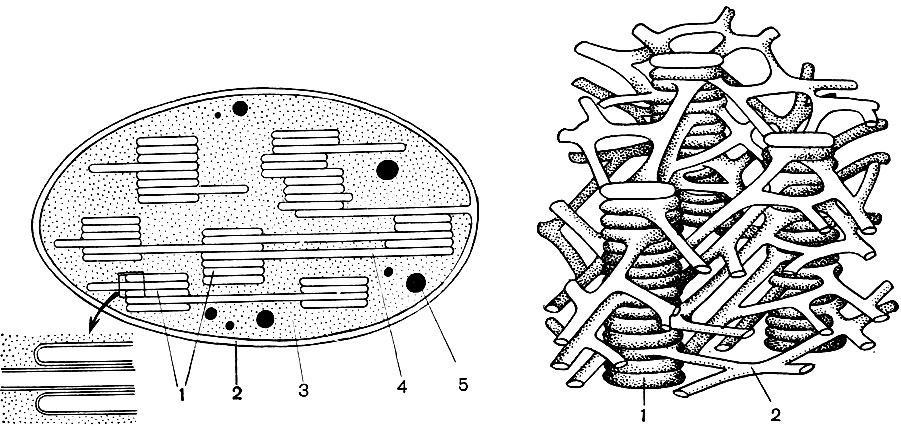
Mitochondria are small bodies of a rounded or oblong shape, 0.5 - 1.5 microns in size, that is, the size of a bacterium. Their number in a cell is usually large, about 100 - 3000. There are, however, cells with a small number of mitochondria. So, the sperm of the fucus seaweed contains only 4 mitochondria, and the unicellular micromonas alga contains only one. Mitochondria are visible under a light microscope, but their fine structure can be studied only with an electron microscope (Table 10, diagram of the structure - Fig. 31). Mitochondria are formations built from lipoprotein membranes immersed in a basic substance - a matrix. The mitochondrial membrane is formed by two membranes, between which there is a gap.
The inner membrane of the shell gives numerous invaginations inside, these are cristae. There is a matrix between them. Both the inner membrane of the mitochondrial membrane, and the cristae formed by it, are built from the ordering of located enzymes. Due to the folds - cristae, the working surface of the membranes inside the mitochondria is very large. A number of enzymes are found in the mitochondrial matrix, that is, between the cristae.
The combination of these enzymes carries out intracellular respiration and the storage of energy released during respiration in the form of ATP. The work of mitochondria is closely related to the processes taking place in the hyaloplasm, where the first stages of the breakdown of glucose and other substances to pyruvic acid take place. In mitochondria, its further splitting takes place. Pyruvic acid penetrates into mitochondria, and here, step by step, it is oxidized to carbon dioxide and water, and oxygen is consumed at the same time. This is intracellular respiration, in which the cell, splitting and oxidizing substances, produces a lot of energy, which it can then use for a variety of its needs.
The first stage of the breakdown of the glucose molecule, during which it is split in half and which takes place in the hyaloplasm, gives the cell only two ATP molecules.
As a result of the second stage, leading to the complete "combustion" of glucose, 36 more ATP molecules are formed. Therefore, mitochondria, in their function, are the power stations of the cell, machines for obtaining the bulk of energy. The very breakdown of glucose breakdown products occurs in the mitochondrial matrix, while ATP is formed due to reactions that take place on its inner membranes, which include respiratory enzymes and enzymes that ensure the formation of ATP. The number of cristae in mitochondria may vary. The more of them, the higher the biochemical activity of mitochondria.
We are talking here about glucose as a substance, breaking down which the cell produces energy. Glucose is central, but not the only such substance. Its molecule has a skeleton of six carbon atoms connected to each other. As a result of a long and complex chain of reactions, its molecule is crushed, oxidized and, in the end, split into six molecules of an inorganic substance - carbon dioxide (CO 2), each molecule of which contains only one carbon atom, and it is extremely oxidized. By adding up all the successive reactions of glucose oxidation and excluding all intermediate products, you can get the total reaction of this process:
Starch is easily converted to glucose, after which it undergoes the above-described degradation. Proteins and fats give various organic acids, which are converted into intermediate products of the breakdown of glucose and then oxidized in the same way as the latter, and with the help of the same enzymes.
Complete biological oxidation of organic matter is similar to its combustion. In both cases, the result is carbon dioxide, water and energy released. However, during combustion, this energy is released in the form of heat, and completely at once; during biological oxidation, the energy of chemical bonds is released in portions, and its main part is bound, passing into the energy of the phosphate chemical bond of ATP. As a result, the cell receives a concentrate of energy in such a form, which then at the right time and at the appropriate point can be used to create new chemical bonds, to synthesize new substances, as well as to perform other types of work - electrical, mechanical, as well as work on the transport of substances from environment to cell, from cell to environment and from cell to cell.
The more active the vital activity of the cell, the greater its need for energy and the more mitochondria it contains. They can be unevenly distributed within the same cell: there are more of them in the part of the cell that is currently working more actively.
Mitochondria are able to synthesize some of the substances of which they themselves are composed. Thanks to this, mitochondria can multiply.
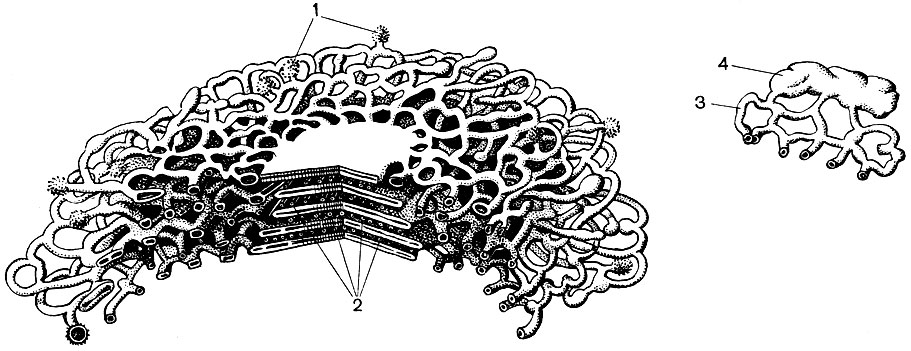
The endoplasmic reticulum is an organoid of the cytoplasm, in which a great many substances are synthesized (Table 10). The endoplasmic reticulum is a system of channels that penetrate the cytoplasm and which narrow in some areas, expand in others, forming either cisterns, or flat bags, or branching tubes. The walls of all these formations are built of membranes that include enzymes.
As in other membrane formations of the cell, enzymes in the reticulum are arranged in an orderly manner. In this case, neighboring enzymes carry out sequentially proceeding reactions (working operations), and their group carries out the entire chain of reactions leading to the creation of this or that substance.
Distinguish between agranular (smooth) and granular endoplasmic reticulum. On the outer surface of the channels of the granular reticulum, there are numerous small organelles - ribosomes, whose function is the synthesis of protein molecules. The agranular endoplasmic reticulum, which quantitatively prevails over the granular one in the plant cell, does not carry ribosomes.
The endoplasmic reticulum, in addition to being a conveyor for many types of enzymatic conversion of substances, mainly for their synthesis, is also a system of highways along which substances move through the cell. The reticulum begins from the outer membrane of the membrane of the nucleus and, branching, approaches various organelles of the cytoplasm, as well as the plasmalemma. Thus, he connects all parts of the cell with each other. In addition, the channels of the endoplasmic reticulum pass through the plasmodesmata, connecting the reticulum of neighboring cells.
Further, the membranes of the endoplasmic reticulum dissect the cytoplasm into numerous compartments, due to which the cell cannot be represented as a homogeneous mass in which a variety of substances are mixed. Conditions in one compartment may be very different from any other; the processes taking place in it can only take place in it, while in each other other processes take place.
Finally, the membranes of the endoplasmic reticulum are those surfaces along which biocurrents propagate, which are signals that change the selective permeability of membranes and thereby the activity of enzymes. Thanks to this, some chemical reactions are set in motion, others are inhibited - the metabolism obeys regulation and proceeds in a coordinated manner.
Many of the substances synthesized in the cell must be concentrated and released from the cell either into the external environment or into the intracellular vacuole. In addition, a cell also concentrates substances that enter it from other cells, for example, if it stores them in reserve. This work is done by dictyosomes. Usually in a plant cell there are several dictyosomes, and their whole set is called the Golge apparatus (or complex) and the given cell. Each dictyosome is a system of stacked membranes (Table 10 - section, Fig. 32 - volumetric diagram). The cavities between the membranes that form this organoid look like narrow slits, then flat sacs - cisterns, then bubbles. Their shape changes during the operation of the organoid and, apparently, depends on the degree of filling of the intermembrane spaces with the secreted and accumulated substances. The formed and overgrown vesicles are separated from the organoid. Apparently, many cellular vacuoles, surrounded by membranes - tonoplast, are the product of the activity of the Golgi apparatus; these are bubbles detached from it and then enlarged.
The Golgi apparatus is especially developed in excretory (secretory) cells, in which various substances are deposited or from which various substances are excreted. It synthesizes and releases substances that form the cell membrane.
Lysosomes are rather small (about 0.5 microns in diameter) rounded bodies - another organoid of the cytoplasm. They are covered with a membrane - a lipoprotein membrane. The contents of lysosomes are enzymes that digest proteins, carbohydrates, nucleic acids and lipids. The lysosome membrane prevents the release of enzymes from the organoid into the hyaloplasm, otherwise the latter would be digested by these enzymes.
It can be thought that lysosomes are products of the activity of the Golgi apparatus, bubbles detached from it, in which this organoid has accumulated digesting enzymes.
Those parts of the cell that die off during its development are destroyed by lysosomal enzymes. In a deceased cell, lysosomes are destroyed, enzymes are found in the cytoplasm, and the entire cell, with the exception of the membrane, undergoes digestion.
Ribosomes are very small organelles, about 250A in diameter. They are almost spherical in shape. Some of them are attached to the outer (hyaloplasmic) surfaces of the membranes that form the channels of the granular endoplasmic reticulum; some are in a free state in the hyaloplasm. A cell can contain up to 5 million ribosomes. They are "apparatus for protein synthesis. Therefore, there are especially many of them in cells that actively form protein - in growing cells, in cells that secrete protein substances. Ribosomes are also found in mitochondria and chloroplasts, where they synthesize some of the proteins from which these organelles.
Organelles called microtubules are found in many cells. Their very name speaks of their shape - these are tubes with a channel inside. Their outer diameter is about 250A. Sometimes these are double tubes - two single ones, lying side by side with each other and having a common wall that separates their cavities. The walls of microtubules are built from protein molecules. It is believed that microtubules are associated with the contractile (motor) activity of the cytoplasm and its formations. From them, as from building parts, the contractile structures of the flagellum, an organoid, with the help of which some unicellular and colonial algae move, as well as cells that serve for the reproduction of many lower plants, are apparently built. During cell division, spindle threads are formed from microtubules, which will be discussed below. During the period of division, microtubules gather in groups and form these filaments. At the end of division, the filaments again disintegrate into separate microtubules. In cells or parts of them that lack a dense membrane, microtubules possibly perform a supporting function, making up the inner skeleton of the cell.
Plastids are organelles inherent only in plant cells. Usually these are large bodies, clearly visible under a light microscope.
There are 3 types of plastids: colorless - leukoplasts, green - chloroplasts, colored in other colors - chromoplasts. Each type of plastids have their own structure and carry their own inherent functions. However, transitions of plastids from one type to another are possible. Thus, the greening of potato tubers is caused by the rearrangement of their leukoplasts into chloroplasts. In the carrot root crop, the leukoplasts pass into chromoplasts. All three types of plastids are formed from proplastids.
Proplastids are colorless bodies, similar to mitochondria, but somewhat larger than them. They are found in large quantities in meristematic cells. Leukoplasts are found in the cells of uncolored parts of plants (fruits, seeds, roots, leaf epidermis). Their form is indefinite. Most often, there are leukoplasts in which starch is deposited (it is formed from sugars). There are leukoplasts that store proteins. The least common are fat-filled leukoplasts; they are formed during the aging of chloroplasts. There are no significant differences between leukoplasts and proplastids.
Chloroplasts are plastids of higher plants in which the process of photosynthesis is taking place, that is, the use of the energy of light rays to form organic substances from inorganic substances (carbon dioxide and water) with the simultaneous release of oxygen into the atmosphere Chloroplasts have the form of a biconvex lens, their size is about 4 - 6 mk. They are found in the narenchymal cells of leaves and other green parts of higher plants. Their number in a cell varies from 25 to 30.
The structure of the chloroplast is given in Tables 11 and 12 (sectional view) and the diagram in Figure 33. Outside, the chloroplast is covered with a membrane consisting of two lipoprotein membranes. Under it, in the main substance (stroma), numerous formations - lamellae - are ordered. They form flat pouches that lie on top of each other in regular stands. These stoics, which resemble coins stacked in a column, are called grains. Longer lamellae pass through them, so that all the chloroplast grains are connected into a single system. The composition of the membranes forming the granules includes a green pigment - chlorophyll. It is here that the light reactions of photosynthesis take place - the absorption of light rays by chlorophyll and the conversion of light energy into the energy of excited electrons. Electrons excited by light, that is, having excess energy, give up energy to decompose water and synthesize ATP. When water decomposes, oxygen and hydrogen are formed. Oxygen is released into the atmosphere, and hydrogen is bound by the protein ferredoxin. Ferredoxin is then re-oxidized, releasing this hydrogen to a reducing agent, abbreviated as NADP. NADP goes into the reduced form - NADP-H 2. Thus, the result of light reactions of photosynthesis is the formation of ATP, NADP-H 2 and oxygen, and water and light energy are consumed.
ATP accumulates a lot of energy - it is then used for syntheses, as well as for other needs of the cell. NADP-H 2 is a hydrogen accumulator, and it then gives it away easily. Therefore, NADP-H 2 is a chemical reducing agent. A large number of biosyntheses are associated with reduction, and NADPH 2 acts as a supplier of hydrogen in these reactions.
Further, with the help of chloroplast stromal enzymes, that is, outside the granules, dark reactions occur: hydrogen and the energy contained in ATP are used to reduce atmospheric carbon dioxide (CO2) and include it in the composition of organic substances. The first organic matter, formed as a result of photosynthesis, undergoes a large number of rearrangements and gives rise to the whole variety of organic substances synthesized in the plant and making up its body. A number of these transformations take place right there, in the chloroplast stroma, where there are enzymes for the formation of sugars, fats, as well as everything necessary for protein synthesis. The sugars can then either move from the chloroplast to other cell structures, and from there to other plant cells, or form starch, the grains of which can often be seen in chloroplasts. Fats are also deposited in chloroplasts either in the form of drops, or in the form of simpler substances, precursors of fats, are released from the chloroplast.
The complication of substances is associated with the creation of new chemical bonds and usually requires energy. Its source is all the same photosynthesis. The fact is that a significant proportion of the substances formed as a result of photosynthesis again decomposes in the hyaloplasm and mitochondria (in the case of complete combustion, to substances that serve as the starting material for photosynthesis - C0 2 and H 2 0). As a result of this process, which is essentially the reverse of photosynthesis, the energy previously accumulated in the chemical bonds of decomposed substances is released and - again through the aid of ATP - is spent on the formation of new chemical bonds of the synthesized molecules. Thus, a significant part of the production of photosynthesis is needed only in order to bind the energy of light and, having converted it into chemical, use it for the synthesis of completely different substances.
And only part of the organic matter formed during photosynthesis is used as a building material for these syntheses.
The production of photosynthesis (biomass) is colossal. For a year on the globe, it is about 1010 tons. Organic substances created by plants are the only source of life not only for plants, but also for animals, since the latter process ready-made organic substances, feeding either directly on plants or other animals that, in turn, they feed on plants. Thus, photosynthesis is at the heart of all modern life on Earth. All transformations of substances and energy in plants and animals are rearrangements, recombinations and transfers of matter and energy of the primary products of photosynthesis. Photosynthesis is important for all living things and the fact that one of its products is free oxygen, originating from a water molecule and released into the atmosphere. It is believed that all the oxygen in the atmosphere comes from photosynthesis. It is essential for respiration for both plants and animals.
Chloroplasts are able to move around the cell. In low light, they are located under the cell wall that faces the light. In doing so, they turn to light with their larger surface. If the light is too intense, they turn to face it and line up along the shades parallel to the light rays. At medium illumination, chloroplasts occupy a position intermediate between the two extreme ones. In any case, one result is achieved: chloroplasts are in the most favorable lighting conditions for photosynthesis. Such movement of chloroplasts (phototaxis) is a manifestation of one of the types of irritability in plants.
Chloroplasts have a certain autonomy in the cell system. They have their own ribosomes and a set of substances that determine the synthesis of a number of chloroplast's own proteins. There are also enzymes, the work of which leads to the formation of lipids that make up the lamellae, and chlorophyll. As we have seen, chloroplast also has an autonomous energy production system. Thanks to all this, chloroplasts are able to independently build their own structures. There is even a view that chloroplasts (like mitochondria) originated from some lower organisms that settled in a plant cell and first entered into symbiosis with it, and then became its integral part, an organoid.
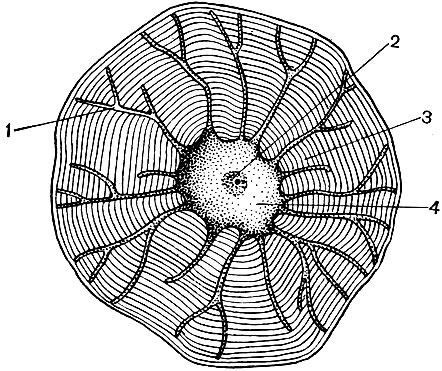
In lower plants, photosynthesis is also carried out by specialized, although not as highly developed as in the chloroplast, membrane structures. In photosynthetic bacteria, membranes containing chlorophyll form a network that permeates the bacterial body. In blue-green algae, photosynthetic membranes are fused into flat bubbles. In green and other algae, the system of these membranes is separated from the rest of the cell by a covering membrane and forms a special organoid - a chromatophore. The number of chromatophores in a cell is small; often a cell contains only one chromatophore. Their shape is very different in algae of different species.
In Spirogyra, the chromatophore has the form of a ribbon spiraling along the cell walls; in Clostridium, these are ribbed cylinders; the zygnema has stellate bodies.
Chromoplasts arise either from proplastids, or from chloroplasts, or from leukoplasts. Their inner membrane structure is much simpler than that of chloroplasts. Gran is not, the stroma contains a lot of yellow or orange pigment. Chromoplasts are found in the cells of petals, fruits, and root crops.
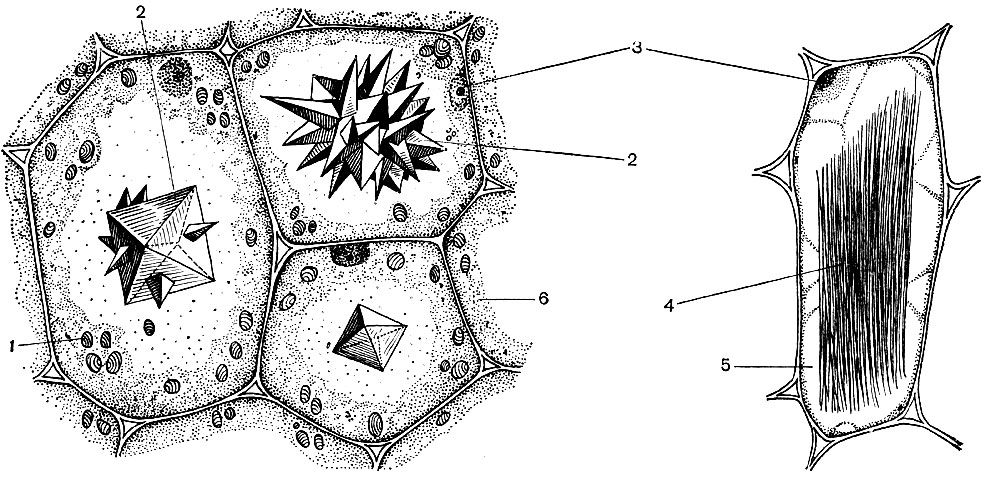
A typical plant cell has a large vacuole filled with liquid contents. Often the vacuole occupies almost the entire volume of the cell, so that the cytoplasm is only a thin layer adjacent to the cell membrane. Young cells have several small vacuoles, which, as the cells develop, grow and merge into one. The content of the vacuole - cell sap - is an aqueous solution of very many substances: sugars, amino acids, other organic acids, pigments (dyes), vitamins, tannins, alkaloids, glycosides, inorganic salts (nitrates, phosphates, chlorides), and sometimes proteins.
All these substances are the waste products of the cell. Some of them are stored in vacuolar (cell) sap as reserve substances and, over time, re-enter the cytoplasm for use. Others are metabolic waste products removed from the cytoplasm. So, oxalic acid is removed into the vacuole; crystals of calcium oxalate are often deposited in vacuolar sap - sometimes in the form of single crystals, in other cases in the form of a conglomerate of crystals of this salt - multifaceted (druses) or acicular (raffids) - shown in Figure 34.
The vast majority of plants (with the exception of prokaryotic organisms) each living cell has a nucleus or several nuclei. A cell devoid of a nucleus can only live for a short time. Anucleated cells of sieve tubes are living cells. But they do not live long. In all other cases, the non-nuclear cells are dead.

The nucleus always lies in the cytoplasm. The shape of the nucleus can be different - round, oval, strongly elongated, irregularly multi-lobed. In some cells, the contours of the nucleus change during its functioning, and lobes of various sizes are formed on its surface. The sizes of the nuclei are not the same in the cells of different plants, and in different cells of the same plant. Relatively large nuclei are found in young, meristematic cells, in which they can occupy up to 3/4 of the volume of the entire cell. The relative and sometimes absolute sizes of the nuclei in developed cells are much smaller than in young ones.
Outside, the core is covered with a shell consisting of two membranes, between which there is a gap - the perinuclear space. The shell is interrupted by pores. The outer of the two membranes of the envelope gives out outgrowths that directly pass into the walls of the endoplasmic reticulum of the cytoplasm. Both the pores and the direct connection of the endoplasmic reticulum with the perinuclear space provide close contact between the nucleus and the cytoplasm.
The content of the nucleus is a granular basic substance (nuclear juice, or nucleoplasm), in which denser structures are placed - chromosomes and the nucleolus. The nucleolus is the apparatus for synthesizing the material of ribosomes and the place of their assembly from this material.
Chromosomes are built from a large number of deoxyribonucleic acid (DNA) molecules linked to histone protein molecules. DNA molecules are long, complexly packed double strands. Each molecule consists of two strands, twisted spirally around one another. A thread, in turn, is a chain of a huge number of so-called nucleotides. Nucleotide is a compound of nitrogenous base, carbohydrate (deoxyribose) and phosphoric acid.
Each of the nucleotides contains one of the following four nitrogenous bases: adenine, guanine, cytosine, or thymine. Accordingly, 4 different nucleotides are distinguished in DNA: adenine (A), guanine (G), cytosine (C) and thymine (T). The nucleotides are linked together through their phosphate groups to form a long chain. Two chains, twisted around each other and forming one DNA molecule, are bonded together by chemical interaction (the so-called hydrogen bonds) of the nitrogenous bases of their nucleotides. The bases form pairs - one base from one chain, the other from the second. The diagram in Figure 35 gives an idea of \u200b\u200bthe order in which the nucleotides are linked into a chain, and the two chains - with each other.
Although all deoxyribonucleic acid molecules are built according to the described single plan, their specific qualitative composition is different; they also differ in the size of the molecules. A DNA molecule contains up to 50 - 100 thousand base pairs, but the number of pairs in each molecule is different from the number of pairs of nucleotides in any other molecule. In addition, there are 4 different types of nucleotides, and each DNA molecule has its own inherent percentage of nucleotides of each given type. In other words, the quantitative ratio A: G: C: T for each DNA molecule is different. And finally, in each DNA molecule, the order of alternation of nucleotides A, G, C, T is characteristic only for this molecule. Permutation of only two pairs of 50 thousand base pairs, even if adjacent, dramatically changes the properties of the entire molecule; the same, replacing at least one pair, say From - G to A - T or G - Iz; the same - the absence of one pair from the same 50 thousand. In reality, DNA molecules may differ from each other not in one link, not in one or two pairs of nucleotides, but in a huge number of them at once. The number of possible permutations of nucleotide pairs in a DNA molecule is infinite, and, accordingly, the number of different molecules is infinite, each of which has its own properties. The sequence of nucleotides in a DNA molecule is an encrypted record of the composition of a particular protein characteristic of a given cell. Throughout each DNA molecule, records of the composition of several proteins are sequentially fit, and in all DNA molecules of all chromosomes of the nucleus - records of the composition of all proteins of a cell that can be synthesized in it during its life. The essence of these coded records is as follows.
The molecules of each protein are chains of amino acids connected in series. There are about 20 different amino acids, and the nature of a protein is determined by which amino acids its molecule consists of, what is their total amount in the molecule and in what sequence they are connected to each other. For example, a DNA segment responsible for the composition of a certain protein (each such segment is called a gene) is a record of a specific sequence of amino acids that form a molecule of a given protein, their total number in it. Every 3 consecutive nucleotides of the DNA chain designate (encode) one amino acid of the corresponding protein molecule. The next 3 nucleotides encode the next amino acid, and so on. The number of possible differing combinations of 3 nucleotides from among four different types is more than enough to encode 20 amino acids. Interestingly, the same amino acids throughout the living world are encoded by the same nucleotide combinations. As a result, in one gene, its nucleotide composition encodes the amino acid composition of all proteins that can be synthesized in a given cell.
Cellular proteins are enzymes. Enzymes determine the course of all reactions that make up the essence of the life of cells. The formation and transformation of all other substances of the cell depends on the presence of certain enzymes, be it fats, carbohydrates, alkaloids, resins, etc. This, in turn, forms all the properties of the cell that distinguish it from the cells of other plants in the cell nuclei which encoded the composition of another set of enzymes. That is why information about the composition of all proteins that can be formed in a cell is information about all the properties of a cell and an organism. Further we will see that this information is hereditary, that is, that it is fully transmitted from cell to cell during their reproduction and from the mother plant to the daughter plants.
This information is stored in chromosomes. However, the realization of this information - the synthesis of proteins - does not take place in them. Chromosome genes act only as initiators of this synthesis. When a cell needs to form a particular protein, the gene in which the composition of this protein is encoded is activated. This means that molecules of the so-called informational ribonucleic acid (i-RNA) are formed on the section of the DNA strand that makes up this gene.
The structure and composition of these molecules, which are single chains of nucleotides, reflect the nucleotide structure of the gene on which they were formed. Thus, the information about the composition of the future protein is copied.
The formation of m-RNA molecules (a lot of them are formed on each gene) also means the multiplication of information, as it were, the imprinting of many identical matrices that reflect the structure of the same gene and thereby carry information about the amino acid composition of a given protein in the nucleotide sequence of their molecules. These gene prints pass from the nucleus to the cytoplasm. Here, the information contained in the matrices - i-RNA molecules, is deciphered, the information is realized, it is translated from the language of the nucleotide sequence into the language of the amino acid sequence, i.e., the synthesis of protein molecules of a given composition.
In the decoding and synthesis, in addition to the i-RNA molecule, a large number of molecules of various transport ribonucleic acids (t-RNA), ribosomes and a number of enzymes are involved. Amino acids bind to t-RNA - molecule by molecule. Each of the 20 types of amino acids has its own t-RNA. So, the t-RNA molecule has chemical groups that can recognize its amino acid, choosing it from all available amino acids. This happens with the help of special enzymes. Having recognized its amino acid, t-RNA enters into a connection with it. The ribosome is attached to the beginning of the i-RNA molecule, which, moving along the i-RNA, connects to each other in a polypeptide chain exactly those amino acids whose order is encrypted by the nucleotide sequence of the given i-RNA. This reading of information, this decoding of it occurs thanks to a special mechanism in the transport RNA, the molecules of which line up exactly those amino acids that are "designated", "written" by the nucleotide sequence of the i-RNA molecule one after another. The ribosome is an apparatus for chemical binding of amino acids into a protein molecule.
So, the general scheme is as follows. On the activated gene - a section of one of the chains of a DNA molecule - nucleotide chains, i-RNA molecules, are synthesized, the composition of which accurately reflects the composition of the gene, and therefore carries a record of the composition of the protein encoded by this gene. In the cytoplasm of the ribosome, on the basis of information transferred from the gene by the m-RNA molecules, using transport RNAs, different amino acids are connected in a given sequence, resulting in the formation of a molecule of the protein whose composition is encoded in this gene.
The formation of i-RNA molecules on the activated gene serves as both a command and a specific program for the synthesis of a strictly defined protein. The same mRNA molecule is used as a blueprint to create many of the same protein molecules. However, it is rather short-lived, therefore, for a long-term synthesis of new molecules of the same protein, the formation of identical, but new copies of m-RNA molecules on the same gene is necessary. With the transition of the gene to an inactive state - and this happens when the cell's need for this protein disappears - it is blocked, stops forming i-RNA, and soon the synthesis of this protein stops. During the life of a cell, it needs various proteins. Each time, the genes that define them are activated and the corresponding composition of i-RNA molecules is formed.
Each ribosome produces many molecules of different proteins during its life. It can work on the basis of any i-RNA, and the result, the nature of the protein created by it, depends only on the composition of the i-RNA in contact with which the ribosome was working this time.
Thus, the cell nucleus performs the following interrelated functions. It stores information about the composition of all proteins that can be synthesized in a given cell during its life. (The exception is some proteins of mitochondria and chloroplasts. Their composition is encoded in their own DNA of these organelles, where these DNAs are located. Here, on the spot, all stages of decoding the information contained in these DNAs take place, including the synthesis of these proteins using own ribosomes.)
The nucleus stores information about all the properties of the cell and the organism. The nucleus organizes the synthesis of each of these proteins at the right time. During cell division, accompanied by nuclear division, all this information is transferred in full to each of the newly formed nuclei, to each new cell. This is possible due to the fact that before division, all the gene material of the chromosomes is self-doubling, two identical copies are formed, and one of them ends up in each of the new nuclei. The nucleus of each cell contains a complete set of genes characteristic of a given organism. However, during the life of various specialized cells, not all genes work. In some cells, one part of the genes functions, in others - another, in the third - a third. That is why the cells of different tissues of one organism differ from each other. A significant part of genes remains in a passive, only "storing information" state from birth to the death of a cell or organism. In addition, genes in a given cell do not work simultaneously: some are active in one period of the cell's life, at a certain stage of its development; others - on the other, etc.
Why some genes remain inactive, how others turn on and off is a very complex and important problem that is now being intensively investigated. Proteins-histones seem to play an important role in blocking and unblocking genes. They are part of chromosomes, being in conjunction with DNA. Perhaps the activation, "liberation" of a gene occurs when a histone molecule is detached from the corresponding region of DNA, thereby exposing its chains, allowing them to unravel and begin to function chemically. The attachment of histone leads to the blockage of the gene. However, how histone attachment and detachment is controlled is not clear enough.
The gene material, as said, is found in the chromosomes. Therefore, the work of the nucleus in storing hereditary information, in doubling it and transferring it from cell to cell, in organizing the synthesis of various proteins during the life of a cell is primarily the work of chromosomes. Although they exist throughout the life of a cell, they are distinguished in the nucleus in the form of clear structures only during cell division. At this time, the condensation of the material of the chromosomes and the chromosome can be identified by microscopy methods as individual, clearly defined formations.
Chromosomes have different shapes. These are either straight or curved sticks, or various hooks, oval bodies, balls (Fig. 36). They vary greatly in size. Each plant cell of a given species contains in its nucleus the same set (or sets) of chromosomes from a strictly defined number of different, but strictly defined chromosomes. In all higher and some lower plants, two generations alternate during their life cycle: with cells containing a single (haploid, n) set of chromosomes in their nuclei, and with cells whose nuclei have a double (diploid, 2 n) set of chromosomes. A haploid set consists of n different chromosomes, one of each type. The numerical value is strictly constant for all haploid cells of all plants of a given species. So, for wild strawberries this number is 7, for peas - also 7, but for beans - 11, for an apple tree - 17, for an earthen pear - 51, for sugar cane - 60. A diploid set is two haploid ones folded together. It contains two chromosomes of each type. If a pea has 7 different chromosomes in a haploid set, then a diploid set has 7 different pairs of chromosomes, and two chromosomes within each pair are the same. It is easy to calculate that for the listed plants the diploid set (2n) is 14 for strawberries, 14 for peas, 22 for beans, 34 for apples, 102 for an earthen pear, and 120 chromosomes for sugar cane.
In the life cycle of mosses and ferns, organisms alternate with diploid and haploid cells. In ferns, these organisms lead an independent, isolated life. In mosses, a diploid plant lives on a haploid plant. In a diploid organism, haploid cells are formed - spores. Each 1 of them, germinating, gives a haploid organism, that is, an organism built from haploid cells. In it, haploid germ cells are formed - gametes.
At the moment of fertilization, the male and female gametes merge, forming one cell - a zygote, which has one nucleus - the result of the fusion of the nuclei of both gametes. From each of the gametes, the nucleus of the zygote receives a haploid set of chromosomes, and as a result it has a double, diploid set of them. An organism develops from a zygote, each cell of which has a diploid set of chromosomes.

In a diploid set, two chromosomes of each pair are identical in shape, internal structure, and contain genes that control the appearance of homogeneous traits (Fig. 36). They are called homologous chromosomes. One of them comes from the haploid set of the paternal gamete, the other from the maternal one. Therefore, in dioecious organisms, one of them carries genes that determine the development of the characteristics under its jurisdiction according to the paternal type, the second - according to the maternal type.
Homologous chromosomes of another pair in the same way determine the development of another series of characters, the third pair-third row, etc. Although the paternal and maternal organisms belong to the same species, their hereditary properties are not identical - they also have individual differences. Therefore, some genes of one homologous chromosome are not identical to the corresponding genes of the second.
The haploid set, which is part of the diploid and originating from the paternal gamete, carries paternal heredity with its individual features, and the haploid set from the maternal gamete carries the maternal one. The complex interaction of homogeneous, but not always identical genes of two haploid sets, which together form one diploid, determines which traits will appear in the diploid offspring, which, in essence, is a hybrid of the father and mother.
In gymnosperms and angiosperms, the alternation of generations occurs, in principle, as in mosses and ferns, but their haploid phase is greatly reduced and is often represented only by a group of cells. She does not live independently, but in the body of a haploid plant. In angiosperms, the female haploid generation is enclosed in the embryo sac located in the ovule, and the male generation is inside the pollen grain.
Even before cell division, each DNA molecule in each chromosome attaches itself a copy of itself - a second similar molecule. As a result, the entire hereditary material of the cell doubles, and each chromosome now consists of two equal parts - chromatids. Further, the "task" of the cell is to divide each chromosome into chromatids and distribute them strictly equally between the future daughter cells: one chromatid from each chromosome must be sent to each of them. This is done as follows. Immediately before cell division, the chromosomes are strongly condensed and contracted. Then they are located in one plane - along the equator of the nucleus, with one chromatid of each of them facing one pole of the cell, the other - to the opposite. The nuclear envelope disappears, and the nucleolus also dissolves. Filaments appear between the poles of the cell, which together form a spindle shape. The filaments are collected from microtubules connecting to each other. The spindle consists of two kinds of threads. Some are continuous cells going from one pole to the other. Others are pulling, each of which connects a pole to one of the chromatids. In the chromosome there is a site - the kinetochore, to which the pulling threads are attached - one from one pole, the other from the other. Further, the chromatids diverge. Chromatids, which previously constituted each chromosome, are separated from each other and, pulled up by threads, diverge to opposite poles of the cell. As a result, one complete set of chromatids is collected at each pole, which are now chromosomes. After that, the threads of the spindle disintegrate, a nuclear envelope is formed around each set of chromosomes, the chromosomes greatly swell (despiralize). A nucleolus appears in each nucleus. The nucleus acquires the structure characteristic of the nuclei of non-dividing cells. In the median plane of the cell, a septum forms, dividing the cell into two daughter cells.
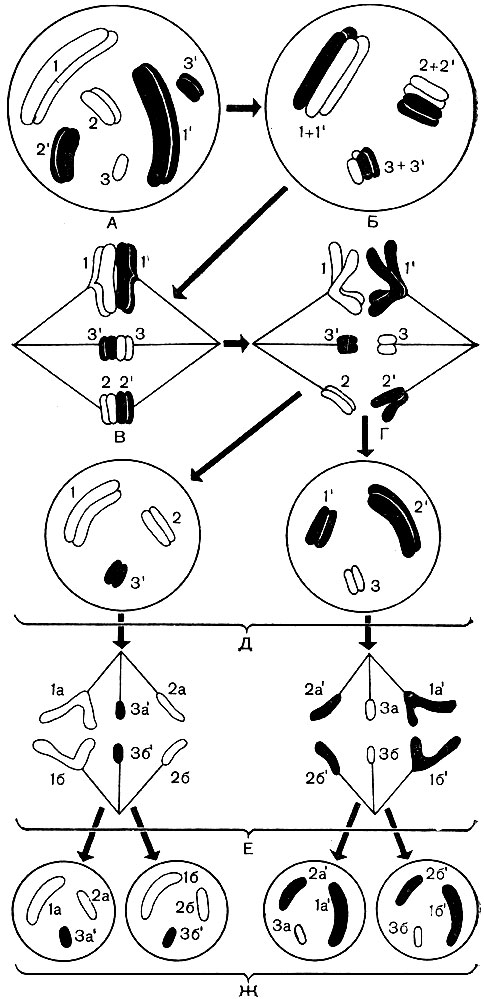
Figure: 38. Scheme of the behavior of the chromosomes of a "conditional" cell during meiosis. A - depicted is a diploid cell nucleus containing 3 pairs of chromosomes (1 and 1 "- the 1st pair of homologous chromosomes, 2 and 2" - the 2nd pair, 3 and 3 "- the 3rd pair). 2n \u003d 6.3 chromosomes originating from the maternal organism are light (numbers without a stroke); their paired homologous chromosomes originating from the paternal organism are blackened (numbers with a stroke). Homologous chromosomes are designated by a common number. Each chromosome consists of two chromatids. B - fusion of homologous chromosomes At this stage, homologous chromosomes exchange separate sections with each other. As a result, some redistribution of the maternal and paternal hereditary material between the chromosomes (crossing over) occurs. C - spindle threads are formed that attach to chromosomes, the nuclear membrane disappears. D - homologous chromosomes diverge to opposite poles cells, at the poles there is one homologous chromosome from each pair, the total number of chromosomes at each pole is half that in the original nucleus A. D - image there are two nuclei with a haploid set of chromosomes in each; one of them got more grandmother's (light, maternal in relation to the nucleus A) and less grandfather's (blackened, paternal in relation to the nucleus A) chromosomes, in the other - on the contrary; thus, the new nuclei are not completely identical to each other in the composition of their hereditary substance; their difference is also due to crossing over, which occurs with chromosomes at stage B. E - G - mitotic division of each of the haploid nuclei D: longitudinal splitting of each chromosome, divergence of chromosomes to the poles, the formation of two haploid nuclei from each nucleus D. As a result, 4 cells appeared with haploid nuclei instead of one cell with a diploid set of chromosomes
Organelles are not distributed strictly equally between daughter cells, but then their constituent parts are synthesized in the cells, new copies of organelles of each species are assembled, and their number in each cell is restored to normal. The cells themselves grow. In chromosomes, the hereditary material is duplicated, after which the chromosomes consist, as before cell division, of two chromatids. The cell is ready for new division.
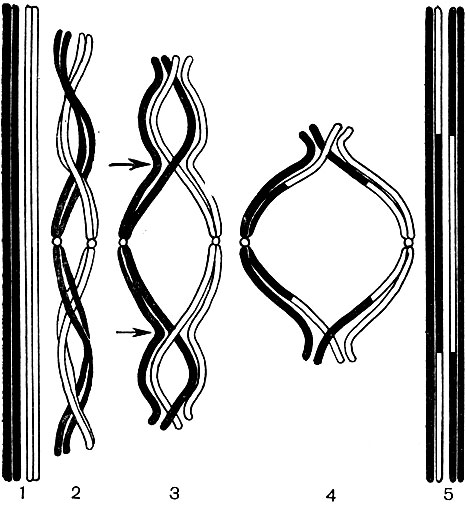
Figure: 38A. Crossover diagram during meiosis occurring at stage B Fig. 38. 1 - two homologous chromosomes, each of which consists of two sister chromatids; 2 - homologous chromosomes are intertwined with each other; 3 - each chromosome is split into its constituent chromatids; chromosomes begin to repel each other and diverge, but at some points (in the figure - at four) they remain connected; sister chromatids are doubled in the central loop, and non-sister chromatids in both extreme loops; points where crossing over is possible are shown by arrows; 4 - chromatids have exchanged sections, and chromosomes, having undergone a kind of mutual hybridization, diverge; 5 - two diverged, but already "hybrid" (unlike those shown on 1) chromosomes
When the diploid generation of cells is replaced by the haploid one, the so-called reduction nuclear division - meiosis occurs. During meiosis (Fig. 38), the homologous chromosomes of each pair approach each other, closely adjoin each other along their length, and twist. Separate sections are exchanged between contiguous homologous chromosomes. As a result, some of the genes of the paternal chromosomes are transferred into the composition of the maternal chromosomes, and the corresponding genes of the maternal chromosomes occupy the vacated places in the paternal chromosomes (crossing over phenomenon - Fig. 38A). As a result, the appearance of those and other chromosomes does not change, but their qualitative composition becomes different. Paternal and maternal inheritance is redistributed and mixed. Further, the nuclear envelope and the nucleolus dissolve, a spindle apparatus is formed, the same as during mitosis. Homologous chromosomes are separated and, with the help of spindle threads, diverge to the poles of the cell. One pole has one haploid set of chromosomes (one homologous chromosome from each pair), the other has a second haploid set.
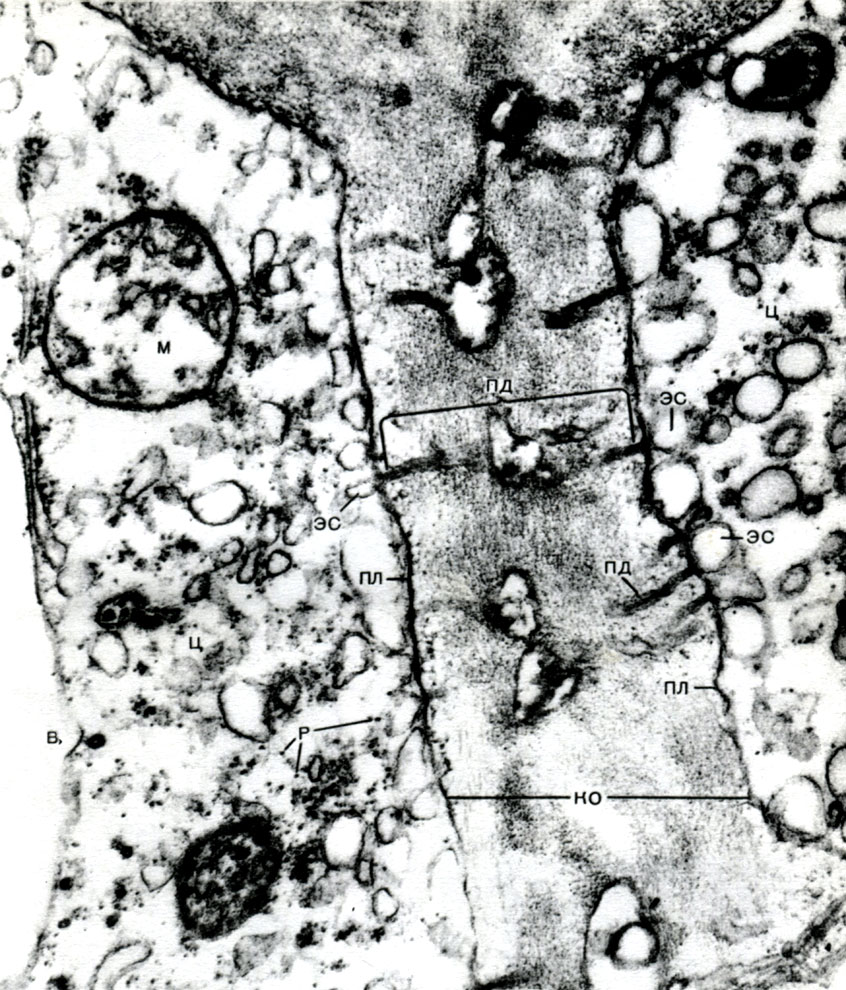
Following meiosis, the formed haploid nuclei divide by mitosis. In this case, each chromosome of the haploid set is split into two chromatids, they diverge and daughter haploid cells are formed.
Meiosis differs from mitosis in two fundamental ways. First, during meiosis, each pair of homologous chromosomes clump together and a part of the hereditary material is exchanged between them, which is not the case during mitosis. Secondly, during meiosis, homologous chromosomes diverge to the poles of the cell, one from each pair, and during mitosis, each chromosome is split into chromatids and chromatids diverge to the poles, one from each chromosome. As a result of meiosis, haploid cells are formed from a diploid cell.
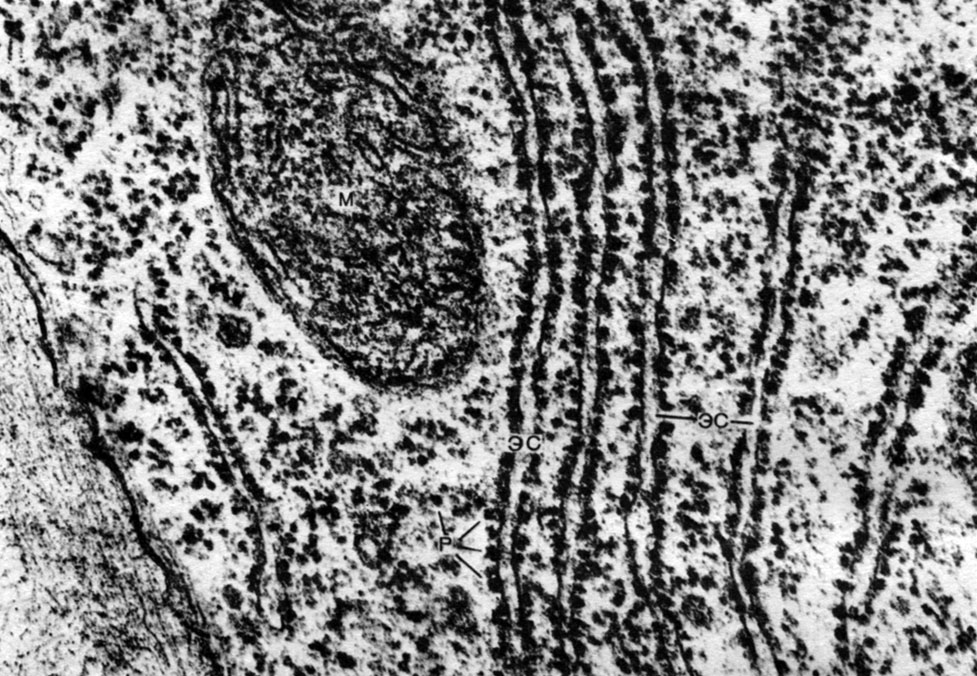
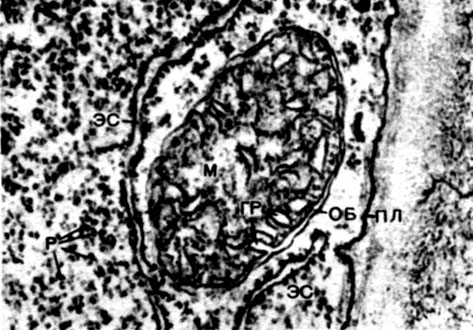
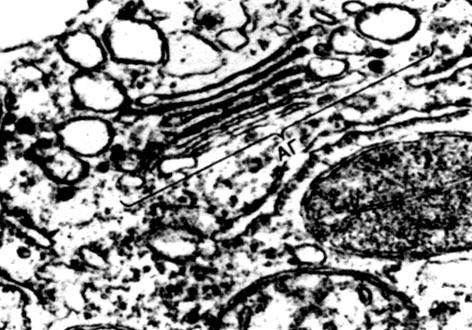
Table 10. Plant cell structure. Golgi apparatus in the cytoplasm of the developing root hair of a radish (Raphanus sativus). Electron micrograph (magnification x 52000) by M.F.Danilova: ag - Golgi apparatus, sections of flat sacs and bubbles are visible
In the cells of some tissues, during their development, incomplete mitosis occurs: the chromosome material in the nuclei doubles, the chromosomes divide in half, but instead of forming two nuclei, they remain in the original nucleus. From that moment on, it contains not a diploid, but a tetraploid (quadruple) set of chromosomes. The process leading to this duplication of chromosomes within one nucleus is called endomitosis - internal mitosis. If it happens to one nucleus twice, then it becomes an octoploid (eightfold) set, etc. Cells whose nuclei carry more than two sets of chromosomes are called polyploid, that is, multiploid. Cell polyploidy in some cases increases their viability, since each gene is duplicated by several other genes of the same. Single-quality genes act in unison, and damage to any of them does not lead to the loss of the trait it determines, since it is compensated by the work of the remaining homogeneous genes. In many cases, polyploid cells are larger and richer in content than diploid cells. The varieties of polyploid cultivated plants have been developed, which have increased economic qualities.
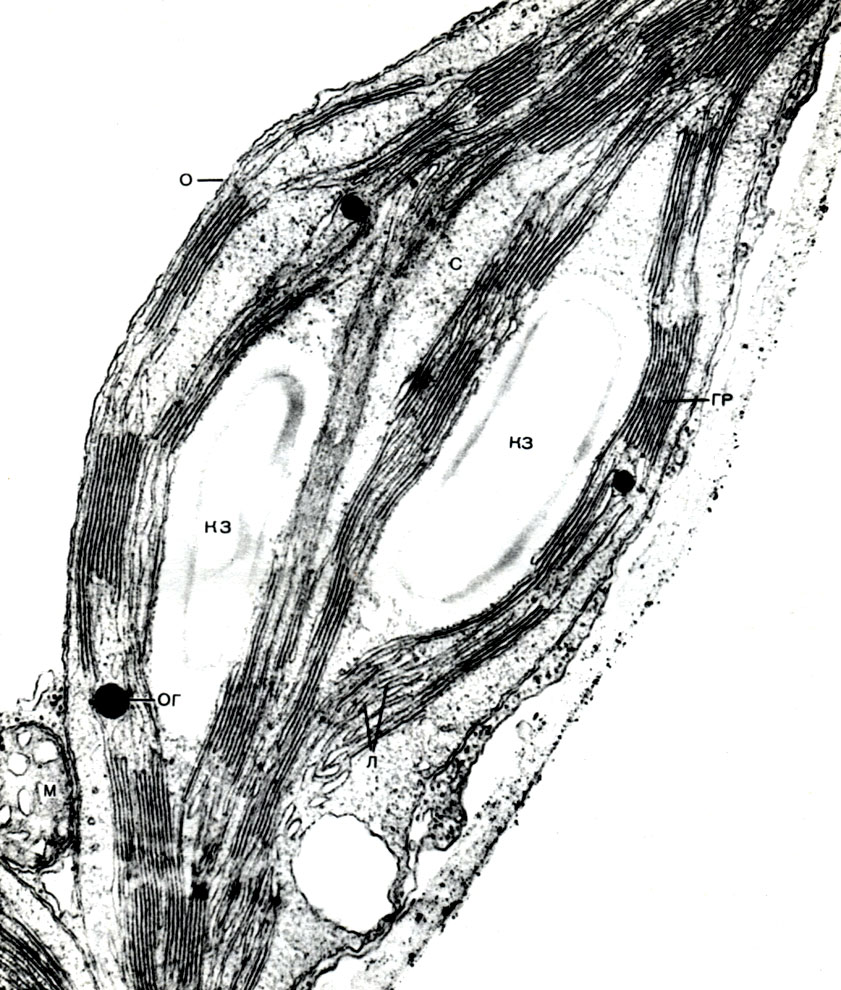
The science that studies the cell is called cytology. The cell is the basis of the structure of all living things and all life processes; therefore, most of the most important general biological problems can be solved with the involvement of cytology and methods of its research.
With the help of cytology, many practically important issues are solved: the fight against plant diseases (fungal, bacterial), the breeding of new varieties of cultivated plants, overcoming the sterility (sterility) of hybrid varieties.
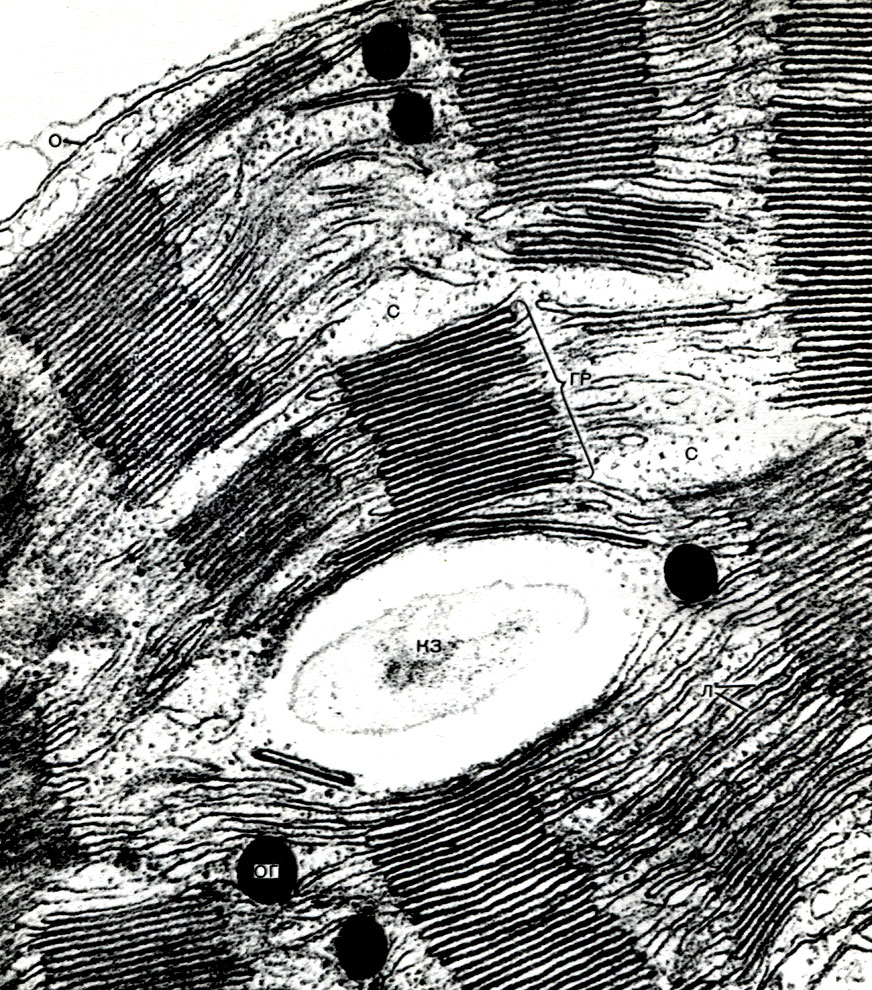
Modern cytology investigates these problems using a variety of methods - microscopic, biochemical, biophysical, genetic - and closely interacting with other related biological sciences.