Випадок 2: Закриття Кореня і Створення кератінізірованних Тканини. У пацієнта локалізована. будову рослинної клітини, тканинах, вегетативних і репродук- тивних органах ... стьях зустрічається столбчатая (палісадна) (рисунок 9), губчаста. Клітини майже всіх багатоклітинних рослин не однорідні, а зібрані в Т. У нижчих рослин Т. ... система (хлорофіллоноснимі паренхіма, губчаста і столбчатая Т.). ... на якій рослинний організм розвивається. До цієї. Тематичний тест по темі «Будова і функції клітин», 10 клас. 1 варіант. Частина А В єдиному рослинному організмі все клітини, подібні за будовою і. газ і проникає через продихи до межклетникам губчастої тканини, а звідти до. Показані палісадна (зверху, щільно упаковані клітини) і губчаста (знизу, пухко. Рихлість губчастої тканини відіграє важливу роль в газообміні листа. Рослинні химери) в результаті соматичних мутацій можливо. утворюють певний малюнок і форму, а потім опадають, в той час як. Dateiformat: PDF / Adobe Acrobat Тут знаходиться освітня тканину, що дає зростання листової пластинки і ... Губчаста тканина - основна тканина, клітини якої мають округлу форму, ... якщо виконати нескладне пристосування, як показано на малюнку .. Весь процес дихання протікає в клітинах рослинного організму. . з натурального вигодовування на коров'яче молоко з рослинними добавками .. Матриця для солей кальцію всередині клітин - лізосоми і мітохондрії .. У ділянках з вапном може формуватися кісткова тканина, а навколо неї буде. Нефрокальциноз при діагностиці відрізняють від губчастої нирки,.
Губчастої тканини (губчаста паренхіма), нижня пухка частина м'якоті листа. Характеризується неправильною формою клітин і великими межклетниками. набивная бавовняна тканина з східним малюнком. [Правка] ... при якій контрастну речовину вводять в губчасту тканину остистоговідростка ... рослинна тканина, що складається з клітин, видом дуже схожих на камбій. На малюнку зображена частина поперечного розрізу листа фісташки з дуже. Клітини вентиляційної губчастої тканини часто мають вельми. Продукти внутрішньої секреції рослинних організмів розміщені або в. В єдиному рослинному організмі все клітини, подібні за будовою і. газ і проникає через продихи до межклетникам губчастої тканини, а звідти до. Тканина - це зібрання клітин, подібних за будовою і функціями .. клітини губчастої паренхіми; 5 - замикають клітини продихів, щілину між кожною їх парою. Цитоплазма у всіх представлених на малюнку клітинах розташована. Клітини мезофіла відносяться до основної хлорофіллоносной паренхімі .. клітини губчастої тканини більш-менш ізодіаметрічни і, як правило ,. Тут знаходиться освітня тканину, що дає зростання листової пластинки і ... Губчаста тканина - основна тканина, клітини якої мають округлу форму, ... якщо виконати нескладне пристосування, як показано на малюнку .. Весь процес дихання протікає в клітинах рослинного організму. У більшості рослинних клітин присутні порожнини - вакуолі ,. Клітини механічних тканин мають потовщену оболонку. 15. ... II - губчастої тканини. ... Замалюйте рослина картопля, вказавши на малюнку плід і бульба. Тканина - група клітин, які мають спільне походження, виконують одну або. Приклад перших - столбчатая паренхіма, губчаста паренхіма ,. Викладачі Анкети кращих репетиторів. Знайти репетитора Заповніть форму запиту, і вам. Поставити питання про сучасній освіті, висловити ідею або вирішити назрілу проблему.
5 Вересня 2014. Пробка, або феллема (лат. Phellema) - вторинна покривна тканина осьових. Як згадувалося вище, клітини феллеми відкладаються. У них виявлено мезофіл тільки помірковано- губчастого. (Гомогенний). (Широко. Анатомічні малюнки виконані за допомогою малювального апарату.. Обсяг однієї клітини, число клітин в одиниці площі вайи. У всіх рослин ці тканини по загальній формі їх клітин відносяться до Паренхімні. Епідерміс, 4 - водоносна тканину, 5 - столбчатая паренхіма, 6 - губчаста паренхіма.. Зробити схематичний малюнок, позначивши водоносну тканину,. рісунак дерева з олівцем. як обклеїти пінопластом яйце. купити комірець плетений з бісеру губчастої тканини (губчаста паренхіма), нижня пухка частина м'якоті листа. Характеризується неправильною формою клітин і великими межклетниками. малюнок 1 - Елементи анатомічної будови епідермісу листка. Внутрішня частина листа заповнена пухкими клітинами губчастої тканини. Губчаста тканина у таких хворих дуже часто на всьому протязі була заміщена. Збережені ендотеліальні клітини мали ознаки набухання. 5 Вересня 2014. Пробка, або феллема (лат. Phellema) - вторинна покривна тканина осьових. Як згадувалося вище, клітини феллеми відкладаються.Сполучна тканина складається з клітин різних типів,. Малюнок 9.3.2.1. . Губчаста кісткова тканина складається з тонких кісткових елементів,. Губчастої тканини (губчаста паренхіма), нижня пухка частина м'якоті листа. Характеризується неправильною формою клітин і великими межклетниками. Мікрофотографія губчастого речовини, що демонструє його трабекули (рожеві) і мозкову речовину (синє). . Губчаста кісткова тканина (губчаста речовина, трабекулярную речовина, лат.. Клітини, хондробласти, хондроцит. Неспеціалізовані в структурному відношенні клітини паренхіми проте метаболічно активні: багато. Особливо розвинена система повітроносних межклетников в губчастої паренхіми. 5.. Малюнки в біології. Показані палісадна (зверху, щільно упаковані клітини) і губчаста .. утворюють певний малюнок і форму, а потім опадають, в той час як. клітини мезофіла відносяться до основної хлорофіллоносной паренхімі. Вони тонкостінні і. клітини губчастої тканини більш-менш ізодіаметрічни і, як правило , розташовуються дуже пухко.. Випадковий малюнок. Малюнки виконували за допомогою малювального апарату РА-4М.. Клітини верхнього і нижнього епідермісу ромашки аптечної сорти «Карагандинська» дрібні,. У губчастої паренхіми розташовані резервуари з ефірною олією. 30 Січня 2009. Це спостерігається при заміщення старих форм клітин остеоцитів і. Визначається на рентгенограмі крупнопетлістий малюнок кістки . Щодо губчастої кістки альвеолярного відростка внутрішній. Неспеціалізовані в структурному відношенні клітини паренхіми проте метаболічно активні: багато. Особливо розвинена система повітроносних межклетников в губчастої паренхіми. 5.. Малюнки в біології.
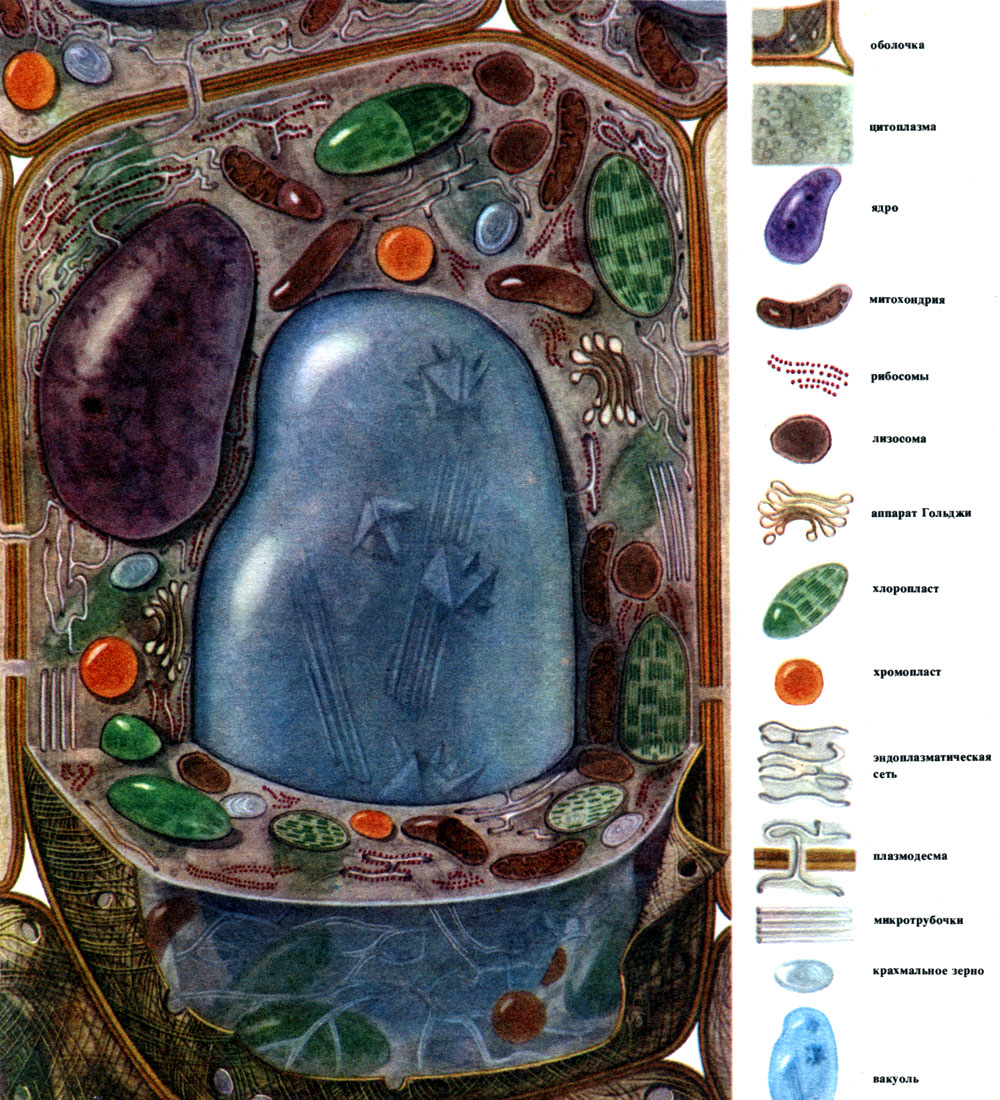
Рослинна клітина (Н. І. Арронет)
Рослина, як і будь-який живий організм, складається з клітин, причому кожна клітина породжується теж кліткою. Клітка - це найпростіша і обов'язкова одиниця живого, це його елемент, основа будови, розвитку і всієї життєдіяльності організму.
Існують рослини, побудовані з однієї-єдиної клітини. До них відносяться одноклітинні водорості і одноклітинні гриби. Зазвичай це мікроскопічні організми, але є і досить великі одноклітинні (довжина одноклітинної морської водорості ацетабулярії досягає 7 см). Більшість рослин, з якими ми стикаємося в повсякденному житті, - це багатоклітинні організми, побудовані з великої кількості клітин. Наприклад, в одному аркуші деревної рослини їх близько 20 000 000. Якщо дерево має 200 000 листів (а це цілком реальна цифра), то число клітин в усіх них становить 4 000 000 000 000. Дерево в цілому містить ще раз в 15 більше клітин .
Рослини, за винятком деяких нижчих, складаються з органів, кожен з яких виконує свою функцію в організмі. Наприклад, у квіткових рослин органами є корінь, стебло, лист, квітка. Кожен орган зазвичай побудований з декількох тканин. Тканина - це зібрання клітин, подібних за будовою і функціями. Клітини кожної тканини мають свою спеціальність. Виконуючи роботу по своїй спеціальності, вони вносять вклад в життя цілого рослини, яка складається в поєднанні і взаємодії різних видів роботи різних клітин, органів, тканин.
Основними, найбільш загальними компонентами, з яких побудовані клітини, є ядро, цитоплазма з численними органоидами різної будови і функцій, оболонка, вакуоль. Оболонка покриває клітку зовні, під нею знаходиться цитоплазма, в ній - ядро \u200b\u200bі одна або декілька вакуолей. Як будова, так і властивості клітин різних тканин в зв'язку з їх різною спеціалізацією суттєво різняться. Перераховані основні компоненти і органели, про які йтиметься далі, розвинені в них в різному ступені, мають неоднакову будову, а іноді той чи інший компонент може зовсім відсутніми.

Найголовнішими групами тканин, у тому числі побудовано вегетативні (безпосередньо не пов'язані з розмноженням) органи вищої рослини, є наступні: покривні, основні, механічні, провідні, видільні, меристематические. У кожну групу зазвичай входить декілька тканин, що мають схожу спеціалізацію, але побудованих кожна по-своєму з певного виду клітин. Тканини в органах не ізольовані один від одного, а складають системи тканин, в яких елементи окремих тканин чергуються. Так, деревина - це система з механічною і проводить, а іноді і основний тканини.


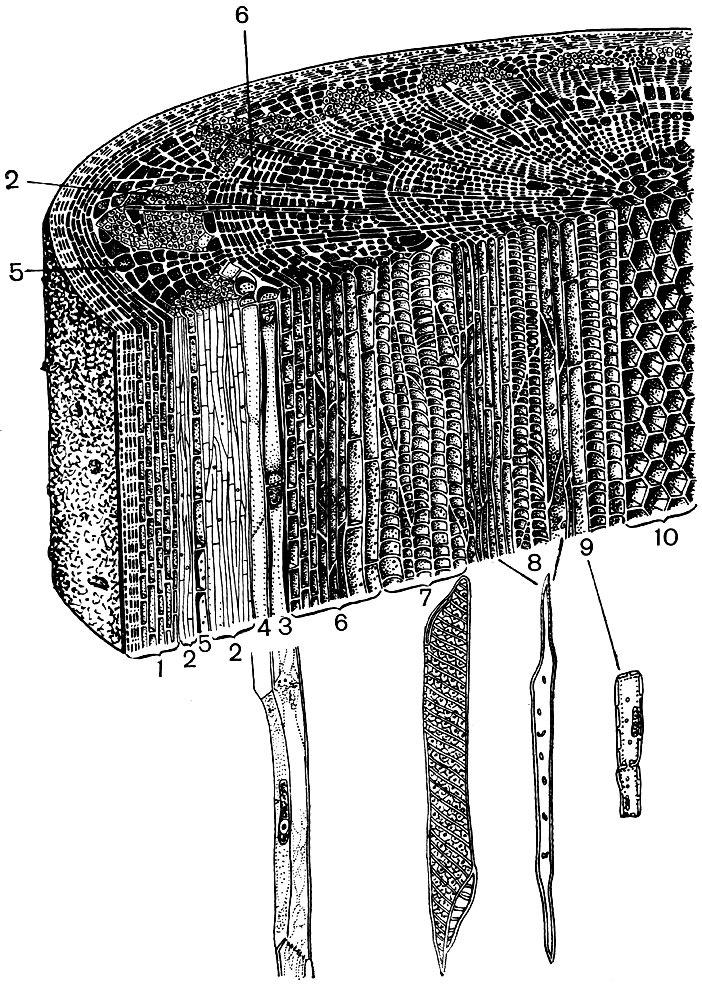
Візьмемо для прикладу два органу вищого рослини - лист і стебло. І ми побачимо, наскільки різноманітні за будовою і роботі клітини, що утворюють їх тканини (рис. 22 і 23). Ми побачимо також, як пристосоване будова клітин кожної тканини до виконання їх спеціальних функцій.


Покривні тканини - епідерміс, пробка, кірка - захищають органи рослини від несприятливих впливів: від висихання, перегріву, переохолодження, променевої енергії, механічних пошкоджень, зайвого намокання, від проникнення чужорідних організмів, наприклад бактерій. Епідерміс покриває обидві поверхні листа, молоді пагони і пелюстки. Як правило, це один шар клітин. Ті стінки клітин, якими вони стикаються між собою, зазвичай звивисті, причому виступ однієї заповнює увігнутість сусідній. Це сприяє міцному з'єднанню клітин, які в результаті утворюють єдиний щільний пласт. Зовнішні, які межують з повітрям стінки клітин більш товсті.

Епідермальні клітини виділяють речовини, що відкладаються на них зовні у вигляді плівки (наприклад, кутин, який утворює щільну кутикулу, і віск; кутин до того ж просочує і самі зовнішні стінки епідермальних клітин). Товста зовнішня оболонка і зовнішня плівка з Кутіна і воску забезпечують ізоляційні, захисні властивості цієї тканини. Крізь її шар утруднене проникнення газів, води, бактерій. Однак повітря обов'язково повинен входити всередину зелених частин рослини, а назовні виходять водяні пари і кисень. Обмін газами, а також водяною парою між атмосферою і внутрішніми тканинами молодих органів, покритих епідермісом, здійснюється за допомогою продихів.
Устячка утворені спеціальними клітинами епідермісу, між двома з яких (так званими замикаючими клітинами продихи) є отвір регульованої величини - устьичная щілину. Через неї здійснюється зв'язок між атмосферою і воздухоноснимі міжклітинними просторами товщі аркуша або іншого органу, покритого епідермісом. Замикають клітини продихів під впливом зміни умов вологості або освітлення змінюють свою форму, зближуючись між собою або розмикаючи. При цьому вони відкривають або, відповідно, закривають устьічнимі щілину. На світлі, коли рослина фотосинтезує і потребує припливу вуглекислого газу з атмосфери, устьічниє щілини відкриті. Вночі вони закриваються; замикають клітини закривають просвіти устьиц і в жарку пору дня, що охороняє рослину від великої втрати води, від в'янення.
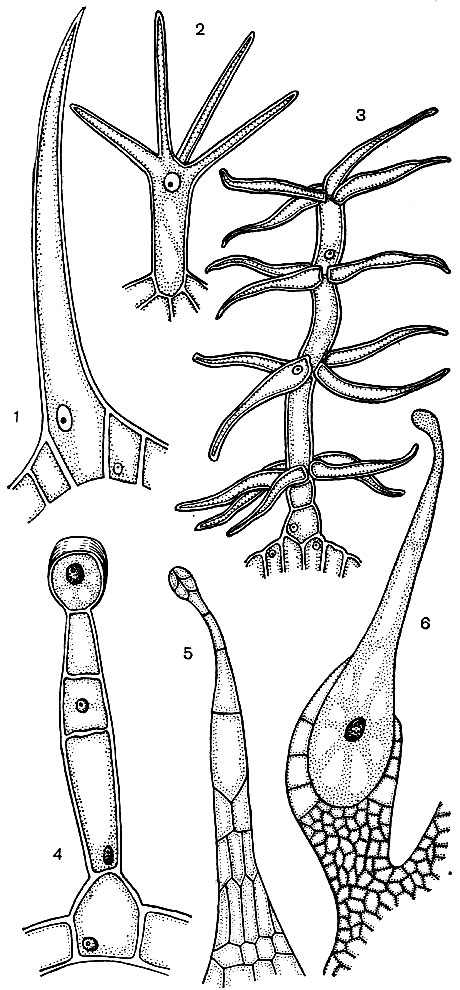
Часто епідермальні клітини утворюють вирости - волоски (рис. 24). Іноді це багатоклітинні освіти, в інших випадках кожен волосок є відросток однієї клітини, що лежить в загальному шарі епідермісу. Вони грають захисну, опорну (наприклад, у кучерявих або сланких рослин), видільну роль. Важливі кореневі волоски - трубчасті вирости епідермальних клітин кореня. Вони збільшують всмоктувальну поверхню останнього. Підраховано, що на 14 млн. Бічних коренів однієї рослини жита розвивається до 14 млрд. Кореневих волосків. Площа поверхні коренів становить при цьому 232 м 2, а волосків - 400 м 2. Епідерміс багатьох насіння або плодів утворює волоски, що сприяють розселенню насіння, а тим самим рослин. Часто волоски використовує і людина. Приклад цього - бавовна, що представляє собою епідермальні волоски насіння бавовнику; його волоски мають товсті целюлозні оболонки і досягають в довжину 70 мм.
У стеблі багаторічних рослин під епідермісом, на зміну йому, розвивається більш груба захисна тканина - пробка. Клітини її відмирають, втрачають протопласт і складаються тільки з товстих стінок, які оточують порожнину, заповнену повітрям або смолистими речовинами. Стінки просякнуті суберином, що робить їх водо- і повітронепроникними, теплоізоляційними, а також міцними, пружними. Вони можуть до того ж і одревесневать. Прикладом цієї тканини служить береста. Вона досить тонка, а ось пробка коркового дуба досягає товщини в кілька десятків сантиметрів. Пробка в деяких місцях переривається чечевичками - утвореннями з іншої тканини, яка проникна для води і повітря. Через чечевички здійснюється обмін внутрішніх частин стовбура з навколишнім середовищем. Чечевички розвиваються на місцях устьиц.
У стеблі під покривними тканинами перебувають клітини лубу. Луб - це система з елементів декількох тканин: провідної, опорної, основний. Найважливіший елемент лубу - сітовідние трубки. Вони побудовані з подовжених клітин, витягнутих уздовж стебла, зчленованих один з одним кінцями. Це живі, але без'ядерні клітини, елементи цитоплазми в яких розташовані пристенно. Оболонки в місцях стику цих клітин мають численні отвори, так що перегородки подібні ситу. В результаті цього суміжні клітини повідомляються між собою і тим самим виявляються об'єднаними в довгі трубки, що тягнуться крізь жилки і черешки листя, по стеблу і корені.
За системою сітовідних трубок продукти, що утворюються в зелених частинах рослин, переміщаються до всіх його іншим частинам, живлять їх. Основним з продуктів, що транспортуються є сахароза. Ситовідниє трубки - це елемент провідної тканини. Близько клітин сітовідних трубок є клітини-супутники. Вони містять ядра, і їх протопласт має безпосередні зв'язки з цитоплазмою без'ядерних сітовідних клітин. У лубе зустрічаються також ділянки, що складаються з паренхімних клітин (т. Е. Клітин, довжина, ширина і висота яких близькі за величиною). У них відкладаються різні речовини - крохмаль, масла, смоли. Це клітини основної тканини.
Крім того, луб містить елементи опорної тканини - луб'яні волокна. Це дуже довгі клітини з потовщеними стінками; довжина клітини може перевищувати її ширину в тисячі разів. У льону довжина тонких витягнутих луб'яних волокон досягає 4 см, а у рами - навіть 35 см. Зазвичай це мертві клітини, без протопласта. Вони виконують механічну функцію, створюючи міцність стебла.
Більшість спеціалізованих клітин не здатна до розмноження. Однак рослина росте все своє життя, і протягом усього життя в ньому утворюються нові клітини. Вони розвиваються з клітин освітніх - меристематичних - тканин. Розмноження поділом є спеціалізацією меристематичних клітин, їх функцією в організмі. Відбулися з них клітини розвиваються, перетворюючись в ті чи інші спеціалізовані клітини - клітини сітовідних трубок, клітини пробки, волокна і т. Д. Освітня тканина знаходиться в різних частинах рослини (наприклад, в точках зростання - на верхівках пагонів, коренів). У товщі стебла зазвичай є кілька шарів освітньої тканини. Під шаром пробки знаходяться клітини феллогена, які, ділячись, поповнюють число клітин пробки, кірки. Всередину від лубу розташовується шар клітин камбію. Ті з породжуваних ними клітин, які утворюються зовні від камбію, розвиваються в клітини лубу (рис. 25). Що опинилися всередині, т. Е. Ближче до осі стовбура, дають початок клітинам деревини.
Деревина складається з елементів провідної, опорної і основний тканин. До перших відносяться волокна деревини - довгі мертві клітини з здеревілими стінками; до других - судини, що представляють собою результат злиття багатьох клітин; до третіх - клітини деревної паренхіми. Клітини, що диференціюються з камбіальних елементів в провідні (судини), ростуть в довжину і ширину. Їх бічні стінки товщають і дерев'яніють. У стінках, однак, залишаються пори, закриті лише тонкою оболонкою. Перегородки на стиках суміжних клітин зникають, протопласт відмирає. В результаті утворюються довгі посудини, що складаються з одних стінок. Провідна система тягнеться крізь корінь і стебло в лист. За таким судинах здійснюється висхідний струм води і розчинених в ній солей від коріння до всіх органів рослини.
У центрі стебла знаходяться клітини серцевини - округлі або багатогранні паренхімні клітини. Це елементи основної тканини. Іноді вони порожнисті, і в них знаходиться повітря; іноді вони заповнені запасними поживними речовинами, різними кристалами, таніни. Стінки їх можуть бути здеревілими. Деревину і луб пронизують радіальні промені. Їх клітини є похідними камбію і несуть запасаючу функцію. На рівні лубу ці промені помітно розширюються.
У м'якоті листа між верхнім і нижнім епідермісом знаходиться основна тканина - клітини з тонкими оболонками і великою кількістю зелених пластид - хлоропластів. У цих клітинах відбувається фотосинтез. Верхні шари складаються здебільшого з довгастих клітин, щільно прилеглих один до одного, - ця столбчатая паренхіма. Нижні шари рихлеет між клітинами розташовані межклетники - простору, заповнені повітрям; це губчаста паренхіма. Паренхіма пронизана розгалуженим жилками, які побудовані з пучків механічної (різноманітні волокна) і проводить (сітовідние трубки і водоносні судини) тканин.
![]()
Це короткий і схематичне виклад плану будови стебла, листа показує, наскільки різноманітні за величиною, формою, будовою, функції клітини одного і того ж рослини. Якщо взяти різні рослини, то можна зустрітися з ще більшим різноманіттям клітин. Однак, як правило, клітини однойменних тканин навіть дуже далеких один від одного рослин подібні, оскільки ці клітини виконують аналогічні функції. Клітини епідермісу листка берези і, наприклад, кульбаби більш схожі один на одного, ніж епідермальна клітина кульбаби і його ж сітовідной клітина. Відмінні риси клітини пов'язані в першу чергу з її спеціальною функцією. Клітини, спеціалізовані в механічної функції, мають потовщені і нерідко здеревілі оболонки. Спеціалізація в фотосинтезі веде до появи в клітинах хлоропластів. Спеціалізація в провідній функції пов'язана з подовженням клітин, з втратою протопласта, зі змінами в оболонках клітин на стиках, завдяки яким порожнини суміжних клітин повідомляються. Для клітин, спеціалізованих в захисної функції, характерні різноманітні зміни зовнішніх стінок, наявність волосків, здатність виробляти захисні речовини.
Спеціалізація в функції запасання поживних речовин може привести до збільшення розмірів клітин, до появи в них дуже великих вакуолей. У клітинах меристематичних, посилено діляться, особливо розвинені ті внутрішньоклітинні структури, які забезпечують синтез речовин - складових частин клітини.
Залежно від характеру спеціалізації одні риси будови і роботи клітини розвинені дуже сильно, інші, навпаки, мало розвинені або зовсім відсутні. Як ми бачили, деякі клітини, ставши спеціалізованими, вмирають і саме в мертвому стані починають виконувати свою спеціальну функцію в багатоклітинних організмі (клітки пробки, деревинні волокна, судини).
Дуже різноманітні і своєрідні клітини багатоклітинних нитчастих водоростей і одноклітинних рослин (рис. 26 і 27). Клітка будь-якого з останніх до того ж сильно відрізняється від клітин багатоклітинних рослин. Їй одній доводиться виконувати кілька функцій, які у багатоклітинних рослин поділені між клітинами різних тканин.
У той же час навіть дуже відрізняються між собою клітини володіють глибоким схожістю в будові і функціях. Для багатоклітинних це пов'язано, по-перше, з тим, що всі клітини організму (якщо виключити вегетативне розмноження, при якому від материнської рослини відразу відділяється в якості нової особини або її зачатка цілий багатоклітинний комплекс) є нащадками однієї і тієї ж клітини-родоначальниці. Тому, яким би чином не були спеціалізовані клітини, вони мають загальний витік, а тому є родичками.
По-друге, спільні риси в будові клітин рослин різних видів пов'язані з тим, що всі рослини складаються в тій чи іншій ступеня споріднення. Всі рослини розвинулися шляхом еволюції від одноклітинних спільних предків. Загальні риси успадковані клітинами сучасних рослин від древніх прабатьківських клітин. З цим пов'язано також наявність спільних рис будови і роботи рослинних і тваринних клітин.
По-третє, подібність пов'язано з тим, що всі живі клітини, яку б спеціальну функцію вони ні несли в організмі, перш за все повинні забезпечувати власне життя. Клітини поглинають поживні речовини, переробляють їх, добуваючи енергію і ладу власне тіло, дихають, звільняються від непотрібних речовин, борються з різними ушкодженнями, реагують на зміни зовнішніх умов, перебудовуючи свою життєдіяльність, ростуть. Всі ці процеси у різних клітин здійснюються подібно і за допомогою однотипних структур, загальних за планом будови не тільки для різних рослинних клітин, але і для клітин рослин і тварин. Треба сказати, що виконання будь-якої спеціальної функції клітини ґрунтується на її загальні властивості. Та чи інша риса, здатність, притаманна всім клітинам, у спеціалізованій клітини розвивається особливо сильно і забезпечує виконання клітиною її основний, спеціальної функції. Ті загальні риси, без яких неможливе виконання цієї спеціальної функції, в клітці зберігаються, а інші можуть втратитися. Мертві спеціалізовані клітини - крайній, граничний випадок цього. Спеціальна функція таких клітин пов'язана з їх оболонкою; протопласт потрібен лише до тих пір, поки він створює оболонку; після цього він відмирає, і вся клітина складається тільки з неживої оболонки, яка і працює на потреби рослини.
Розглядаючи загальні риси будови і життєдіяльності рослинних клітин, зручно говорити про якусь типовий клітці, що увібрала в себе загальні риси різних спеціалізованих клітин. Таку клітку, хоча вона і не існує в "чистому вигляді", можна навіть зобразити (табл. 7, рис. 28). З реальних клітин більш інших схожі на таку "типову" клітини паренхіми листа (табл. 8, рис. 29).
Перейдемо до опису будови і роботи рослинної клітини.
Таблиця 5. Покритонасінні: угорі ліворуч - квітка магнолії; вгорі праворуч - квітки яблуні; внизу - лотос.
Таблиця 6. Покритонасінні: вгорі (зліва направо) - дзвіночки, пеларгоніум, шафран (крокус); внизу зліва - айстри; внизу праворуч - реп'ях.
Зовні рослинна клітина покрита оболонкою, неоднакової по товщині і будовою у різних клітин. Утворюють її речовини виробляються в цитоплазмі і відкладаються зовні від неї, поступово створюючи оболонку. Цими речовинами насамперед є крупномолекулярні полісахариди - пектин, геміцелюлоза і в невеликих кількостях целюлоза. Вони утворюють так звану первинну оболонку. Вона досить еластична, у міру зростання клітини розтягується і теж зростає, а тому не перешкоджає зростанню клітини. Однак вона створює певну міцність клітини і здатна захистити її від механічних пошкоджень. Є клітини, які позбавлені такої первинної оболонки, - це деякі клітини, службовці для безстатевого і статевого розмноження (зооспори і гамети водоростей і нижчих грибів, чоловічі гамети вищих рослин). У багатьох клітин є не тільки первинна, але ще і вторинна оболонка. Вона утворюється під первинної і побудована головним чином з целюлози. Целюлоза - це полісахарид, молекули якого утворюють найтонші нитки - мікрофібрили. В оболонці нитки целюлози занурені в аморфну \u200b\u200bречовину, що складається з пектинових з'єднань. У одних клітин ці мікрофібрили целюлози розташовані поперек довжини клітини, кільцями; завдяки цьому такі клітини можуть розтягуватися в довжину, але не в ширину (наприклад, клітини судин стебла). У інших нитки лежать поздовжньо; клітини з такою оболонкою еластичні при розтягуванні поперек, але дуже жорсткі на поздовжнє розтягання. У третє вони розташовані навскіс, утворюючи спіраль (епідермальні волоски насіння бавовнику, луб'яні волокна). Все це нагадує залізобетонні конструкції, причому нитки целюлози грають тут роль залізних прутів, а пектинові речовини - роль цементу. Клітини, що мають вторинну оболонку, досить міцні. Вони утворюють механічні, опорні тканини рослини. Іноді вторинна оболонка відіграє і роль складу поживних продуктів: що утворюють її речовини можуть перетворюватися в інші, більш прості, які витрачаються як харчування.
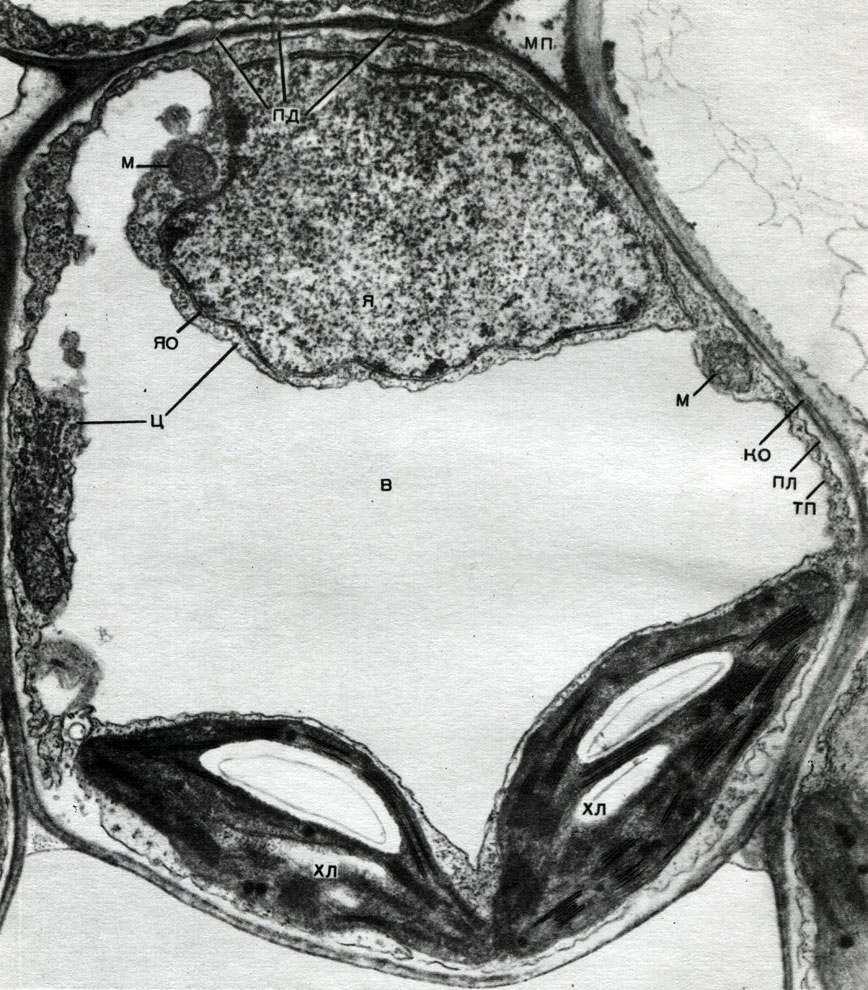
В оболонці є неутолщенние місця - пори (у первинній оболонці вони називаються норовить полями), через які здійснюється зв'язок між сусідніми клітинами. Крізь порові поля і пори проходять тонкі тяжі цитоплазми. Це плазмодесми, які пов'язують цитоплазму сусідніх клітин (табл. 9). За ним здійснюється обмін речовинами між сусідніми клітинами. Плазмодесми поряд з елементами провідної тканини з'єднують клітини і тканини організму в єдине ціле.
Обмін речовинами і поширення збудження дозволяють клітинам впливати на розвиток і роботу один одного, і кожна тканина впливає на життєдіяльність всіх інших тканин. Цим створюється координація роботи всіх частин єдиного організму-цілого рослини.
У багатьох клітинах клітинна оболонка з віком просочується речовинами, ще більш зміцнюють її. Накопичення в ній лігніну веде до одревеснению оболонки. Дерев'яніють стінки клітин деревини і, часто, луб'яних волокон, оболонки клітин шкірки насіння, іноді околоплодника (рис. 30) і навіть пелюсток. Клітини з здеревілими стінками не тільки міцніше, але і менш проникні для мікробів, для води. Оболонки деяких інших клітин - покривних тканин, а також на поверхнях поранений - опробковевают, просочуючись суберином. Вміст клітин після цього відмирає, так як суберин непроникний для води і газів, але тканина з таких мертвих клітин захищає внутрішні живі частини рослини від шкідливих зовнішніх впливів. Як вже говорилося, зовнішні оболонки клітин епідермісу зазвичай просочуються або покриваються кутином і воском, і це зберігає клітини від зайвого випаровування води, від проникнення інфекції. Оболонка може просочуватися і мінеральними речовинами, переважно солями кальцію.
Бувають, однак, в житті клітин і такі моменти, коли їх оболонка повинна стати менш міцною, ніж була до цього. Це відбувається, наприклад, при проростанні насіння.
Полісахаридна клітинна оболонка - характерна риса будови рослинної клітини, що відрізняє її від тваринної клітини.
Оболонка, або клітинна стінка, - це захисне утворення. Під оболонкою знаходиться цитоплазма. Самий зовнішній її шар, що примикає до оболонки, - поверхнева клітинна мембрана - плазмалемма. Вона являє собою комбінацію шарів жироподібних і білкових молекул. Такі мембрани називаються ліпопротеїновими ( "ліпосом" - жир, "протеїн" - білок). Мембрана подібної конструкції відмежовує цитоплазму від вакуолей, ця мембрана називається тонопластом. Багато органели клітини побудовані з ліпопротеїнових мембран. Однак в кожному випадку мембрана побудована з жироподібних речовин (ліпідів) і білків, властивих саме даній мембрані. Якісна розмаїтість ліпідів і особливо білків колосально, звідси величезна різноманітність мембран, що відрізняються за властивостями, і в межах однієї клітини, і в різних клітинах.
Плазмалемма регулює вхід речовин в клітку і вихід їх з неї, забезпечує виборче проникнення речовин в клітку і з клітки. Швидкість проникнення крізь мембрану різних речовин різна. Добре проникають через неї вода і газоподібні речовини. Легко проникають також жиророзчинні речовини, - ймовірно, завдяки тому, що вона має ліпідний шар. Передбачається, що ліпідний шар мембрани пронизаний порами. Це дозволяє проникати крізь мембрану речовин, нерозчинним в жирах. Пори несуть електричний заряд, тому проникнення через них іонів не цілком вільно. При деяких умовах заряд пір змінюється, і цим регулюється проникність мембран для іонів. Однак мембрана неоднаково проникна і для різних іонів з однаковим зарядом, і для різних незаряджених молекул близьких розмірів. В цьому проявляється найважливіша властивість мембрани - вибірковість її проникності: для одних молекул та іонів вона проникна краще, для інших гірше.
Речовина рухається в клітку, диффундируя в неї, якщо його концентрація зовні більше, ніж концентрація всередині. В іншому випадку воно повинно дифундувати з клітини. Але, проникнувши в клітку, молекули даної речовини можуть тут же залучитися до обмін речовин і піддатися перетворенню в інші речовини. Тоді концентрація цієї речовини всередині клітини знову падає, і нова порція його молекул дифундує всередину клітини. В результаті в клітини, активно витрачають ту чи іншу речовину, останнім посилено надходить з навколишніх клітин.
Інший механізм посиленого поглинання речовини кліткою складається в його зв'язуванні білками або іншими речовинами. Зв'язуючись, речовина йде з внутрішньоклітинного розчину і не перешкоджає подальшій дифузії цієї ж речовини. Воно продовжує надходити в клітку і накопичується в ній в пов'язаної формі. Так, в клітинах деяких морських водоростей йод накопичується в концентраціях, в мільйон разів перевищують його концентрацію в морській воді.
У всіх цих випадках мова йде про рух речовин шляхом дифузії. Однак клітинні мембрани мають у своєму розпорядженні і механізмами переміщення речовин через себе від меншої їх концентрації до більшої. Серед білків мембрани є білкові речовини, робота яких полягає в переміщенні різних речовин з одного боку мембрани на іншу. Вони називаються транспортними ферментами. Ймовірно, такий транспортний фермент утворює з стерпним речовиною проміжне з'єднання; Останнім проходить крізь мембрану, розщеплюється потім на вихідні частини, після чого фермент повертається назад, а перенесене речовина залишається по іншу сторону мембрани. Деталі цього процесу невідомі, але сам факт такого активного транспорту речовин безсумнівний. На відміну від дифузного, пасивного, ферментативний транспорт потребує затрати енергії. Ймовірно, саме завдяки такому активному транспорту іонів клітини епідермісу коренів здатні поглинати з грунту потрібні рослині неорганічні речовини і потім передавати їх по рослині від клітини до клітини.
При хімічних або фізичних змінах у зовнішній і внутрішньому середовищі клітинні мембрани змінюють свою проникність, а також ступінь і сам характер її вибірковості. На цьому грунтуються механізми регуляції руху речовин в клітку і з клітки. Зміна проникності мембран для живильних речовин відбивається на інтенсивності обмінних процесів в клітці, на характері що протікають в ній синтезів, на всій її життєдіяльності. У зміні проникності мембран для іонів натрію і калію полягає механізм поширення клітинного збудження - виникнення і переміщення біоструму. До поверхні мембрани приєднано велику кількість катіонів, головним чином іонів калію. Тому вона несе зовні позитивний заряд. Під впливом подразників іони калію отщепляются від ділянки, яка зазнала впливу, заряд ділянки падає і він стає електронегативний по відношенню до сусідніх ділянок мембрани. Ця електронний торгівельний є, в свою чергу, подразником для сусідніх ділянок, тим же шляхом знімаючи їх заряд, деполярізуя їх. Хвиля електронегативності поширюється по мембрані - це і є біострум. Потім іони калію знову сідають на мембрану, надаючи їй вихідний заряд, - за хвилею деполяризації слід хвиля відновлення. Поверхні плазмодесм, що проходять крізь клітинні стінки і з'єднують цитоплазму сусідніх клітин, теж утворені такими мембранами. Біострум рухається і по ним, поширюючись від клітки до клітки. Біоструми рослинної клітини поки ще мало вивчені. Однак ясно, що вони є способом сигналізації, що використовуються в рослинній клітині для пуску в хід одних хімічних реакцій і гальмування інших. Справа в тому, що всі хімічні процеси, що протікають в клітині і становлять сутність її життєдіяльності, йдуть за участю біологічних каталізаторів - ферментів. Кожна реакція можлива лише тоді, коли фермент, що пускає її в хід, активний. Більшість ферментів стають активними під дією тих чи інших неорганічних катіонів: К., Na. , Ca .., Mg .., Mn ... Біострум, змінюючи проникність мембран для заряджених речовин, створює умови для їх проникнення крізь мембрани і контакту з ферментами. Тим самим біострум включає в дію ті чи інші ферменти і цим регулює, направляє обмін речовин в клітині. Регуляція обміну речовин біострумами - це лише один з багатьох способів регуляції внутрішньоклітинного метаболізму.
Сказане про проникності поверхневої мембрани клітини - плазмалемме - відноситься і до інших внутрішньоклітинних мембран, в тому числі до тих, з яких побудовані багато органели клітини.
Цитоплазма, колись вважалася однорідним колоїдним розчином білкових речовин, насправді складно структурована. У міру розвитку мікроскопічної техніки дослідження з'ясовувалися всі більш тонкі деталі будови цитоплазми. У цитоплазмі були відкриті різні органели (органели) - структури, кожна з яких виконує певні фізіологічні та біохімічні функції. Найважливішими органоидами цитоплазми є мітохондрії, ендоплазматичний ретикулум (ендоплазматична сітка), апарат Гольджі, рибосоми, пластиди, лізосоми. У рухливих клітин (зооспори і гамети водоростей, сперматозоїди хвощів, папоротей, саговників, деякі одноклітинні і колоніальні водорості) є органели руху - джгутики.
Особливо багато нових фактів про тонку будову цитоплазми принесло і приносить використання електронного мікроскопа, що дозволяє досліджувати деталі будови самих органоїдів. Сучасні біофізичні і біохімічні методи дозволяють виділяти в чистому вигляді ті чи інші органели цитоплазми і потім вивчати їх хімічний склад і їх функції. Поза клітини, в середовищах складного складу, багато органели здатні виконувати ту роботу, яку вони виробляють, коли знаходяться в клітці.
Частина цитоплазми, в яку занурені органели і яка поки що представляється безструктурної, називається основною речовиною цитоплазми або гиалоплазмой. Гіало-плазма - це аж ніяк не пасивний наповнювач, а активно працююча частина цитоплазми. У ній протікає ряд життєво необхідних хімічних процесів, в її склад входять багато білків-ферменти, за допомогою яких ці процеси здійснюються.
Життя клітини складається в безперервної хімічної роботі, яка в своїй сукупності називається обміном речовин. По суті, клітина являє собою хімічний завод, що виробляє великий асортимент продукції і самостійно добуває енергію, необхідну для її виробництва. Її продукцією є речовини, які необхідні і їй самій для підтримки її власного життя (для побудови свого тіла при зростанні і розвитку, для заміни своїх знесення частини), і для створення дочірніх клітин при розмноженні, і для потреб інших клітин організму.
Всі хімічні реакції, що протікають в клітині, можна розділити на дві групи. В результаті одних ті чи інші речовини розпадаються на більш мелкомолекулярних. В результаті інших з мелкомолекулярних речовин синтезуються речовини з більшими молекулами. Молекула будь-якого речовини складається з атомів, які утримуються між собою хімічними зв'язками, т. Е. Тією чи іншою кількістю сконцентрованої потенційної хімічної енергії. Коли молекула дробиться, зв'язки рвуться і їх хімічна енергія звільняється. Для синтезу, т. Е. Для освіти більшої молекули з дрібних, потрібно створити нові хімічні зв'язки, В них необхідно вкласти деяку порцію енергії. Навпаки, реакції розпаду в кінцевому підсумку йдуть із звільненням енергії, так як при них зв'язку між атомами рвуться. Біологічний сенс реакцій розпаду, що йдуть в клітці, полягає в тому, що при них звільняється хімічна енергія, яка використовується потім кліткою для реакцій синтезу і для виробництва інших видів роботи (електричної, механічної, роботи по транспорту речовин). Так як клітина все своє життя синтезує різні речовини, то їй доводиться безперервно розщеплювати інші речовини. Звільняється енергія використовується для синтезів не відразу. Спочатку вона запасається шляхом утворення спеціальних речовин - акумуляторів хімічної енергії - аденозинтрифосфорної кислоти (АТФ) і споріднених їй з'єднань. У потрібний момент і у відповідній точці клітини АТФ розщеплюється і віддає енергію для синтезу необхідного клітці речовини.
Всі хімічні реакції, що протікають в клітині, - і синтезу і розпаду - здійснюються за допомогою ферментів. Ферменти - білкові речовини, що прискорюють перебіг реакцій. Прискорення це настільки велике, що без ферментів подібні реакції взагалі були б неможливі в клітці. Відомі випадки, коли завдяки ферменту реакція прискорюється в 1011 разів. Це означає, що реакція, що закінчується за участю ферменту протягом 0,01 сек, без нього протікала б 31 рік. Зрозуміло, що такі реакції без ферменту були б просто нереальними.
Крім того, завдяки ферментам протягом хімічних реакцій в клітині керовано, регульовано. Активність ферментів в клітці змінюється відповідно до її потребою в певному речовині або в енергії, т. Е. В кінцевому результаті роботи цих самих ферментів. Коли з'являється потреба в якомусь речовині, включаються в дію або заново синтезуються ті ферменти, завдяки яким воно утворюється. Освіта речовин, звільнення і запасання енергії - це підсумок послідовних реакцій, результат роботи цілого ланцюга ферментів. Всі структури живих частин клітини побудовані саме з ферментів і з речовин, що скріплюють ці ферменти. При цьому ферменти, що беруть участь в суміжних, послідовно протікають реакціях, і розташовані поруч. Вони передають молекули перетворюваних ними речовин один одному, як по конвеєру, причому кожен з них здійснює над молекулою свою робочу операцію.
У гіалоплазме містяться ферменти, що розщеплюють молекули глюкози на більш прості молекули піровиноградної кислоти. Звільняється при цьому енергія запасається шляхом утворення молекул АТФ. Той же процес протікає і в клітинному ядрі. Однак основна маса енергії добувається в особливих органелах цитоплазми - мітохондріях, так як там відбувається більш глибоке розщеплення речовин.
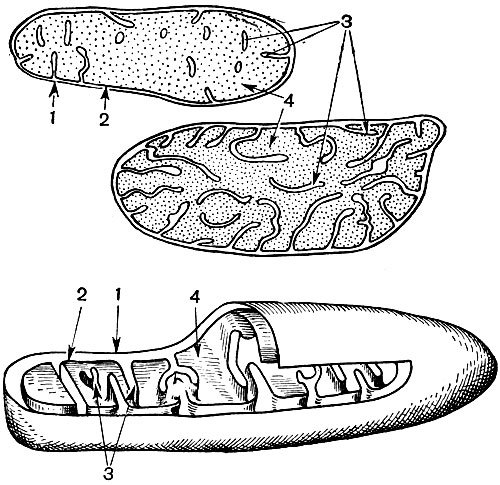
Мітохондрії - дрібні тільця округлої або довгастої форми, розміром 0,5 - 1,5 мк, т. Е. Величиною з бактерію. Число їх в клітці зазвичай велике, близько 100 - 3000. Бувають, однак, клітини і з малою кількістю мітохондрій. Так, в спермін морської водорості фукуса міститься всього 4 мітохондрії, а в одноклітинної водорості мікромонас - одна. Мітохондрії видно під світловим мікроскопом, однак їх тонка будова можна вивчати лише за допомогою електронного мікроскопа (табл. 10, схема будови - рис. 31). Мітохондрії - це утворення, побудовані з ліпопротеїнових мембран, занурених в основну речовину - матрикс. Оболонка мітохондрії утворена двома мембранами, між якими є проміжок.
Внутрішня з мембран оболонки дає численні впячивания всередину, це Крісті. Між ними знаходиться матрикс. І внутрішня мембрана оболонки мітохондрії, - і утворені нею Крісті побудовані з впорядкування розташованих ферментів. Завдяки складкам - Крістен робоча поверхня мембран всередині мітохондрій дуже велика. Ряд ферментів знаходиться в матриксі мітохондрії, т. Е. Між кристами.
Сукупність цих ферментів здійснює внутрішньоклітинне дихання і запасання звільняється при диханні енергії у формі АТФ. Робота мітохондрій тісно пов'язана з процесами, що йдуть в гіалоплазме, де протікають перші етапи розщеплення глюкози і інших речовин до піровиноградної кислоти. В мітохондріях же протікає подальше її розщеплення. Піровиноградна кислота проникає в мітохондрії і тут поступово, крок за кроком, окислюється до вуглекислого газу і води, причому одночасно споживається кисень. Це і є внутрішньоклітинний дихання, при якому клітина, розщеплюючи і окислюючи речовини, видобуває дуже багато енергії, яку вона потім може використовувати для самих різних своїх потреб.
Перший етап розщеплення молекули глюкози, під час якого вона дробиться навпіл і який протікає в гіалоплазме, дає клітці всього лише дві молекули АТФ.
В результаті другого етапу, який приводить до повного "згорянню" глюкози, утворюється ще 36 молекул АТФ. Тому мітохондрії за своєю функцією - це силові станції клітини, машини для добування основного кількості енергії. Саме розщеплення продуктів розпаду глюкози відбувається в матриксі мітохондрії, АТФ ж утворюється завдяки реакціям, розігрували на внутрішніх її мембранах, до складу яких входять дихальні ферменти і ферменти, що забезпечують освіту АТФ. Кількість крист в мітохондріях може бути різним. Чим їх більше, тим вище біохімічна активність мітохондрій.
Ми говоримо тут про глюкозу як про речовину, розщеплюючи яке клітина видобуває енергію. Глюкоза є центральним, але не єдиним з таких речовин. Молекула її має остов з шести атомів вуглецю, з'єднаних між собою. В результаті довгої і складної ланцюга реакцій її молекула дробиться, окислюється і, врешті-решт, розщеплюється на шість молекул неорганічної речовини - вуглекислого газу (СО2), кожна молекула якого містить лише один атом вуглецю, причому він гранично оксиди. Склавши всі послідовні реакції окислення глюкози і виключивши при цьому всі проміжні продукти, можна отримати сумарну реакцію цього процесу:
Крохмаль легко перетворюється в глюкозу, після чого вона піддається вищеописаному розщепленню. Білки і жири дають різні органічні кислоти, які перетворюються в проміжні продукти розпаду глюкози і далі окислюються таким же чином, як остання, і за допомогою тих же ферментів.
Повний біологічне окислення органічної речовини подібно його згорянню. В обох випадках результатом є вуглекислий газ, вода і виділяється енергія. Однак при горінні ця енергія виділяється у вигляді тепла, причому відразу повністю; при біологічному окисленні енергія хімічних зв'язків звільняється порціями, і основна її частина зв'язується, переходячи в енергію фосфатних хімічного зв'язку АТФ. У результаті клітина отримує концентрат енергії в такій формі, яка потім в потрібний момент і у відповідній точці може використовуватися для створення нових хімічних зв'язків, для синтезу нових речовин, а також для виробництва інших видів роботи - електричної, механічної, а також роботи по транспорту речовин з середовища в клітину, з клітки в середу і від клітини до клітини.
Чим активніше життєдіяльність клітини, тим більше у неї потреба в енергії і тим більше в ній мітохондрій. Вони і в межах однієї і тієї ж клітини можуть бути розподілені нерівномірно: їх більше в тій частині клітини, яка в даний момент працює активніше.
Мітохондрії здатні синтезувати частина тих речовин, з яких складаються вони самі. Завдяки цьому мітохондрії можуть розмножуватися.
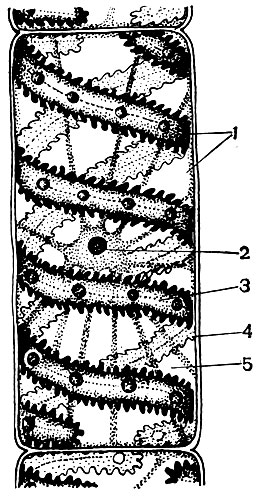
Ендоплазматичнийретикулум - органоид цитоплазми, в якому відбувається синтез дуже багатьох речовин (табл. 10). Ендоплазматичнийретикулум являє собою систему каналів, які пронизують цитоплазму і які в одних ділянках звужуються, в інших розширюються, утворюючи то цистерни, то плоскі мішки, то розгалужені трубки. Стінки всіх цих утворень побудовані з мембран, що включають в свій склад ферменти.
Як і в інших мембранних утвореннях клітки, ферменти в ретикулуме розташовані впорядковано. При цьому сусідні ферменти здійснюють послідовно протікають реакції (робочі операції), а група їх - весь ланцюг реакцій, що ведуть до створення того чи іншого речовини.
Розрізняють гладкий (гладкий) і гранулярний ендоплазматичнийретикулум. На зовнішній поверхні каналів гранулярного ретикулума розташовуються численні дрібні органели - рибосоми, функцією яких є синтез білкових молекул. Гладкий ендоплазматичнийретикулум, який в рослинній клітині кількісно переважає над гранулярним, не несе рибосом.
Ендоплазматичнийретикулум, крім того, що він є конвеєром для багатьох видів ферментативного перетворення речовин, головним чином для їх синтезу, являє собою і систему магістралей, по яких речовини переміщаються по клітці. Починається ретикулум від зовнішньої мембрани оболонки ядра і, гілкуючись, підходить до різних органоидам цитоплазми, а також до плазмалемме. Тим самим він пов'язує між собою всі частини клітини. Крім того, канали ендоплазматичної проходять через плазмодесми, поєднуючи ретикулум сусідніх клітин.
Далі, мембрани ЕПР розчленовують цитоплазму на численні відсіки, завдяки чому клітку не можна уявити як однорідний масив, в якому перемішані самі різні речовини. Умови в одному відсіку можуть бути зовсім іншими, ніж в будь-якому іншому; процеси, в ньому протікають, можуть йти тільки в ньому, тоді як в кожному іншому відбуваються інші процеси.
Нарешті, мембрани ЕПР - це ті поверхні, по яких поширюються біоструми, які є сигналами, що міняють виборчу проникність мембран і тим самим активність ферментів. Завдяки цьому одні хімічні реакції пускаються в хід, інші гальмуються - обмін речовин підпорядковується регуляції і протікає координовано.
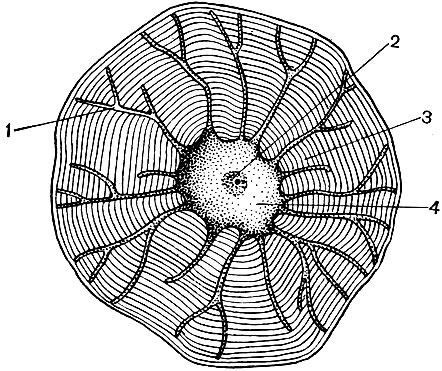
Багато з речовин, синтезованих в клітці, повинні бути сконцентровані і виділені із клітки або в зовнішню середу, або у внутрішньоклітинну вакуоль. Крім того, клітина концентрує і речовини, що надходять в неї з інших клітин, наприклад якщо вона відкладає їх про запас. Цю роботу виконують діктіосоми. Зазвичай в рослинній клітині є кілька диктиосом, і вся їх сукупність називається апаратом (або комплексом) Гольджі і даної клітини. Кожна діктіосома являє собою систему мембран, складених стопкою (табл. 10 - зріз, рис. 32 - об'ємна схема). Порожнини між мембранами, що утворять цей органоид, мають вигляд то вузьких щілин, то плоских мішечків - цистерн, то пухирців. Форма їх змінюється в ході роботи органоида і, мабуть, залежить від ступеня наповнення межмембранное просторів які виділяються і накопичується речовинами. Сформовані і розрослися бульбашки відокремлюються від органоида. Мабуть, багато клітинні вакуолі, оточені мембранами - тонопластом, є продуктом діяльності апарату Гольджі; це відірвалися від нього і потім збільшилися бульбашки.
Апарат Гольджі особливо розвинений у видільних (секреторних) клітках, в яких відкладаються або з яких виводяться різні речовини. Він синтезує і виділяє речовини, що утворюють клітинну оболонку.
Лізосоми - досить дрібні (близько 0,5 мк в діаметрі) округлі тільця - ще один органоид цитоплазми. Вони покриті оболонкою - липопротеиновой мембраною. Вміст лізосом - ферменти, що перетравлюють білки, вуглеводи, нуклеїнові кислоти і ліпіди. Оболонка лізосоми перешкоджає виходу ферментів з органоида в гіалоплазму, в іншому випадку остання переварювалася б цими ферментами.
Можна думати, що лізосоми є продуктами діяльності апарату Гольджі, що відірвалися від нього бульбашками, в яких цей органоид акумулював переварюють ферменти.
Ті частини клітини, які відмирають в процесі її розвитку, руйнуються за допомогою ферментів лізосом. У померлої клітці лізосоми руйнуються, ферменти виявляються в цитоплазмі, і вся клітина, за винятком оболонки, піддається переварюванню.
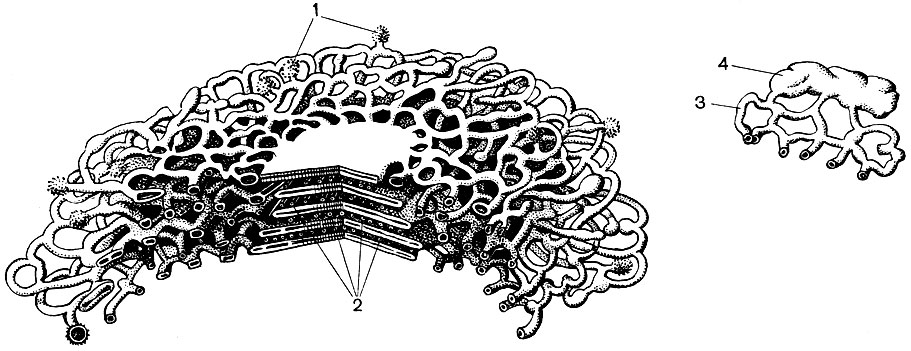
Рибосоми - дуже дрібні органели, діаметр їх близько 250А. За формою вони майже кулясті. Частина їх прикріплена до зовнішніх (гіалоплазматіческім) поверхнях мембран, що утворюють канали шорсткогоЕПР; частина ж знаходиться - у вільному стані в гіалоплазме. У клітці може міститися до 5 млн. Рибосом. Вони являють собою "апарати для синтезу білка. Тому особливо багато їх в клітинах, активно утворюють білок, - в зростаючих клітинах, в клітинах, які секретують білкові речовини. Рибосоми є також в мітохондріях і хлоропластах, де вони синтезують частина білків, у тому числі побудовано ці органели.
У багатьох клітинах виявлені органели, названі мікротрубочками. Саме їх назва говорить про їхню форму - це трубочки з каналом всередині. Зовнішній їх діаметр близько 250А. Іноді це подвійні трубочки - дві одиночні, що лежать пліч-о-пліч один з одним і мають загальну стінку, яка розділяє їх порожнини. Стінки микротрубочек побудовані з білкових молекул. Вважають, що мікротрубочки пов'язані з скорочувальної (рухової) активністю цитоплазми і її утворень. З них, як з будівельних деталей, побудовані, мабуть, скоротливі структури джгутика - органоида, за допомогою якого переміщаються деякі одноклітинні і колоніальні водорості, а також клітини, службовці для розмноження багатьох нижчих рослин. З мікротрубочок під час поділу клітини утворюються нитки веретена, про який мова буде йти далі. В період поділу мікротрубочки збираються в групи і утворюють ці нитки. Після закінчення поділу нитки знову розпадаються на окремі мікротрубочки. У клітинах або їх частинах, які позбавлені щільної оболонки, мікротрубочки, можливо, виконують опорну функцію, складаючи внутрішній скелет клітини.
Пластида - органели, властиві тільки рослинною клітинам. Зазвичай це великі тільця, що абсолютно очевидно під світловим мікроскопом.
Розрізняють 3 типи пластид: безбарвні - лейкопласти, зелені - хлоропласти, пофарбовані в інші кольори - хромопласти. Пластида кожного типу мають свою будову і несуть свої, їм властиві функції. Однак можливі переходи пластид з одного типу в інший. Так, позеленіння бульб картоплі викликається перебудовою їх лейкопластов в хлоропласти. У коренеплоді моркви лейкопласт переходять в хромопласти. Пластида всіх трьох типів утворюються з пропластид.
Пропластид - безбарвні тільця, схожі на мітохондрії, але трохи крупніше їх. У великих кількостях вони зустрічаються в меристематичних клітинах. Лейкопласти знаходяться в клітинах нефарбованих частин рослин (плодів, насіння, коріння, епідермісу листя). Форма їх невизначена. Найчастіше зустрічаються лейкопласт, в яких відкладається крохмаль (він утворюється з Сахаров). Є лейкопласт, що запасають білки. Найменш поширені лейкопласт, заповнені жиром; вони утворюються при старінні хлоропластів. Істотних відмінностей між лейкопластах і пропластид немає.
Хлоропласти - пластиди вищих рослин, в яких йде процес фотосинтезу, т. Е. Використання енергії світлових променів для утворення з неорганічних речовин (вуглекислого газу і води) органічних речовин з одночасним виділенням в атмосферу кисню хлоропласти мають форму двоопуклої лінзи, розмір їх близько 4 - 6 мк. Знаходяться вони в наренхімних клітинах листя та інших зелених частин вищих рослин. Число їх в клітці варіює в межах 25 - 30.
Про будову хлоропласта дають уявлення таблиці 11 і 12 (вид на зрізі) і схемою на малюнку 33. Зовні хлоропласт покритий оболонкою складається з двох ліпопротеїнових мембран. Під нею, в основному речовині (стромі), впорядковано розташовані численні освіти - ламелли. Вони утворюють плоскі мішечки, які лежать один на одному правильними стоїками. Ці стоїки, що нагадують монети, складені стовпчиком, називаються гранами. Крізь них проходять довші ламелли, так що все грани хлоропласта пов'язані в єдину систему. До складу мембран утворюють грани, входить зелений пігмент - хлорофіл. Саме тут відбуваються світлові реакції фотосинтезу - поглинання хлорофілом світлових променів і перетворення енергії світла в енергію збуджених електронів. Електрони, порушені світлом, т. Е. Мають надлишкової енергією віддають енергію на розкладання води і синтез АТФ. При розкладанні води утворюються кисень і водень. Кисень виділяється в атмосферу, а водень зв'язується білком ферредоксин. Ферредоксин потім знову окислюється, віддаючи цей водень речовини-відновники, скорочено позначається НАДФ. НАДФ переходить у відновлену форму - НАДФ-Н 2. Таким чином, підсумком світлових реакцій фотосинтезу є утворення АТФ, НАДФ-Н 2 і кисню, причому споживаються вода і енергія світла.
В АТФ акумулюється багато енергії - вона потім використовується для синтезів, а також для інших потреб клітини. НАДФ-Н 2 - акумулятор водню, причому легко його потім віддає. Отже, НАДФ-Н 2 є хімічним відновником. Велике число биосинтезов пов'язано саме з відновленням, і в якості постачальника водню в цих реакціях виступає НАДФ-Н 2.
Далі, за допомогою ферментів строми хлоропластів, т. Е. Поза гран, протікають темнова реакції: водень і енергія, укладена в АТФ, використовуються для відновлення атмосферного вуглекислого газу (С0 2) і включення його при цьому до складу органічних речовин. Перше органічна речовина, що утворюється в результаті фотосинтезу, піддається великому числу перебудов і дає початок всьому різноманіттю органічних речовин, що синтезуються в рослині і складових його тіло. Ряд з цих перетворень відбувається тут же, в стромі хлоропласта, де є ферменти для утворення Сахаров, жирів, а також все необхідне для синтезу білка. Сахара можуть потім або перейти з хлоропласта в інші структури клітини, а звідти в інші клітини рослини, або утворити крохмаль, зерна якого часто можна бачити в хлоропластах. Жири теж відкладаються в хлоропластах або у вигляді крапель, або у формі простіших речовин, попередників жирів, виходять з хлоропласта.
Ускладнення речовин пов'язане зі створенням нових хімічних зв'язків і звичайно вимагає витрат енергії. Джерело її - все той же фотосинтез. Справа в тому, що значна частка речовин, що утворюються в результаті фотосинтезу, знову розпадається в гіалоплазме і мітохондріях (в разі повного згоряння - до речовин, які служать вихідним матеріалом для фотосинтезу, - С0 2 і Н 2 0). В результаті цього процесу, по своїй суті зворотного фотосинтезу, енергія, раніше акумульована в хімічних зв'язках розкладаються речовин, звільняється і - знову за посередництвом АТФ - витрачається на утворення нових хімічних зв'язків синтезованих молекул. Таким чином, значна частина продукції фотосинтезу потрібна тільки для того, щоб зв'язати енергію світла і, перетворивши її в хімічну, використовувати для синтезу зовсім інших речовин.
І лише частина органічної речовини, що утворюється при фотосинтезі, використовується як будівельний матеріал для цих синтезів.
Продукція фотосинтезу (біомаса) колосальна. За рік на земній кулі вона складає 1010 т. Органічні речовини, створювані рослинами, - це єдине джерело життя не тільки рослин, але і тварин, так як останні переробляють вже готові органічні речовини, харчуючись або безпосередньо рослинами, або іншими тваринами, які, в свою чергу, харчуються рослинами. Таким чином, в основі всього сучасного життя на Землі лежить фотосинтез. Всі перетворення речовин і енергії в рослинах і тваринах є перебудови, перекомбінації і переноси речовини і енергії первинних продуктів фотосинтезу. Фотосинтез важливий для всього живого і тим, що одним з його продуктів є вільний кисень, що відбувається з молекули води і виділяється в атмосферу. Вважають, що весь кисень атмосфери утворився завдяки фотосинтезу. Він необхідний для дихання як рослинам, так і тваринам.
Хлоропласти здатні переміщатися по клітці. На слабкому світлі вони розташовуються під тією стінкою клітини, яка звернена до світла. При цьому вони звертаються до світла своєю більшою поверхнею. Якщо світло занадто інтенсивний, вони повертаються до нього ребром і вибудовуються уздовж відтінку, паралельних променів світла. При середніх освещенностях хлоропласти займають положення, середнє між двома крайніми. У будь-якому випадку досягається один результат: хлоропласти виявляються в найбільш сприятливих для фотосинтезу умовах освітлення. Такі переміщення хлоропластів (фототаксис) - це прояв одного з видів подразливості у рослин.
Хлоропласти мають відому автономією в системі клітини. У них є власні рибосоми і набір речовин, що визначають синтез ряду власних білків хлоропласта. Є також ферменти, робота яких призводить до утворення ліпідів, що входять до складу ламелл, і хлорофілу. Як ми бачили, хлоропласт розташовує і автономною системою добування енергії. Завдяки всьому цьому хлоропласти здатні самостійно будувати власні структури. Існує навіть погляд, що хлоропласти (як і мітохондрії) походять від якихось нижчих організмів, що мешкають в рослинній клітині і спершу вступили з нею в симбіоз, а потім стали її складовою частиною, органоїдом.
У нижчих рослин фотосинтез також здійснюється спеціалізованими, хоча і не настільки високорозвиненими, як в хлоропласті, мембранними структурами. У фотосинтезу-ючий бактерій мембрани, що містять хлорофіл, утворюють мережу, яка пронизує тіло бактерії. У синьо-зелених водоростей фотосинтезирующие мембрани злиті в плоскі пухирці. У зелених та інших водоростей система цих мембран відділена від іншої частини клітини покриває мембраною і утворює спеціальний органоид - хроматофор. Число хроматофорів в клітці невелика, часто клітина містить всього лише один хроматофор. Форма їх дуже різна у водоростей різних видів.
У спірогири хроматофор має вигляд стрічки, спірально в'ється уздовж стінок клітини; у клострідіум - це ребристі циліндри; у зігнеми - зірчасті тіла.
Хромопласти виникають або з пропластид, або з хлоропластів, або з лейкопластов. Їх внутрішня мембранна структура набагато простіше, ніж у хлоропластів. Гран немає, строма містить багато жовтого або оранжевого пігменту. Хромопласти містяться в клітинах пелюсток, плодів, коренеплодів.
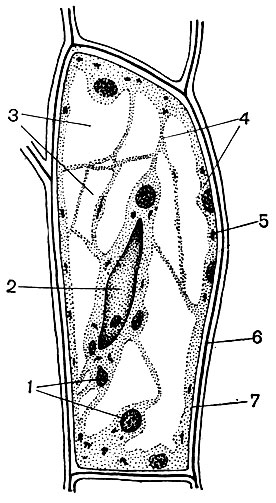
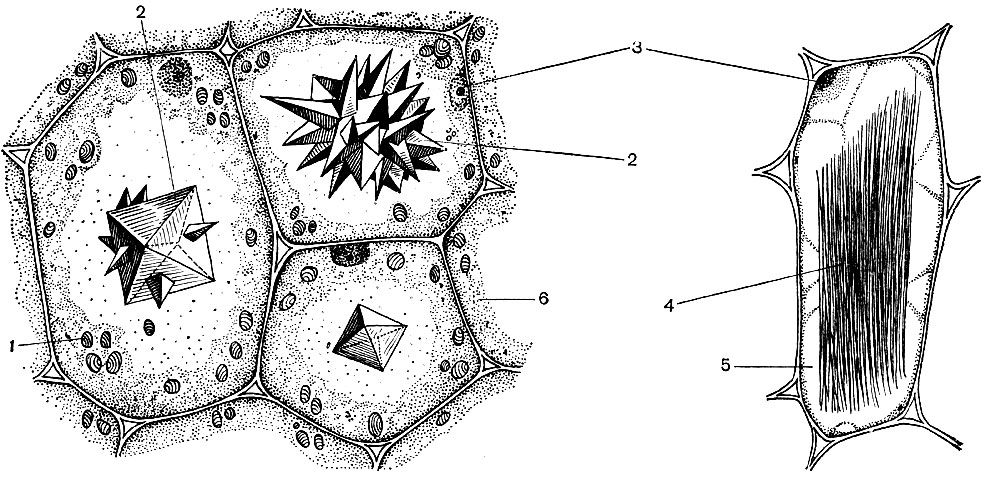
У типовій рослинній клітині є велика вакуоль, наповнена рідким вмістом. Часто вакуоль займає майже весь обсяг клітини, так що цитоплазма становить лише тонкий шар, що прилягає до клітинної оболонці. У молодих клітин буває кілька дрібних вакуолей, які в міру розвитку клітини розростаються і зливаються в одну. Вміст вакуолі - клітинний сік - це водний розчин дуже багатьох речовин: Сахаров, амінокислот, інших органічних кислот, пігментів (барвників), вітамінів, дубильних речовин, алкалоїдів, глікозидів, неорганічних солей (нітратів, фосфатів, хлоридів), іноді - білків.
Всі ці речовини - продукти життєдіяльності клітини. Одні з них зберігаються в вакуолярном (клітинному) соку в якості запасних речовин і з часом знову надходять в цитоплазму для використання. Інші є покидьками обміну речовин, виведеними геть з цитоплазми. Так, в вакуоль виводиться щавлева кислота; в вакуолярном соку часто відкладаються кристали щавлевокислого кальцію - іноді у формі одиночних кристалів, в інших випадках у вигляді конгломерату кристалів цієї солі - багатогранних (друзи) або голчастих (Рафіду), - зображених на малюнку 34.
У переважної більшості рослин (виняток становлять прокаріотів) в кожній живій клітині є ядро \u200b\u200bабо кілька ядер. Клітка, позбавлена \u200b\u200bядра, здатна жити лише короткий час. Без'ядерні клітини сітовідних трубок - живі клітини. Але живуть вони недовго. У всіх інших випадках без'ядерні клітини є мертвими.

Ядро завжди лежить в цитоплазмі. Форма ядра може бути різною - округлої, овальної, сильно витягнутої, неправильно-багатолопатеве. У деяких клітинах контури ядра змінюються в ході його функціонування, причому на його поверхні утворюються лопаті різної величини. Розміри ядер неоднакові і в клітинах різних рослин, і в різних клітинах одного і того ж рослини. Щодо великі ядра бувають у молодих, меристематичних клітинах, в яких вони можуть займати до 3/4 обсягу всієї клітки. Відносні, а іноді і абсолютні розміри ядер в розвинених клітинах значно менше, ніж в молодих.
Зовні ядро \u200b\u200bпокрите оболонкою, що складається з двох мембран, між якими є щілина - околоядерное простір. Оболонка переривається порами. Зовнішня з двох мембран оболонки дає вирости, безпосередньо переходять в стінки ендоплазматичноїмережі цитоплазми. І пори і прямий зв'язок ендоплазматичної мережі з околоядерних простором забезпечують тісний контакт між ядром і цитоплазмою.
Вміст ядра - зернисте основна речовина (ядерний сік, або нуклеоплазма), в якому містяться більш щільні структури - хромосоми і ядерце. Ядро являє собою апарат синтезу матеріалу рибосом і місце їх складання з цього матеріалу.
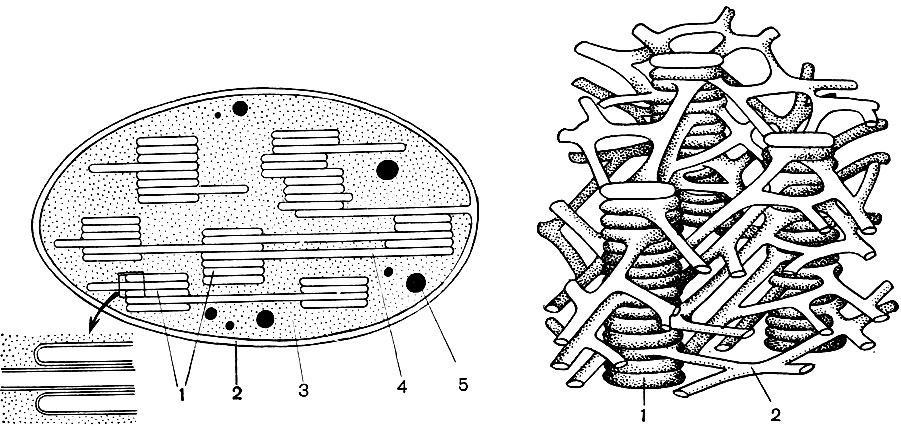
Хромосоми побудовані з великої кількості молекул дезоксирибонуклеїнової кислот (ДНК), з'єднаних з молекулами білків-гістонів. Молекули ДНК - це довгі складно упаковані подвійні нитки. Кожна молекула складається з двох ниток, переплетених спірально одна навколо іншої. Нитка, в свою чергу, - це ланцюжок з величезного числа так званих нуклеотидів. Нуклеотид - з'єднання азотистої основи, вуглеводу (дезоксирибози) і фосфорної кислоти.
До складу кожного з нуклеотидів входить одне з чотирьох наступних азотистих основ: аденін, гуанін, цитозин або тимін. Відповідно, в ДНК розрізняють 4 різних нуклеотиду: адениновую (А), гуанінових (Г), цитозинових (Ц) і тимінових (Т). Нуклеотиди з'єднані між собою через свої фосфатні групи, завдяки чому утворюється довгий ланцюжок. Два ланцюжки, закручені одна навколо іншої і утворюють одну молекулу ДНК, скріплені між собою хімічним взаємодією (так звані водневі зв'язку) азотистих основ своїх нуклеотидів. Підстави утворюють пари - одна підстава з одного ланцюжка, інше - з другої. Схема на малюнку 35 дає уявлення про порядок з'єднання нуклеотидів в ланцюжок, а двох ланцюжків - між собою.
Хоча всі молекули дезоксирибонуклеїнової кислот побудовані за описаним єдиним планом, конкретний їх якісний склад різний; відрізняються вони і за величиною молекул. Молекула ДНК містить до 50 - 100 тис, пар нуклеотидів, але число пар їх в кожній молекулі своє, відмінне від числа пар нуклеотидів в будь-який інший молекулі. Крім того, нуклеотидів існує 4 різних види, і в складі кожної молекули ДНК свій, їй властивий відсоток нуклеотидів кожного даного виду. Інакше кажучи, кількісне відношення А: Г: Ц: Т у кожної молекули ДНК своє. І нарешті, в кожній молекулі ДНК порядок чергування нуклеотидів А, Г, Ц, Т характерний тільки для даної молекули. Перестановка місцями тільки двох пар з 50 тис. Пар нуклеотидів, нехай навіть сусідніх, різко змінює властивості всієї молекули; те ж саме-заміна хоча б однієї пари, скажімо З - Г на А - Т або на Г - З; те ж саме - відсутність однієї пари з числа тих же 50 тис. Насправді молекули ДНК можуть відрізнятися між собою не в одній ланці, не однієї-двома парами нуклеотидів, а в величезному їх числі відразу. Кількість можливих перестановок пар нуклеотидів в молекулі ДНК нескінченно, і відповідно нескінченно кількість різних молекул, у кожної з яких свої властивості. Послідовність нуклеотидів в молекулі ДНК - це зашифрована запис складу того чи іншого білка, властивого даній клітині. Протягом кожної молекули ДНК послідовно вміщаються записи складу декількох білків, а у всіх молекулах ДНК всіх хромосом ядра - записи складу всіх білків клітини, які в ній можуть синтезуватися протягом її життя. Суть цих кодованих записів полягає в наступному.
Молекули кожного білка - це ланцюжки з послідовно з'єднаних амінокислот. Існує близько 20 різних амінокислот, і характер білка залежить від того, з яких саме амінокислот складається його молекула, яка їхня загальна кількість в молекулі і в якій послідовності вони з'єднані один з одним. Наприклад, ділянка ДНК, відповідальний за склад певного білка (кожна така ділянка називається геном), - це є запис конкретної послідовності амінокислот, що утворюють молекулу даного білка, їх загального числа в ній. Кожні 3 послідовних нуклеотиду ланцюжка ДНК позначають (кодують) одну амінокислоту відповідної білкової молекули. Наступні 3 нуклеотиду кодують наступну амінокислоту і т. Д. Кількості можливих відрізняються поєднань по 3 нуклеотиду з числа чотирьох різних видів їх з надлишком вистачає для закодування 20 амінокислот. Цікаво, що одні й ті ж амінокислоти в усьому живому світі кодуються одними й тими ж поєднаннями нуклеотидів. У підсумку в одному гені його нуклеотидним складом зашифрований амінокислотний склад всіх білків, здатних синтезуватися в даній клітці.
Клітинні білки є ферментами. Ферменти визначають протягом всіх реакцій, що становлять суть життєдіяльності клітин. Від наявності тих чи інших ферментів залежить освіту і перетворення всіх інших речовин клітини, будь то жири, вуглеводи, алкалоїди, смоли і т. Д. Це, в свою чергу, формує всі властивості клітини, що відрізняють її від клітин інших рослин, в клітинних ядрах яких закодований склад іншого набору ферментів. Ось чому відомості про склад всіх білків, які можуть утворитися в клітці, - це інформація про всі властивості клітини і організму. Далі ми побачимо, що ця інформація - спадкова, т. Е. Що вона в повному обсязі передається від клітини до клітини при їх розмноженні і від материнської рослини до дочірнім.
Зберігається ця інформація в хромосомах. Однак реалізація цієї інформації - синтез білків - відбувається не в них. Гени хромосом виступають тільки в ролі ініціаторів цього синтезу. Коли в клітці виникає потреба в освіті того чи іншого білка, то ген, у якому зашифровано склад цього білка, активується. Це означає, що на ділянці нитки ДНК, що становить, даний ген, утворюються молекули так званої інформаційної рибонуклеїнової кислоти (І-РНК).
Будова і склад цих молекул, що становлять одиночні ланцюжки з нуклеотидів, відображає нуклеотидное будова того гена, на якому вони утворилися. Таким чином відбувається копіювання інформації про склад майбутнього білка.
Освіта молекул і-РНК (на кожному гені їх утворюється багато) означає і розмноження інформації, як би віддруковування багатьох однакових матриць, що відображають будову одного і того ж гена і тим самим несуть в нуклеотидної послідовності своїх молекул інформацію про амінокислотним складом заданого білка. Ці відбитки гена переходять з ядра в цитоплазму. Тут відбувається розшифровка інформації, що містяться в матрицях - молекулах і-РНК, реалізація інформації, переведення її з мови нуклеотидноїпослідовності на мову послідовності амінокислот, т. Е. Синтез білкових молекул заданого складу.
У розшифровці і синтезі, крім молекули і-РНК, бере участь велика кількість молекул різних транспортних рибонуклеїнових кислот (т-РНК), рибосоми і ряд ферментів. Амінокислоти зв'язуються з т-РНК - молекула з молекулою. Кожному з 20 видів амінокислот відповідає своя т-РНК. Так, у молекули т-РНК є хімічні групи, здатні впізнавати свою амінокислоту, вибираючи саме її з усіх наявних амінокислот. Відбувається це за допомогою спеціальних ферментів. Дізнавшись свою амінокислоту, т-РНК вступає з нею в з'єднання. До початку молекули і-РНК приєднується рибосома, яка, просуваючись по і-РНК, з'єднує один з одним в поліпептидний ланцюжок саме ті амінокислоти, порядок яких зашифрований нуклеотидной послідовністю даної і-РНК. Це зчитування інформації, ця її розшифровка відбувається завдяки спеціальному механізму, наявного в транспортних РНК, молекули яких вибудовують один за одним саме ті амінокислоти, які "позначені", "записані" нуклеотидной послідовністю молекули і-РНК. Рибосома - апарат для хімічного зв'язування амінокислот у молекулу білка.
Отже, загальна схема така. На активованому гені - ділянці однієї з ланцюжків молекули ДНК - синтезуються нуклеотидні ж ланцюжка, молекули і-РНК, склад яких точно відображає склад гена, а значить, несе в собі запис складу білка, кодованого даним геном. У цитоплазмі рибосоми на основі інформації, перенесеної з гена молекулами і-РНК, за допомогою транспортних РНК з'єднують різні амінокислоти в заданій послідовності, в результаті чого утворюється молекула того білка, склад якого закодований в цьому гені.
Освіта молекул і-РНК на активованому гені служить і командою і конкретною програмою для синтезу строго певного білка. Одна і та ж молекула і-РНК використовується як креслення для створення багатьох однакових молекул білка. Однак вона досить недовговічна, тому для тривало триваючого синтезу нових молекул того ж білка необхідно освіту на одному і тому ж гені однакових, але нових екземплярів молекул і-РНК. З переходом гена в неактивний стан - а це відбувається при зникненні у клітини потреби в даному білку - він блокується, перестає утворювати і-РНК, і незабаром синтез цього білка припиняється. В ході життя клітини у неї виникають потреби в різних білках. Всякий раз активуються визначають їх гени і утворюються молекули і-РНК відповідного складу.
Кожна рибосома виробляє за своє життя багато молекул різних білків. Вона може працювати на основі будь-якої і-РНК, і результат, характер створеного нею білка залежить тільки від складу тієї і-РНК, в контакті з якої рибосома працювала на цей раз.
Таким чином, клітинне ядро \u200b\u200bвиконує наступні взаємопов'язані функції. У ньому зберігаються відомості про склад всіх білків, здатних синтезуватися в даній клітці протягом її життя. (Виняток становлять собою деякі білки мітохондрій і хлоропластів. Їх склад зашифрований у власних ДНК цих органоїдів, де ці ДНК і знаходяться. Тут же, на місці, відбуваються всі етапи розшифровки відомостей, укладених в цих ДНК, включаючи і сам синтез даних білків за допомогою власних рибосом.)
В ядрі зберігаються відомості про всі властивості клітини і організму. Ядро організовує синтез кожного з цих білків в потрібний момент. При поділів клітини, що супроводжується розподілом ядра, вся ця інформація в повному обсязі переходить в кожне з знову утворених ядер, в кожну нову клітку. Це можливо завдяки тому, що перед розподілом весь генний матеріал хромосом самоудваівается, утворюються два однакових його примірника і по одному з них виявляється в кожному з нових ядер. Ядро кожної клітини містить повний набір генів, властивих даному організму. Однак протягом життя різних спеціалізованих клітин працюють далеко не всі гени. В одних клітинах функціонує одна частина генів, в інших - інша, по-третє - третя. Саме тому клітини різних тканин одного організму відрізняються один від одного. Значна частина генів так і залишається в пасивному, лише "зберігає інформацію" стані від народження до самої смерті клітини, організму. Крім того, гени в даній клітині працюють не одночасно: одні активні в один період життя клітини, на такому-то етапі її розвитку; інші - на іншому і т. д.
Чому одні гени так і залишаються неактивними, яким чином включаються і вимикаються інші - дуже складна і важлива проблема, яка зараз інтенсивно досліджується. Велику роль у блокуванні і деблокування генів грають, мабуть, білки-гістони. Вони входять до складу хромосом, перебуваючи в з'єднанні з ДНК. Можливо, активація, "розкріпачення" гена відбувається тоді, коли молекула гистона від'єднується від відповідної ділянки ДНК, тим самим оголюючи його ланцюжка, дозволяючи їм розплестися і почати функціонувати хімічно. Приєднання гистона веде до блокування гена. Однак, чим керується приєднання і від'єднання гістонів, недостатньо ясно.
Генний матеріал, як говорилося, перебуває в хромосомах. Тому робота ядра зі зберігання спадкової інформації, по її подвоєння і передачі з клітки в клітку, по організації синтезу різних білків протягом життя клітини - це перш за все робота хромосом. Хоча вони існують протягом усього життя клітини, але у вигляді чітких структур хромосоми помітні в ядрі тільки під час поділу клітини. У цей час відбувається конденсація матеріалу хромосом і хромосоми вдається виявити методами мікроскопії як індивідуальні, чітко окреслені освіти.
Хромосоми мають різну форму. Це або прямі або вигнуті палички, або різноманітні гачки, овальні тільця, кульки (рис. 36). Сильно варіюють вони і за розмірами. Кожна клітина рослин даного виду містить в своєму ядрі однаковий набір (або набори) хромосом з чітко визначеного числа різних, але строго певних хромосом. У всіх вищих і деяких нижчих рослин протягом їх життєвого циклу чергуються два покоління: з клітинами, що містять в ядрах одноразовий (гаплоїдний, n) набір хромосом, і з клітинами, ядра яких мають подвійний (диплоїдний, 2 n) набір хромосом. Гаплоїдний набір складається з n різних хромосом, по одній кожного типу. Чисельна величина строго постійна для всіх гаплоїдних клітин всіх рослин даного виду. Так, для лісової суниці це число становить 7, для гороху - теж 7, але для квасолі - 11, для яблуні - 17, для земляної груші - 51, для цукрової тростини - 60. Диплоїдний набір - це два гаплоїдних, складених разом. У ньому по дві хромосоми кожного типу. Якщо у гороху в гаплоидном наборі 7 різних хромосом, то в диплоидном 7 різних пар хромосом, причому дві хромосоми в межах кожної пари однакові. Легко порахувати, що для перелічених рослин диплоїдний набір (2n) становить для суниці 14, для гороху - 14, для квасолі - 22, для яблуні - 34, для земляної груші - 102, а для цукрової тростини - 120 хромосом.
В життєвому циклі мохів, папоротей чергуються організми з диплоїдними і гаплоїдний клітинами. Ці організми у папоротей ведуть самостійну, ізольовану один від одного життя. У мохів диплоидное рослина живе на гаплоидном. У диплоидном організмі утворюються гаплоїдні клітини - спори. Кожна 1 з них, проростаючи, дає гаплоїдний організм, т. Е. Організм, побудований з гаплоїдних клітин. У ньому утворюються гаплоїдні же статеві клітини - гамети.
У момент запліднення чоловіча та жіноча гамети зливаються, утворюючи одну клітку - зиготу, що має одне ядро \u200b\u200b- результат злиття ядер обох гамет. Від кожної з гамет ядро \u200b\u200bзиготи отримує по гаплоидному набору хромосом, і в результаті воно має подвійний, диплоїдний набір їх. З зиготи розвивається організм, кожна клітина якого має диплоїдний набір хромосом.

У диплоїдний набір дві хромосоми кожної пари однакові за формою, внутрішньою будовою, містять гени, що керують появою однорідних ознак (рис. 36). Вони називаються гомологічними хромосомами. Одна з них відбувається з гаплоидного набору батьківської гамети, інша - материнської. Тому у раздельнополих організмів одна з них несе гени, що визначають розвиток підвідомчих їй ознак за батьківським типу, друга - по материнському.
Гомологічні хромосоми іншої пари таким же чином визначають розвиток іншого ряду ознак, третьої пари-третього ряду і т. Д. Хоча батьківський і материнський організми належать до одного виду, але спадкові властивості їх не тотожні - у них є і індивідуальні відмінності. Тому деякі гени однієї гомологічної хромосоми не тотожні відповідним генам другий.
Гаплоїдний набір, що входить до складу диплоидного і відбувається з батьківської гамети, несе батьківську спадковість з її індивідуальними рисами, а гаплоїдний набір з материнської гамети - материнську. Складна взаємодія однорідних, але не завжди тотожних генів двох гаплоїднийнаборів, в сумі утворюють один диплоїдний, визначає, які ознаки проявляться у диплоїдного потомства, яке, по суті, є гібридом батька і матері.
У голонасінних і покритонасінних рослин чергування поколінь відбувається в принципі, так само як у мохів і папоротей, але їх гаплоидная фаза сильно редукована і представлена \u200b\u200bчасто лише групою клітин. Вона живе не самостійно, а в тілі гаплоидного рослини. У покритонасінних рослин жіноче гаплоидное покоління укладено в зародковому мішку, що знаходиться в семяпочке, а чоловіче всередині пилкового зерна.
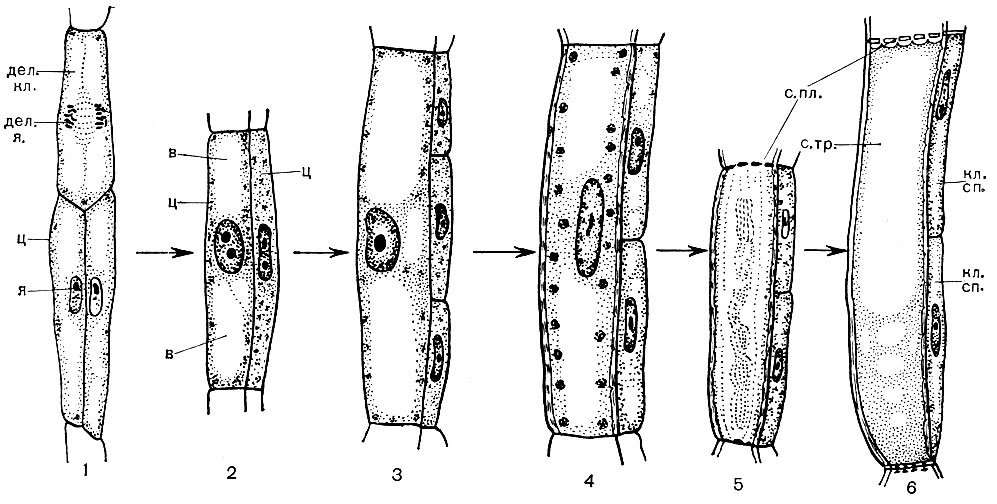
Ще до поділу клітини кожна молекула ДНК в кожній хромосомі прилаштовує біля себе свою копію - другу таку ж молекулу. В результаті весь спадковий матеріал клітини подвоюється, а кожна хромосома складається тепер з двох рівноцінних частин - хроматид. Далі "завдання" клітини полягає в тому, щоб, розділивши кожну хромосому на хроматиди, суворо порівну розподілити їх між майбутніми дочірніми клітинами: в кожну з них потрібно направити по одній хроматиді від кожної хромосоми. Це здійснюється наступним чином. Безпосередньо перед поділом клітини хромосоми сильно ущільнюються і скорочуються. Потім вони розташовуються в одній площині - по екватору ядра, причому одна хроматида кожної з них звернена до одного полюса клітини, інша - до протилежного. Ядерна оболонка зникає, розчиняється і ядерце. Між полюсами клітини з'являються нитки, у своїй сукупності утворюють фігуру веретена. Нитки збираються з з'єднуються один з одним микротрубочек. Веретено складається з ниток двох родів. Одні-безперервні, що йдуть від одного полюса клітини до іншого. Інші - тягнуть, кожна з яких з'єднує полюс з однією з хроматид. У хромосомі є ділянка - кінетохор, до якого і прикріплюються тягнуть нитки - одна від одного полюса, інша від другого. Далі відбувається розбіжність хроматид. Хроматиди, що складали до цього кожну хромосому, відокремлюються одна від одної і, підтягує нитками, розходяться до протилежних полюсів клітини. В результаті у кожного полюса збирається по одному повного набору хроматид, які тепер вже є хромосомами. Після цього нитки веретена розпадаються, навколо кожного набору хромосом утворюється ядерна оболонка, хромосоми сильно розбухають (деспирализуются). У кожному ядрі з'являється ядерце. Ядро знаходить структуру, властиву ядрам, неделящихся клітин. У серединній площині клітини утворюється перегородка, що розділяє клітку на дві дочірні.
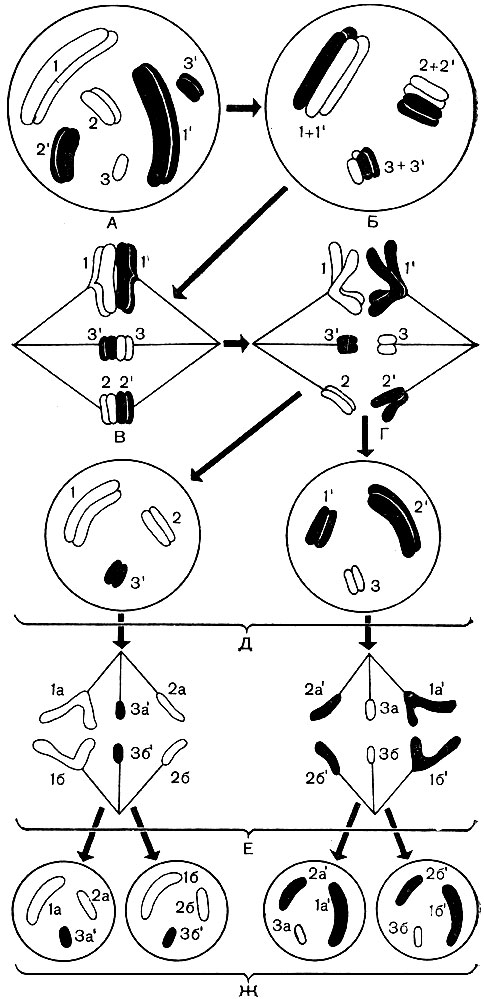
Рис. 38. Схема поведінки хромосом "умовної" клітини при мейозі. А - зображено диплоидное ядро \u200b\u200bклітини, що містить 3 пари хромосом (1 і 1 "- 1-я пара гомологічних хромосом, 2 і 2" - 2-я пара, 3 і 3 "- 3-я пара). 2n \u003d 6. 3 хромосоми, що походять від материнського організму, - світлі (цифри без штриха); парні їм гомологічні хромосоми, що походять від батьківського організму, зачорнені (цифри зі штрихом). гомологічні хромосоми позначені загальним номером. Кожна хромосома складається з двох хроматид. Б - злиття гомологічниххромосом . На цій стадії гомологічні хромосоми обмінюються між собою окремими ділянками. У результаті відбувається певний перерозподіл материнського і батьківського спадкового матеріалу між хромосомами (кросинговер). В - утворюються нитки веретена, що прикріплюються до хромосом, зникає оболонка ядра. Г - гомологічні хромосоми розходяться до протилежних полюсів клітини, у полюсів виявляється по одній гомологичной хромосомі з кожної пари, загальне число хромосом у кожного полюса вдвічі менше, ніж у вихідному ядрі А. Д - образ уются два ядра з гаплоїдний набором хромосом в кожному; в одне з них потрапило більше бабусиних (світлих, материнських по відношенню до ядра А) і менше дідових (зачерненого, батьківських по відношенню до ядра А) хромосом, в інше - навпаки; таким чином, нові ядра не цілком тотожні один одному за складом свого спадкового речовини; їх відмінність обумовлена \u200b\u200bтакож і кросинговером, що відбувається з хромосомами на стадії Б. Е - Ж - мітотичний поділ кожного з гаплоїдних ядер Д: поздовжнє розщеплення кожної хромосоми, розбіжність хромосом до полюсів, утворення двох гаплоїдних ядер з кожного ядра Д. В результаті з'явилося 4 клітини з гаплоїдними ядрами замість однієї клітини з диплоїдним набором хромосом
Органели розподіляються між дочірніми клітинами не строго порівну, але потім в клітинах синтезуються їх складові частини, відбувається складання нових екземплярів органоидов кожного виду, і число їх в кожній клітині відновлюється до нормального. Самі клітини ростуть. У хромосомах відбувається подвоєння спадкового матеріалу, після чого хромосоми складаються, як перед поділом клітини, з двох хроматид. Клітка готова до нового поділу.
Рис. 38А. Схема кросинговеру під час мейозу, що відбувається на стадії Б рис. 38. 1 - дві гомологічні хромосоми, кожна з яких складається з двох сестринських хроматид; 2 - гомологічні хромосоми сплелися один з одним; 3 - кожна хромосома розщеплюється на складові її хроматиди; хромосоми починають відштовхуватися один від одного і розходитися, але в деяких точках (на малюнку - в чотирьох) вони залишаються пов'язаними; в центральній петлі здвоєні сестринські хроматиди, а в обох крайніх петлях - несестрінскіе; точки, де можливий кросинговер, показані стрілками; 4 - хроматиди обмінялися ділянками, і хромосоми, зазнавши як би взаємну гібридизацію, розходяться; 5 - дві розійшлися, але вже "гібридні" (на відміну від показаних на 1) хромосоми
При зміні диплоидного покоління клітин гаплоидним відбувається так зване редукційний розподіл ядра - мейоз. Під час мейозу (рис. 38) гомологічні хромосоми кожної пари зближуються, тісно прилягають один до одного по своїй довжині, перекручуються. Між дотичними гомологічними хромосомами відбувається обмін окремими ділянками. В результаті цього частина генів батьківських хромосом переходить до складу материнських хромосом, а відповідні їм гени материнських хромосом займають місця, що звільнилися в батьківських хромосомах (явище кросинговеру - рис. 38А). Зовнішній вигляд тих і інших хромосом в результаті цього не змінюється, але їх якісний склад стає іншим. Батьківська і материнська спадковості перерозподіляються і змішуються. Далі ядерна оболонка і ядерце розчиняються, утворюється апарат веретена, такий же, як при мітозі. Гомологічні хромосоми роз'єднуються і за допомогою ниток веретена розходяться до полюсів клітини. У одного полюса виявляється один гаплоїдний набір хромосом (по одній гомологичной хромосомі з кожної пари), в іншого - другий гаплоїдний набір.
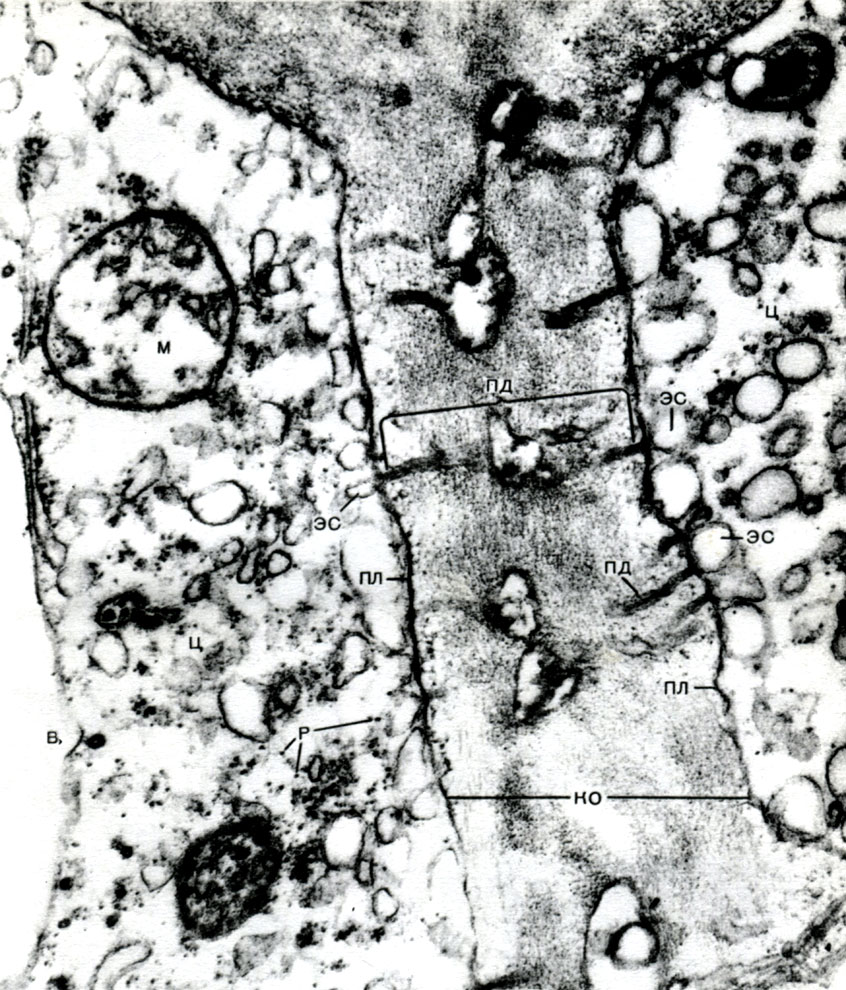
Слідом за мейозом утворилися гаплоїдні ядра діляться шляхом мітозу. При цьому кожна хромосома гаплоїдного набору розщеплюється на дві хроматиди, вони розходяться і утворюються дочірні гаплоїдні клітини.
Мейоз відрізняється від мітозу двома принциповими моментами. По-перше, при мейозі відбуваються злипання кожної пари гомологічних хромосом і обмін між ними частиною спадкового матеріалу, чого немає при мітозі. По-друге, при мейозі до полюсів клітини розходяться гомологічні хромосоми, по одній від кожної пари, а при мітозі кожна хромосома розщеплюється на хроматиди і до полюсів розходяться хроматиди, по одній від кожної хромосоми. В результаті мейозу з диплоидной клітини утворюються гаплоїдні.
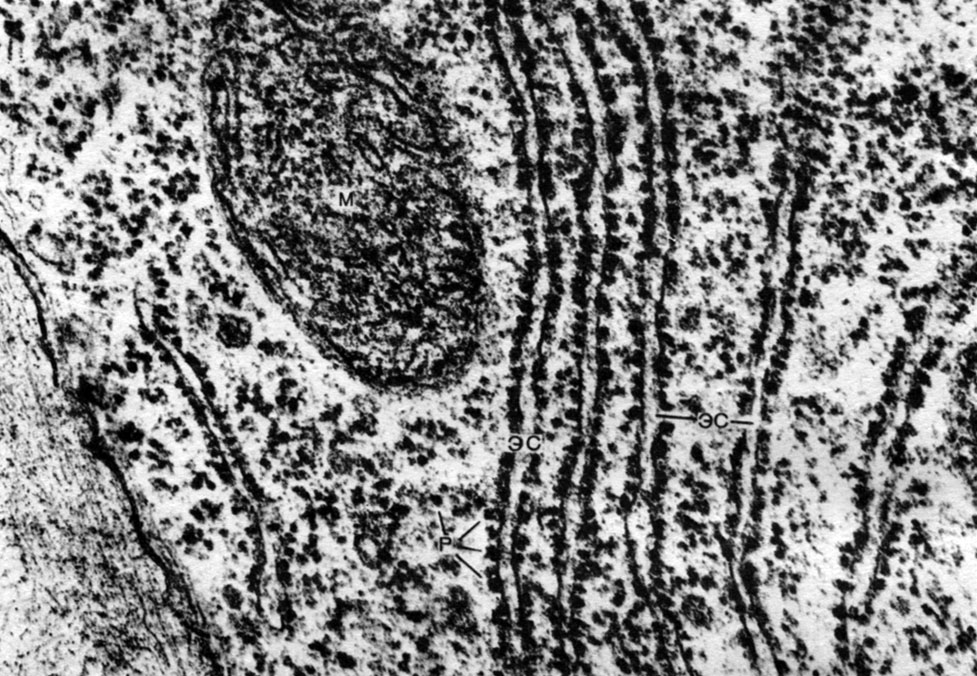
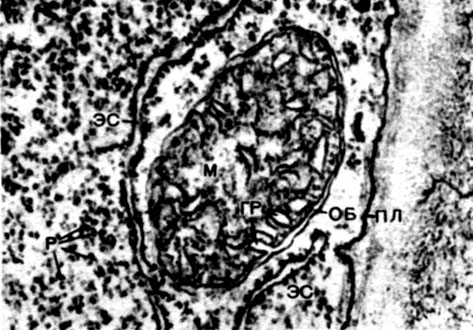
Таблиця 10. Структура рослинної клітини. Апарат Гольджі в цитоплазмі розвивається кореневого волоска редису (Raphanus sativus). Електронна мікрофотографія (повів. Х 52000) М. Ф. Данилової: аг - апарат Гольджі, видно зрізи плоских мішечків і бульбашок
У клітинах деяких тканин при їх розвитку відбувається незавершений мітоз: матеріал хромосом в ядрах подвоюється, хромосоми діляться навпіл, але, замість того щоб утворити два ядра, залишаються в початковому ядрі. З цього моменту воно укладає в собі не диплоїдний, а тетраплоїдний (четверний) набір хромосом. Процес, що веде до подібного подвоєння хромосом всередині одного ядра, називається Ендомітоз - внутрішнім митозом. Якщо він відбувається з одним ядром двічі, то воно стає октоплоідним (восьмиразовим) набором і т. Д. Клітини, ядра яких несуть в собі більше двох наборів хромосом, називаються поліплоїдні, т. Е. Многоплоіднимі. Полиплоидия клітин в ряді випадків підвищує їх життєздатність, оскільки кожен ген дублюється кількома іншими такими ж генами. Однокачественностью гени діють в унісон, і пошкодження якогось із них не веде до випадання визначається їм ознаки, так як компенсується роботою інших однорідних генів. У багатьох випадках поліплоїдні клітини більші і багатше вмістом, ніж диплоїдні. Виведено сорти полиплоидних культурних рослин, які мають підвищені господарськими якостями.
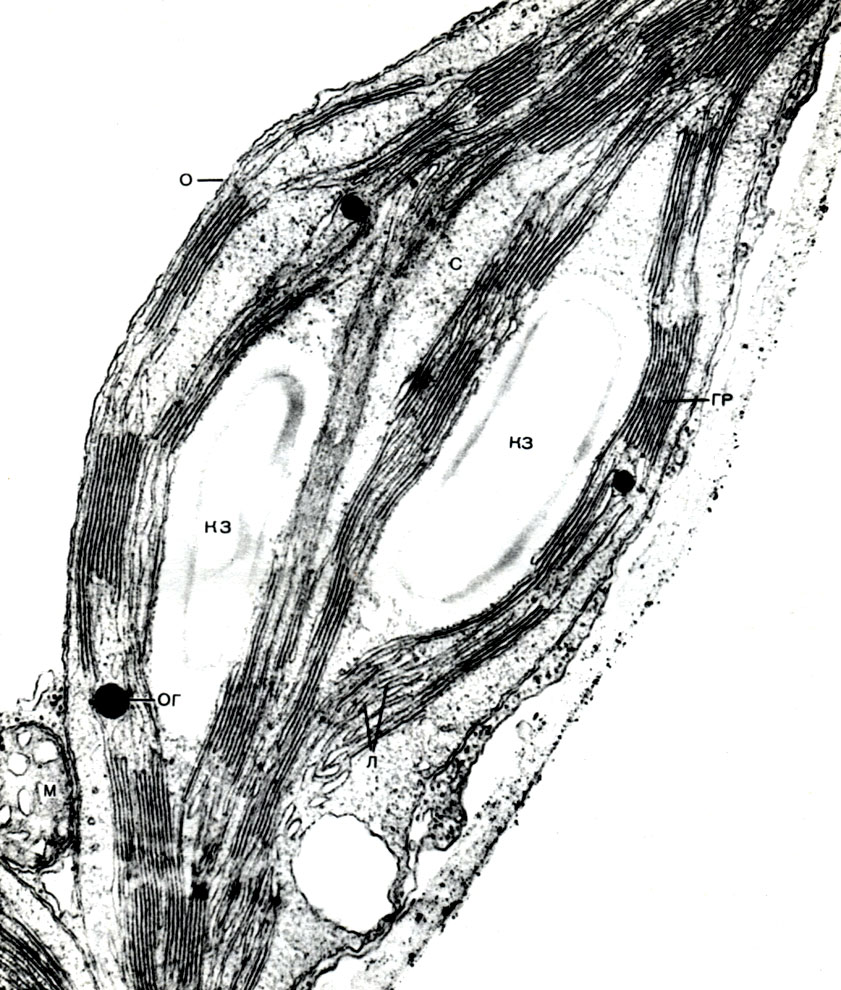
Наука, що вивчає клітку, називається цитологією. Клітка являє собою основу будови всього живого і всіх життєвих процесів; тому більшість найважливіших общебиологических проблем можна вирішити із залученням цитології і методів її дослідження.
За допомогою цитології вирішуються багато практично важливі питання: боротьба з хворобами рослин (грибними, бактеріальними), виведення нових сортів культурних рослин, подолання стерильності (безпліддя) гібридних сортів.
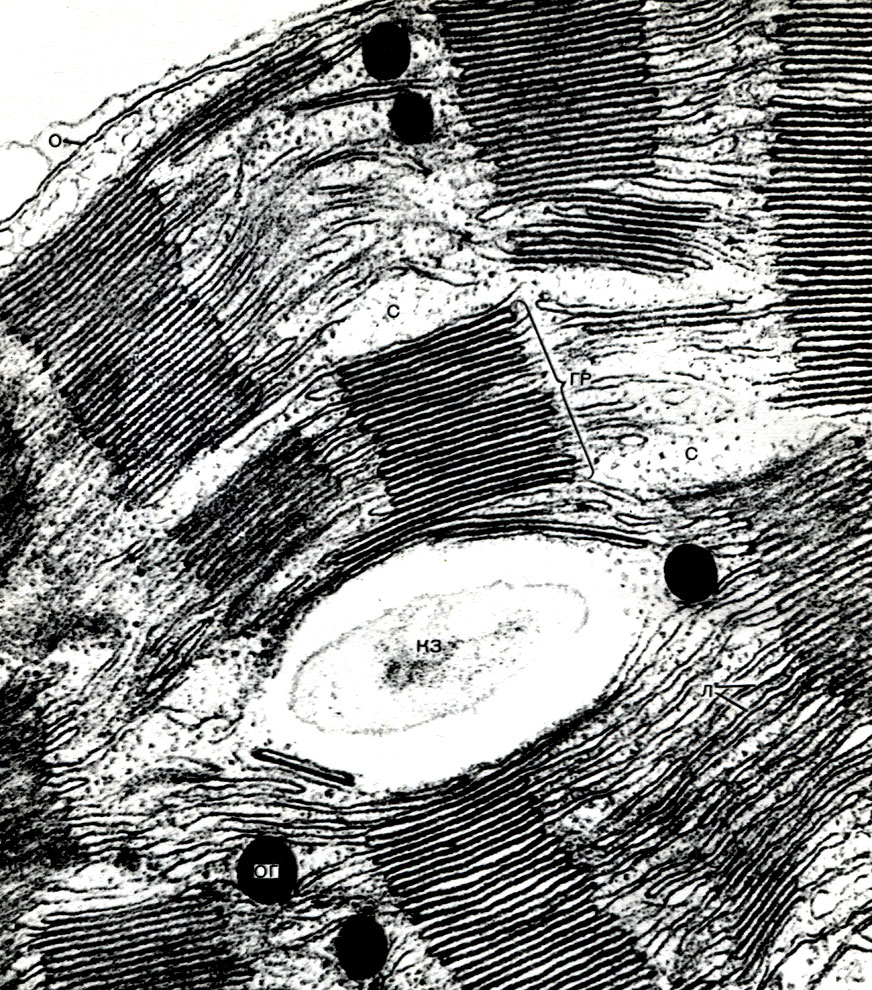
Ці проблеми сучасна цитологія досліджує, застосовуючи різноманітні методи - мікроскопічні, біохімічні, біофізичні, генетичні - і тісно взаємодіючи з іншими суміжними біологічними науками.