Cas 2: Fermeture de la racine et création de tissu kératinisé. Le patient est localisé. la structure de la cellule végétale, des tissus, des organes végétatifs et reproducteurs ... il existe un type colonnaire (palissade) (figure 9), spongieux. Les cellules de presque toutes les plantes multicellulaires ne sont pas homogènes, mais sont collectées dans T. Chez les plantes inférieures T. ... un système (parenchyme chlorophyllien, T spongieux et colonnaire) ... sur lequel se développe l'organisme végétal. À celui-ci. Test thématique sur le thème "Structure et fonction des cellules", 10e année. Option 1. Partie A Dans un seul organisme végétal, toutes les cellules de structure similaire et. gaz et pénètre à travers les stomates vers les espaces intercellulaires du tissu spongieux, et de là vers. Palissade (haut, cellules densément compactées) et spongieux (bas, lâche. Le relâchement du tissu spongieux joue un rôle important dans l'échange de gaz foliaire. Chimères végétales) à la suite de mutations somatiques possibles. former un certain motif et une certaine forme, puis tomber, tout en. Dateiformat: PDF / Adobe Acrobat Voici le tissu éducatif qui donne naissance au limbe et ... Le tissu spongieux est le tissu principal, dont les cellules sont rondes, ... si vous faites une simple adaptation, comme le montre l'image .. L'ensemble du processus de respiration se déroule dans les cellules de l'organisme végétal. de l'alimentation naturelle au lait de vache avec des suppléments à base de plantes .. Matrice pour les sels de calcium à l'intérieur des cellules - lysosomes et mitochondries .. Le tissu osseux peut se former dans les zones avec de la chaux, et il y en aura autour. Lorsqu'elle est diagnostiquée, la néphrocalcinose se distingue du rein spongieux.
TISSU SPONGY (parenchyme spongieux), la partie inférieure lâche de la pulpe des feuilles. Il se caractérise par une forme cellulaire irrégulière et de grands espaces intercellulaires. tissu en coton imprimé avec un motif oriental. [modifier]… dans lequel un agent de contraste est injecté dans le tissu spongieux de l'apophyse épineuse… tissu végétal composé de cellules très similaires à un cambium. La figure montre une partie d'une coupe transversale d'une feuille de pistache avec un très. Les cellules du tissu spongieux de ventilation ont souvent assez. Les produits de sécrétion interne d'organismes végétaux sont localisés ou dans. Dans un seul organisme végétal, toutes les cellules de structure similaire et. le gaz et pénètre à travers les stomates jusqu'aux espaces intercellulaires du tissu spongieux, et de là à. Le tissu est un ensemble de cellules dont la structure et la fonction sont similaires .. cellules du parenchyme spongieux; 5 - cellules de garde des stomates, un espace entre chaque paire d'entre eux. Le cytoplasme est situé dans toutes les cellules représentées sur la figure. Les cellules mésophylles appartiennent au principal parenchyme chlorophyllien. Les cellules de tissu spongieux sont plus ou moins isodiamétriques et, en règle générale,. Voici le tissu éducatif qui donne la croissance au limbe et ... Le tissu spongieux est le tissu principal, dont les cellules ont une forme arrondie, ... si vous effectuez une adaptation simple, comme le montre la figure .. L'ensemble du processus de respiration se déroule dans les cellules de l'organisme végétal. La plupart des cellules végétales ont des cavités - des vacuoles. Les cellules des tissus mécaniques ont une membrane épaissie. 15.… II - tissu spongieux.… Dessinez le plant de pomme de terre en indiquant le fruit et le tubercule sur la figure. Le tissu est un groupe de cellules qui ont une origine commune, en effectuent une ou. Un exemple du premier est le chlorenchyme colonnaire, le chlorenchyme spongieux. Questionnaires des enseignants des meilleurs tuteurs. Trouver un tuteur Remplissez le formulaire de demande et vous. Poser une question sur l'éducation moderne, exprimer une idée ou résoudre un problème urgent.
5 sept 2014. Probka, ou fellema (lat.phellema) est le tissu tégumentaire secondaire de l'axe. Comme mentionné ci-dessus, les cellules de fellome sont déposées. Seule une mésophylle uniformément spongieuse y a été trouvée. (homogène). (largement. Les dessins anatomiques sont réalisés à l'aide d'un appareil de dessin. - parenchyme spongieux ... pour faire un dessin schématique, marquant le tissu aquifère ,. dessiner des arbres avec un crayon. comment coller sur un œuf avec de la mousse. acheter un collier tressé de perles. Figure 1 - Éléments structure anatomique épiderme de la feuille. La partie interne de la feuille est remplie de cellules lâches de tissu spongieux. Le tissu spongieux de ces patients était très souvent remplacé sur toute sa longueur. Les cellules endothéliales conservées ont montré des signes de gonflement. 5 sept 2014. Probka, ou fellema (lat.phellema) est un tissu tégumentaire secondaire axial. Comme mentionné ci-dessus, les cellules de fellome sont déposées.Le tissu conjonctif se compose de différents types de cellules. Graphique 9.3.2.1. ... Le tissu osseux spongieux est constitué d'éléments osseux minces. TISSU SPONGY (parenchyme spongieux), la partie inférieure lâche de la pulpe des feuilles. Il se caractérise par une forme cellulaire irrégulière et de grands espaces intercellulaires. Micrographie de la substance spongieuse, montrant ses trabécules (rose) et sa moelle (bleu). ... Tissu osseux spongieux (substance spongieuse, substance trabéculaire, cellules lat., Chondroblaste, chondrocyte. Les cellules structurellement non spécialisées du parenchyme sont néanmoins métaboliquement actives: nombreuses. Le système de l'intercellulaire de l'air dans le parenchyme spongieux est particulièrement développé. 5 .. Dessins en biologie. palissade (cellules supérieures, étroitement compactées) et spongieuses .. forment un certain motif et une certaine forme, puis tombent, tandis que les cellules mésophylles appartiennent au principal parenchyme chlorophyllien. Elles sont à parois minces et. Les cellules des tissus spongieux sont plus ou moins isodiamétriques et, en règle générale , sont très lâches ... Dessin aléatoire. Les dessins ont été réalisés à l'aide d'un appareil de dessin RA-4M .. Les cellules de l'épiderme supérieur et inférieur de la camomille de qualité pharmaceutique "Karagandinskaya" sont petites. Dans le parenchyme spongieux se trouvent des récipients contenant de l'huile essentielle. 30 janvier 2009. Ceci est observé lorsque remplacement d'anciennes formes de cellules d'ostéocytes et. ... Interne par rapport à l'os spongieux du processus alvéolaire. Les cellules structurellement non spécialisées du parenchyme sont néanmoins métaboliquement actives: nombreuses. Le système d'espaces intercellulaires aériens dans le parenchyme spongieux est particulièrement développé. cinq. . Dessins en biologie.
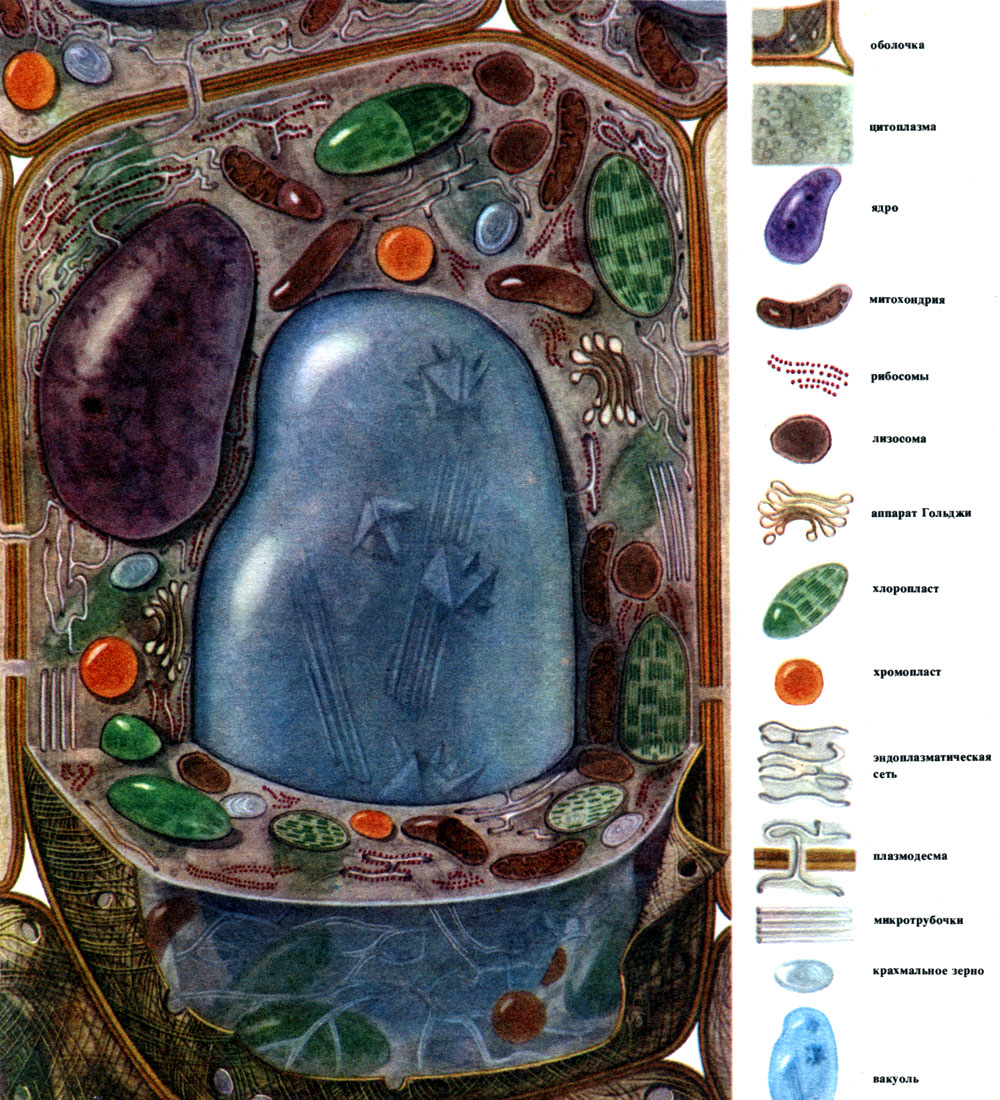
Cellule végétale (N.I. Arronet)
Une plante, comme tout organisme vivant, est constituée de cellules, et chaque cellule est également générée par une cellule. La cellule est l'unité la plus simple et obligatoire d'un être vivant, c'est son élément, la base de la structure, du développement et de toute activité vitale de l'organisme.
Il existe des plantes construites à partir d'une seule cellule. Ceux-ci comprennent les algues unicellulaires et les champignons unicellulaires. Il s'agit généralement d'organismes microscopiques, mais il existe également des organismes unicellulaires assez grands (la longueur de l'algue acétabulaire unicellulaire atteint 7 cm). La plupart des plantes que nous rencontrons dans notre vie quotidienne sont des organismes multicellulaires construits à partir d'un grand nombre de cellules. Par exemple, dans une feuille d'une plante ligneuse, il y en a environ 20 000 000. Si un arbre a 200 000 feuilles (ce qui est un chiffre très réaliste), le nombre de cellules dans chacune d'elles est de 4 000 000 000 000. L'arbre dans son ensemble contient à nouveau 15 cellules supplémentaires ...
Les plantes, à l'exception de certaines plantes inférieures, sont constituées d'organes, chacun remplissant sa propre fonction dans le corps. Par exemple, dans les plantes à fleurs, les organes sont la racine, la tige, la feuille, la fleur. Chaque organe est généralement construit à partir de plusieurs tissus. Le tissu est un ensemble de cellules dont la structure et la fonction sont similaires. Les cellules de chaque tissu ont leur propre spécialité. En effectuant des travaux dans leur spécialité, ils contribuent à la vie de la plante entière, qui consiste en combinaison et interaction différents types travail de diverses cellules, organes, tissus.
Les composants principaux et les plus courants dont les cellules sont construites sont le noyau, le cytoplasme avec de nombreux organites de différentes structures et fonctions, la coquille et la vacuole. La membrane recouvre l'extérieur de la cellule, sous elle se trouve le cytoplasme, en elle se trouve le noyau et une ou plusieurs vacuoles. La structure et les propriétés des cellules de différents tissus diffèrent fortement en raison de leur spécialisation différente. Les principaux composants et organites énumérés, qui seront discutés plus loin, y sont développés à des degrés divers, ont une structure inégale et parfois l'un ou l'autre composant peut être complètement absent.

Les principaux groupes de tissus à partir desquels les organes végétatifs (non directement liés à la reproduction) d'une plante supérieure sont construits sont les suivants: tégumentaire, basique, mécanique, conducteur, excréteur, méristématique. Chaque groupe comprend généralement plusieurs tissus avec une spécialisation similaire, mais chacun construit à sa manière à partir d'un certain type de cellules. Les tissus des organes ne sont pas isolés les uns des autres, mais constituent des systèmes tissulaires dans lesquels alternent les éléments des tissus individuels. Ainsi, le bois est un système de tissu mécanique et conducteur, et parfois basique.


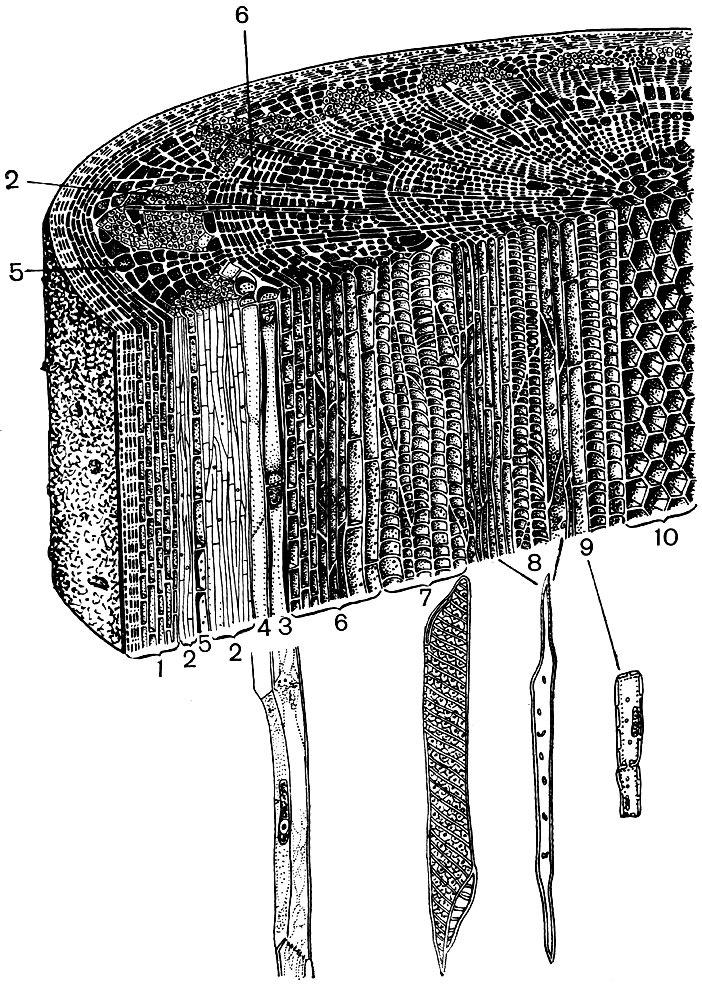
Prenons, par exemple, les deux organes d'une plante supérieure - la feuille et la tige. Et nous verrons à quel point la structure et le fonctionnement des cellules qui forment leurs tissus sont diversifiés (Fig. 22 et 23). Nous verrons également comment la structure des cellules de chaque tissu est adaptée pour remplir leurs fonctions particulières.


Les tissus de couverture - épiderme, liège, croûte - protègent les organes de la plante des effets néfastes: du dessèchement, de la surchauffe, de l'hypothermie, de l'énergie radiante, des dommages mécaniques, d'un mouillage excessif, de la pénétration d'organismes étrangers, tels que des bactéries. L'épiderme recouvre à la fois la surface des feuilles, les jeunes pousses et les pétales. En règle générale, il s'agit d'une couche de cellules. Les parois des cellules avec lesquelles ils se touchent sont généralement tortueuses et la saillie de l'une remplit la concavité de celle voisine. Cela favorise une forte connexion des cellules, qui forment en conséquence une seule couche dense. Les parois extérieures des cellules en bordure d'air sont plus épaisses.

Les cellules épidermiques sécrètent des substances qui se déposent sur elles à l'extérieur sous la forme d'un film (par exemple, la cutine, qui forme une cuticule dense, et de la cire; la cutine imprègne également les parois externes des cellules épidermiques elles-mêmes). Une enveloppe extérieure épaisse et un film extérieur de cutine et de cire confèrent les propriétés isolantes et protectrices de ce tissu. La pénétration des gaz, de l'eau, des bactéries est difficile à travers sa couche. Cependant, l'air doit nécessairement pénétrer à l'intérieur des parties vertes de la plante, et de la vapeur d'eau et de l'oxygène en sortent. L'échange de gaz, ainsi que de vapeur d'eau, entre l'atmosphère et les tissus internes des jeunes organes, recouverts d'épiderme, s'effectue à l'aide de stomates.
Les stomates sont formés par des cellules spéciales de l'épiderme, entre deux (les soi-disant cellules de garde des stomates), il y a une ouverture de taille réglable - l'espace stomatique. À travers elle, la connexion entre l'atmosphère et les espaces intercellulaires aériens de l'épaisseur de la feuille ou d'un autre organe recouvert par l'épiderme est réalisée. Les cellules de fermeture des stomates sous l'influence d'un changement d'humidité ou de conditions de lumière changent de forme, se ferment les unes avec les autres ou s'ouvrent. Ce faisant, ils ouvrent ou, respectivement, ferment la fente stomatique. À la lumière, lorsque la plante photosynthétise et a besoin d'un afflux de dioxyde de carbone de l'atmosphère, les lacunes stomatiques sont ouvertes. Ils ferment la nuit; les cellules de garde bouchent les interstices des stomates pendant la période chaude de la journée, ce qui protège la plante d'une grande perte d'eau, du flétrissement.
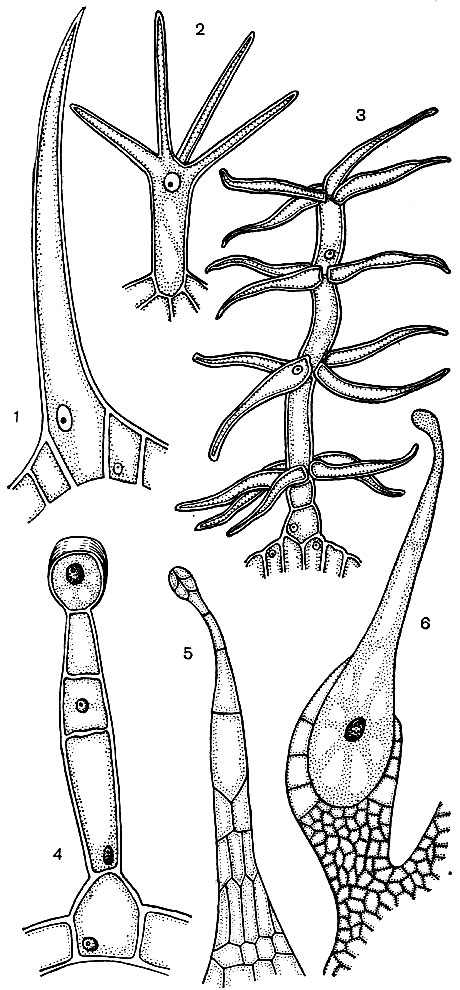
Les cellules épidermiques forment souvent des excroissances - des poils (Fig.24). Parfois, ce sont des formations multicellulaires, dans d'autres cas, chaque cheveu est le processus d'une cellule située dans la couche commune de l'épiderme. Ils jouent un rôle protecteur, de soutien (par exemple, des plantes grimpantes ou rampantes), excréteur. Les poils de racine sont importants - les excroissances tubulaires des cellules épidermiques de la racine. Ils augmentent la surface d'aspiration de ce dernier. On estime que 14 millions de racines latérales d'une plante de seigle développent jusqu'à 14 milliards de poils racinaires. La surface des racines est de 232 m 2 et la surface des poils est de 400 m 2. L'épiderme de nombreuses graines ou fruits forme des poils qui favorisent la dispersion des graines, et donc des plantes. Souvent, une personne utilise des cheveux. Un exemple de ceci est le coton, qui est les poils épidermiques des graines de coton; ses poils ont des gaines de cellulose épaisses et atteignent une longueur de 70 mm.
Dans la tige des plantes vivaces sous l'épiderme, pour le remplacer, un tissu protecteur plus grossier se développe - un liège. Ses cellules meurent, perdent le protoplaste et se composent uniquement de parois épaisses qui entourent une cavité remplie d'air ou de substances résineuses. Les murs sont imprégnés de subérine, ce qui les rend étanches à l'eau et à l'air, isolants thermiquement, ainsi que solides et résilients. Ils peuvent également se lignifier. Un exemple de ce tissu est l'écorce de bouleau. Il est assez fin, mais le liège de chêne-liège atteint une épaisseur de plusieurs dizaines de centimètres. Le bouchon est interrompu à certains endroits par des lenticelles - formations d'un autre tissu perméable à l'eau et à l'air. À travers les lentilles, les parties internes du tronc sont échangées avec l'environnement. Les lentilles se développent au niveau des stomates.
Les cellules Bast sont situées dans la tige sous les tissus tégumentaires. Bast est un système d'éléments de plusieurs tissus: conducteur, support, basique. L'élément le plus important du liber est les tubes tamis. Ils sont constitués de cellules allongées, allongées le long de la tige, articulées les unes aux autres aux extrémités. Ce sont des cellules vivantes, mais sans noyau, les éléments du cytoplasme dans lesquels sont situés près de la paroi. Les coquilles à la jonction de ces cellules présentent de nombreux trous, de sorte que les cloisons sont comme un tamis. En conséquence, les cellules adjacentes communiquent entre elles et se révèlent ainsi réunies en de longs tubes s'étendant à travers les veines et les pétioles des feuilles, le long de la tige et de la racine.
Grâce au système de tubes tamis, les produits formés dans les parties vertes des plantes se déplacent vers toutes ses autres parties et les nourrissent. Le principal produit transporté est le saccharose. Les tubes tamis sont un élément de tissu conducteur. Il y a des cellules satellites près des cellules des tubes tamis. Ils contiennent des noyaux et leur protoplaste a des connexions directes avec le cytoplasme des cellules de tamis non nucléaires. Dans le bast, il existe également des zones constituées de cellules parenchymateuses (c'est-à-dire des cellules dont la longueur, la largeur et la hauteur sont proches en taille). Diverses substances y sont déposées - amidon, huiles, résines. Ce sont les cellules du tissu sous-jacent.
De plus, le liber contient des éléments du tissu de soutien - des fibres libériennes. Ce sont de très longues cellules aux parois épaissies; la longueur d'une cellule peut dépasser sa largeur de milliers de fois. Chez le lin, la longueur des fines fibres libériennes allongées atteint 4 cm, et dans la ramie - même 35 cm.Il s'agit généralement de cellules mortes, sans protoplaste. Ils remplissent une fonction mécanique, créant la force de la tige.
La plupart des cellules spécialisées sont incapables de se reproduire. Cependant, la plante pousse tout au long de sa vie et tout au long de sa vie, de nouvelles cellules s'y forment. Ils se développent à partir des cellules de tissus éducatifs - méristématiques -. La reproduction par division est une spécialisation des cellules méristématiques, leur fonction dans le corps. Les cellules qui en proviennent se développent et se transforment en l'une ou l'autre des cellules spécialisées - cellules de tubes tamis, cellules de liège, fibres, etc. Le tissu éducatif est situé dans différentes parties de la plante (par exemple, aux points de croissance - au sommet des pousses, des racines). Il y a généralement plusieurs couches de tissu éducatif dans l'épaisseur de la tige. Sous la couche de liège, il y a des cellules phellogènes qui, en se divisant, reconstituent le nombre de cellules de liège et de croûte. Une couche de cellules de cambium est située à l'intérieur du bast. Celles des cellules qu'elles génèrent, qui se forment à l'extérieur du cambium, se transforment en cellules libériennes (Fig. 25). Une fois à l'intérieur, c'est-à-dire plus près de l'axe du tronc, donnent naissance à des cellules de bois.
Le bois se compose d'éléments de tissus conducteurs, de support et de base. Les premiers comprennent les fibres de bois - de longues cellules mortes avec des parois lignifiées; au second - vaisseaux, qui sont le résultat de la fusion de nombreuses cellules; à la troisième - cellules du parenchyme ligneux. Les cellules qui se différencient des éléments cambiaux en cellules vasculaires (vaisseaux) se développent en longueur et en largeur. Leurs parois latérales sont épaissies et lignifiées. Dans les murs, cependant, des pores restent, fermés seulement par une fine coque. Les cloisons aux jonctions des cellules adjacentes disparaissent, le protoplaste meurt. En conséquence, de longs vaisseaux se forment, composés de quelques murs. Le système conducteur s'étend à travers la racine et la tige dans la feuille. À travers de tels vaisseaux, un flux ascendant d'eau et de sels dissous est effectué des racines à tous les organes de la plante.
Au centre de la tige se trouvent des cellules centrales - des cellules parenchymateuses rondes ou polyédriques. Ce sont les éléments du tissu principal. Parfois, ils sont creux et contiennent de l'air; parfois ils sont remplis de nutriments de réserve, de cristaux divers, de tanins. Leurs murs peuvent être lignifiés. Les rayons radiaux pénètrent dans le bois et le liber. Leurs cellules sont des dérivés du cambium et ont une fonction de stockage. Au niveau de la bast, ces rayons se dilatent sensiblement.
Dans la pulpe des feuilles, entre l'épiderme supérieur et inférieur, se trouve le tissu principal - des cellules à membranes fines et un grand nombre de plastes verts - les chloroplastes. La photosynthèse a lieu dans ces cellules. Les couches supérieures sont principalement constituées de cellules oblongues, étroitement adjacentes les unes aux autres - ce parenchyme colonnaire. Les couches inférieures se relâchent entre les cellules, il y a des espaces intercellulaires - des espaces remplis d'air; c'est un parenchyme spongieux. Le parenchyme est imprégné de veines ramifiées, constituées de faisceaux de tissus mécaniques (diverses fibres) et conducteurs (tubes de tamis et aquifères).
![]()
Cette présentation brève et schématique du plan de la structure de la tige, feuille montre à quel point la taille, la forme, la structure, les fonctions des cellules d'une même plante sont diverses. Si vous prenez différentes plantes, vous pouvez rencontrer une plus grande variété de cellules. Cependant, en règle générale, les cellules des tissus du même nom, même dans les plantes très éloignées les unes des autres, sont similaires, car ces cellules remplissent des fonctions similaires. Les cellules épidermiques d'une feuille de bouleau et, par exemple, le pissenlit sont plus similaires les unes aux autres que la cellule épidermique d'un pissenlit et sa cellule de tamis. Les traits distinctifs de la cellule sont principalement associés à sa fonction spéciale. Les cellules spécialisées dans la fonction mécanique ont des membranes épaissies et souvent lignifiées. La spécialisation en photosynthèse conduit à l'apparition de chloroplastes dans les cellules. La spécialisation dans la fonction conductrice est associée à l'allongement cellulaire, à la perte du protoplaste, à des modifications des membranes cellulaires aux jonctions, grâce auxquelles les cavités des cellules adjacentes communiquent. Les cellules spécialisées dans la fonction protectrice sont caractérisées par divers changements dans les parois extérieures, la présence de poils et la capacité à produire des substances protectrices.
La spécialisation dans la fonction de stockage des nutriments peut conduire à une augmentation de la taille des cellules, à l'apparition de très grandes vacuoles. Dans les cellules méristématiques, à division intensive, ces structures intracellulaires sont spécialement développées pour assurer la synthèse de substances - les parties constitutives de la cellule.
Selon la nature de la spécialisation, certaines caractéristiques de la structure et du travail de la cellule sont très fortement développées, tandis que d'autres, au contraire, sont peu développées ou totalement absentes. Comme nous l'avons vu, certaines cellules, devenues spécialisées, meurent et c'est à l'état mort qu'elles commencent à remplir leur fonction particulière dans un organisme multicellulaire (cellules de liège, fibres de bois, vaisseaux).
Les cellules des algues filamenteuses multicellulaires et des plantes unicellulaires sont très diverses et particulières (fig. 26 et 27). De plus, la cellule de l'un quelconque de ces derniers est très différente des cellules des plantes multicellulaires. Elle seule doit remplir plusieurs fonctions qui, chez les plantes multicellulaires, sont réparties entre des cellules de tissus différents.
Dans le même temps, même des cellules très différentes ont de profondes similitudes de structure et de fonction. Pour les organismes multicellulaires, cela est dû, d'une part, au fait que toutes les cellules du corps (si l'on exclut la reproduction végétative, dans laquelle tout un complexe multicellulaire est immédiatement séparé de la plante mère en tant que nouvel individu ou son rudiment) sont des descendants de la même cellule progénitrice. Par conséquent, quelle que soit la spécialisation des cellules, elles ont une origine commune et sont donc apparentées.
Deuxièmement, les caractéristiques communes de la structure des cellules de plantes de différentes espèces sont associées au fait que toutes les plantes sont liées à un degré ou à un autre. Toutes les plantes ont évolué par évolution d'ancêtres unicellulaires communs. Les caractéristiques communes sont héritées par les cellules des plantes modernes d'anciennes cellules progénitrices. Ceci est également associé à la présence de caractéristiques communes de la structure et du travail des cellules végétales et animales.
Troisièmement, la similitude est due au fait que toutes les cellules vivantes, quelle que soit la fonction spéciale qu'elles exercent dans le corps, doivent avant tout assurer leur propre vie. Les cellules absorbent les nutriments, les traitent, gagnent de l'énergie et construisent leur propre corps, respirent, se débarrassent des substances inutiles, combattent divers dommages, réagissent aux changements des conditions extérieures, reconstruisent leur activité vitale et se développent. Tous ces processus dans différentes cellules sont effectués de manière similaire et à l'aide de structures du même type, qui sont communes en termes de structure non seulement pour différentes cellules végétales, mais également pour les cellules végétales et animales. Il faut dire que la performance de toute fonction spéciale d'une cellule est basée sur ses propriétés générales. L'une ou l'autre caractéristique, une capacité inhérente à toutes les cellules, se développe particulièrement fortement dans une cellule spécialisée et garantit que la cellule remplit sa fonction principale et spéciale. Ces caractéristiques communes, sans lesquelles il est impossible d'exécuter cette fonction spéciale, sont conservées dans la cellule et le reste peut être perdu. Les cellules spécialisées mortes en sont un cas extrême. La fonction spéciale de ces cellules est liée à leur membrane; le protoplaste n'est nécessaire que tant qu'il crée une coquille; après cela, il meurt, et la cellule entière se compose uniquement d'une coquille inanimée, qui fonctionne pour les besoins de la plante.
Compte tenu des caractéristiques générales de la structure et de l'activité vitale des cellules végétales, il convient de parler d'une certaine cellule typique qui a absorbé les caractéristiques communes de diverses cellules spécialisées. Une telle cellule, bien qu'elle n'existe pas sous une "forme pure", peut même être représentée (tableau 7, figure 28). Parmi les cellules réelles, elles sont plus similaires à de telles cellules "typiques" du parenchyme foliaire (tableau 8, figure 29).
Passons à la description de la structure et du travail d'une cellule végétale.
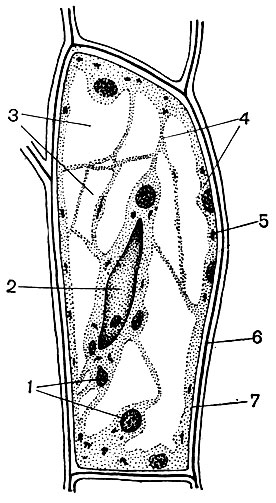
Tableau 5. Angiospermes: en haut à gauche - fleur de magnolia; en haut à droite - fleurs de pommier; ci-dessous est un lotus.
Tableau 6. Angiospermes: ci-dessus (de gauche à droite) - cloches, pélargonium, safran (crocus); en bas à gauche - asters; en bas à droite - bardane.
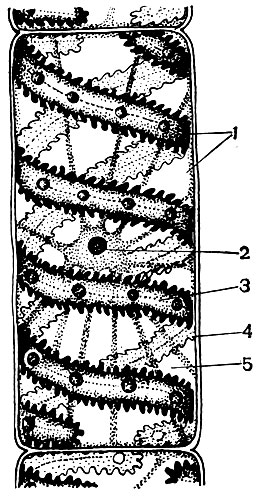
À l'extérieur, la cellule végétale est recouverte d'une membrane qui n'a pas la même épaisseur et la même structure dans différentes cellules. Les substances qui le forment sont produites dans le cytoplasme et se déposent à l'extérieur de celui-ci, créant progressivement une coquille. Ces substances sont principalement des polysaccharides à gros poids moléculaire - pectine, hémicellulose et cellulose en petites quantités. Ils forment la soi-disant coquille primaire. Il est assez élastique, car la cellule se développe, s'étire et se développe également, et n'interfère donc pas avec la croissance cellulaire. Cependant, il crée une certaine résistance de la cellule et est capable de la protéger des dommages mécaniques. Il existe des cellules dépourvues d'une telle membrane primaire - ce sont des cellules qui servent à la reproduction asexuée et sexuée (zoospores et gamètes d'algues et de champignons inférieurs, gamètes mâles de plantes supérieures). De nombreuses cellules ont non seulement une membrane primaire, mais également une membrane secondaire. Il est formé sous le primaire et est construit principalement à partir de cellulose. La cellulose est un polysaccharide dont les molécules forment les filaments les plus fins - les microfibrilles. Dans l'enveloppe, les filaments de cellulose sont immergés dans une substance amorphe constituée de composés de pectine. Dans certaines cellules, ces microfibrilles de cellulose sont situées sur toute la longueur de la cellule, en anneaux; pour cette raison, ces cellules peuvent s'étirer en longueur, mais pas en largeur (par exemple, les cellules des vaisseaux de la tige). D'autres ont des fils longitudinalement; les cellules avec une telle membrane sont élastiques lorsqu'elles sont étirées à travers, mais très rigides en étirement longitudinal. Dans d'autres, ils sont situés obliquement, formant une spirale (poils épidermiques de graines de coton, fibres libériennes). Tout cela ressemble à des structures en béton armé, et les fils de cellulose jouent ici le rôle de tiges de fer, et les substances pectines jouent le rôle de ciment. Les cellules avec une membrane secondaire sont très durables. Ils forment les tissus de soutien mécaniques de la plante. Parfois, la coquille secondaire joue également le rôle d'un entrepôt de produits nutritifs: les substances qui la forment peuvent être converties en d'autres, plus simples, qui sont consommées comme nourriture.
Dans la coquille, il y a des endroits non épaissis - les pores (dans la coquille primaire, ils sont appelés champs de terriers), à travers lesquels la communication entre les cellules voisines est effectuée. De minces cordes de cytoplasme traversent les champs de pores et les pores. Ce sont des plasmodesmes qui lient le cytoplasme des cellules voisines (tableau 9). Ils échangent des substances entre les cellules voisines. Les plasmodesmes, ainsi que des éléments de tissu conducteur, relient les cellules et les tissus du corps en un seul tout.
L'échange de substances et la propagation de l'excitation permettent aux cellules d'influencer le développement et le travail de l'autre, et chaque tissu affecte les fonctions vitales de tous les autres tissus. Cela crée la coordination du travail de toutes les parties d'un seul organisme, la plante entière.
Dans de nombreuses cellules, la membrane cellulaire avec l'âge est imprégnée de substances qui la renforcent davantage. L'accumulation de lignine dans celui-ci conduit à la lignification de la coquille. Les parois des cellules du bois et, souvent, les fibres libériennes, les coquilles cellulaires de l'écorce de la graine, parfois le péricarpe (Fig.30) et même les pétales se lignifient. Les cellules aux parois lignifiées sont non seulement plus résistantes, mais aussi moins perméables aux microbes et à l'eau. Les membranes de certaines autres cellules - tissus tégumentaires, ainsi que sur la surface des plaies - deviennent bouchonneuses, imbibées de subérine. Le contenu des cellules meurt alors, car la subérine est imperméable à l'eau et aux gaz, mais le tissu de ces cellules mortes protège les parties vivantes internes de la plante des influences externes nocives. Comme déjà mentionné, les membranes externes des cellules épidermiques sont généralement imprégnées ou recouvertes de cutine et de cire, ce qui empêche les cellules d'une évaporation excessive de l'eau, de l'infection. L'enveloppe peut également être imprégnée de minéraux, principalement des sels de calcium.
Cependant, il y a des moments dans la vie des cellules où leur membrane devrait devenir moins durable qu'auparavant. Cela se produit, par exemple, lors de la germination des graines.
La membrane cellulaire polysaccharidique est une caractéristique de la structure d'une cellule végétale, qui la distingue d'une cellule animale.
La membrane, ou paroi cellulaire, est une formation protectrice. Il y a un cytoplasme sous la membrane. Sa couche la plus externe, adjacente à la membrane, est la membrane cellulaire de surface - le plasmalemme. C'est une combinaison de couches de molécules lipidiques et protéiques. Ces membranes sont appelées membranes lipoprotéiques («lipos» - graisse, «protéine» - protéine). Une membrane de cette conception délimite le cytoplasme des vacuoles; cette membrane est appelée un tonoplaste. De nombreux organites cellulaires sont construits à partir de membranes lipoprotéiques. Cependant, dans chaque cas, la membrane est constituée de substances de type graisse (lipides) et de protéines inhérentes à cette membrane particulière. La diversité qualitative des lipides et en particulier des protéines est colossale, d'où la grande variété de membranes dont les propriétés diffèrent, tant au sein d'une même cellule que dans des cellules différentes.
Le plasmalemme régule l'entrée des substances dans la cellule et leur sortie, assure la pénétration sélective des substances dans et hors de la cellule. Le taux de pénétration à travers la membrane de différentes substances est différent. L'eau et les substances gazeuses y pénètrent bien. Les substances liposolubles pénètrent également facilement, probablement en raison du fait qu'elles comportent une couche lipidique. On suppose que la couche lipidique de la membrane est imprégnée de pores. Cela permet aux substances insolubles dans les graisses de passer à travers la membrane. Les pores portent une charge électrique, de sorte que la pénétration des ions à travers eux n'est pas totalement gratuite. Dans certaines conditions, la charge des pores change, ce qui régule la perméabilité des membranes aux ions. Cependant, la membrane n'est pas également perméable à différents ions avec la même charge, et à différentes molécules non chargées de tailles proches. C'est la manifestation de la propriété la plus importante de la membrane - la sélectivité de sa perméabilité: pour certaines molécules et ions, elle est mieux perméable, pour d'autres, elle est pire.
Une substance pénètre dans la cellule et s'y diffuse si sa concentration à l'extérieur est supérieure à la concentration à l'intérieur. Sinon, il devrait se diffuser hors de la cellule. Mais, ayant pénétré dans la cellule, les molécules de cette substance peuvent être immédiatement impliquées dans le métabolisme et se transformer en d'autres substances. Ensuite, la concentration de cette substance à l'intérieur de la cellule baisse à nouveau et une nouvelle partie de ses molécules se diffuse dans la cellule. En conséquence, les cellules qui consomment activement telle ou telle substance, cette dernière pénètre de manière intensive depuis les cellules environnantes.
Un autre mécanisme pour une meilleure absorption d'une substance par une cellule est sa liaison par des protéines ou d'autres substances. En se liant, la substance quitte la solution intracellulaire et n'interfère pas avec la diffusion ultérieure de la même substance. Il continue à entrer dans la cellule et s'y accumule sous une forme liée. Ainsi, dans les cellules de certaines algues, l'iode s'accumule à des concentrations un million de fois supérieures à sa concentration dans l'eau de mer.
Dans tous ces cas, on parle du mouvement des substances par diffusion. Cependant, les membranes cellulaires ont également des mécanismes pour déplacer les substances à travers elles-mêmes d'une concentration inférieure à une concentration supérieure. Parmi les protéines de la membrane, il existe des substances protéiques dont le travail consiste à déplacer diverses substances d'un côté de la membrane à l'autre. Ils sont appelés enzymes de transport. Probablement, une telle enzyme de transport forme un intermédiaire avec la substance transportée; celle-ci passe à travers la membrane, puis se décompose en ses parties d'origine, après quoi l'enzyme revient et la substance transférée reste de l'autre côté de la membrane. Les détails de ce processus sont inconnus, mais le fait même d'un tel transport actif de substances ne fait aucun doute. Contrairement au transport enzymatique diffus, passif, nécessite une dépense énergétique. Probablement, c'est précisément grâce à ce transport actif d'ions que les cellules de l'épiderme racinaire sont capables d'absorber les substances inorganiques nécessaires à la plante du sol puis de les transférer à travers la plante d'une cellule à l'autre.
Avec les changements chimiques ou physiques de l'environnement externe et interne, les membranes cellulaires modifient leur perméabilité, ainsi que le degré et la nature même de sa sélectivité. Les mécanismes de régulation du mouvement des substances dans et hors de la cellule sont basés sur cela. Une modification de la perméabilité des membranes aux nutriments se reflète dans l'intensité des processus métaboliques dans la cellule, sur la nature des synthèses qui s'y déroulent et tout au long de sa vie. Le changement de la perméabilité des membranes aux ions sodium et potassium est le mécanisme de propagation de l'excitation cellulaire - l'émergence et le mouvement du biocourant. Un grand nombre de cations, principalement des ions potassium, sont attachés à la surface de la membrane. Par conséquent, il porte une charge positive de l'extérieur. Sous l'influence de stimuli, les ions potassium sont séparés de la zone affectée, la charge de la zone diminue et elle devient électronégative par rapport aux zones voisines de la membrane. Cette électronégativité est, à son tour, un irritant pour les zones voisines, de la même manière en supprimant leur charge, en les dépolarisant. La vague d'électronégativité se propage à travers la membrane - c'est le courant biologique. Ensuite, les ions potassium reposent à nouveau sur la membrane, lui donnant la charge initiale - une vague de dépolarisation est suivie d'une vague de restauration. Les surfaces des plasmodesmes traversant les parois cellulaires et reliant le cytoplasme des cellules voisines sont également formées par de telles membranes. Le biocourant se déplace également le long d'eux, se propageant de cellule en cellule. Les biocourants des cellules végétales sont encore mal connus. Cependant, il est clair qu'il s'agit d'une méthode de signalisation utilisée dans une cellule végétale pour déclencher certaines réactions chimiques et en inhiber d'autres. Le fait est que tous les processus chimiques se produisant dans la cellule et constituant l'essence de son activité vitale sont réalisés avec la participation de catalyseurs biologiques - des enzymes. Chaque réaction n'est possible que lorsque l'enzyme qui la déclenche est active. La plupart des enzymes deviennent actives sous l'action de certains cations inorganiques: K., Na. , Ca .., Mg .., Mn ... Le biocourant, modifiant la perméabilité de la membrane pour les substances chargées, crée les conditions de leur pénétration à travers les membranes et du contact avec les enzymes. Ainsi, le biocourant active certaines enzymes et régule et dirige ainsi le métabolisme dans la cellule. La régulation du métabolisme par les biocourants n'est que l'un des nombreux moyens de réguler le métabolisme intracellulaire.
Ce qui a été dit au sujet de la perméabilité de la membrane de surface de la cellule - le plasmalemme - s'applique également à d'autres membranes intracellulaires, y compris celles dont de nombreuses organites cellulaires sont construites.
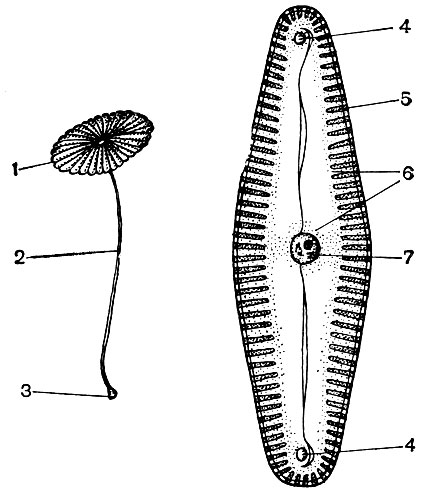
Le cytoplasme, autrefois considéré comme une solution colloïdale homogène de substances protéiques, est en fait structuré de manière complexe. Avec le développement des techniques de recherche microscopique, des détails de plus en plus subtils de la structure du cytoplasme ont été clarifiés. Dans le cytoplasme, divers organites (organites) ont été découverts - des structures dont chacune remplit certaines fonctions physiologiques et biochimiques. Les organites les plus importants du cytoplasme sont les mitochondries, le réticulum endoplasmique (réticulum endoplasmique), l'appareil de Golgi, les ribosomes, les plastes, les lysosomes. Les cellules mobiles (zoospores et gamètes d'algues, spermatozoïdes de prêles, fougères, cycadales, certaines algues unicellulaires et coloniales) ont des organites de mouvement - les flagelles.
Surtout beaucoup de faits nouveaux sur la structure fine du cytoplasme ont amené et amènent l'utilisation d'un microscope électronique, qui permet d'étudier les détails de la structure des organites eux-mêmes. Les méthodes biophysiques et biochimiques modernes permettent d'isoler certains organites du cytoplasme sous forme pure puis de les étudier composition chimique et leurs fonctions. En dehors de la cellule, dans des environnements de composition complexe, de nombreux organites sont capables de faire le travail qu'ils font lorsqu'ils sont dans la cellule.
La partie du cytoplasme dans laquelle les organites sont immergés et qui jusqu'à présent semble sans structure est appelée la substance de base du cytoplasme ou hyaloplasme. Hyalo-plasma n'est en aucun cas une charge passive, mais une partie active du cytoplasme. Un certain nombre de processus chimiques vitaux s'y déroulent, il contient de nombreuses protéines enzymatiques, à l'aide desquelles ces processus sont effectués.
La vie cellulaire consiste en un travail chimique continu, qui dans sa totalité est appelé métabolisme. Essentiellement, la cellule est une usine chimique qui produit une large gamme de produits et extrait indépendamment l'énergie nécessaire pour la produire. Ses produits sont des substances dont elle-même a besoin pour maintenir sa propre vie (pour construire son corps pendant la croissance et le développement, pour remplacer ses parties usées) et pour créer des cellules filles pendant la reproduction et pour les besoins des autres cellules du corps.
Toutes les réactions chimiques qui ont lieu dans la cellule peuvent être divisées en deux groupes. En conséquence, certaines de ces substances ou de ces substances se décomposent en molécules plus petites. À la suite d'autres substances de faible poids moléculaire, des substances avec des molécules plus grosses sont synthétisées. Une molécule de n'importe quelle substance est constituée d'atomes qui sont maintenus ensemble par des liaisons chimiques, c'est-à-dire par une certaine quantité d'énergie chimique potentielle concentrée. Lorsque la molécule est rompue, les liaisons sont rompues et leur énergie chimique est libérée. Pour la synthèse, c'est-à-dire pour la formation d'une plus grande molécule à partir de petites, il est nécessaire de créer de nouvelles liaisons chimiques. Il est nécessaire d'y mettre une partie d'énergie. Au contraire, les réactions de désintégration se déroulent finalement avec la libération d'énergie, car pendant elles les liaisons entre les atomes sont rompues. La signification biologique des réactions de désintégration en cours dans la cellule est qu'elles libèrent de l'énergie chimique, qui est ensuite utilisée par la cellule pour des réactions de synthèse et pour la production d'autres types de travail (électrique, mécanique, travail sur le transport de substances). Comme la cellule a synthétisé diverses substances tout au long de sa vie, elle doit continuellement décomposer d'autres substances. L'énergie libérée n'est pas immédiatement utilisée pour les synthèses. Premièrement, il est stocké par la formation de substances spéciales - accumulateurs d'énergie chimique - acide adénosine triphosphorique (ATP) et composés apparentés. Au bon moment et au bon endroit dans la cellule, l'ATP est scindé et abandonne l'énergie pour la synthèse de la substance nécessaire à la cellule.
Toutes les réactions chimiques dans la cellule - à la fois la synthèse et la désintégration - sont effectuées à l'aide d'enzymes. Les enzymes sont des substances protéiques qui accélèrent le cours des réactions. Cette accélération est si grande que sans enzymes, de telles réactions ne seraient pas du tout possibles dans la cellule. Il y a des cas où, grâce à l'enzyme, la réaction est accélérée 1011 fois. Cela signifie qu'une réaction qui se termine par la participation de l'enzyme dans un délai de 0,01 s, sans elle, se déroulerait pendant 31 ans. Il est clair que de telles réactions sans l'enzyme seraient tout simplement irréalistes.
De plus, grâce aux enzymes, le déroulement des réactions chimiques dans la cellule est contrôlé et régulé. L'activité des enzymes dans une cellule change en fonction de son besoin d'une certaine substance ou énergie, c'est-à-dire à la suite du travail de ces mêmes enzymes. Lorsqu'une substance est nécessaire, ces enzymes, grâce auxquelles elle se forme, sont activées ou synthétisées à nouveau. La formation de substances, la libération et le stockage d'énergie sont le résultat de réactions successives, le résultat du travail de toute une chaîne d'enzymes. Toutes les structures des parties vivantes de la cellule sont construites précisément à partir d'enzymes et de substances qui maintiennent ces enzymes ensemble. Dans ce cas, les enzymes participant à des réactions adjacentes se déroulant séquentiellement sont placées côte à côte. Ils transfèrent les molécules des substances qu'ils convertissent les uns aux autres, comme sur un convoyeur, et chacun d'eux effectue sa propre opération de travail sur la molécule.
L'hyaloplasme contient des enzymes qui décomposent les molécules de glucose en molécules d'acide pyruvique plus simples. L'énergie libérée dans ce cas est stockée par la formation de molécules d'ATP. Le même processus a lieu dans le noyau cellulaire. Cependant, la majeure partie de l'énergie est obtenue dans des organites spéciaux du cytoplasme - les mitochondries, car il y a une dégradation plus profonde des substances.
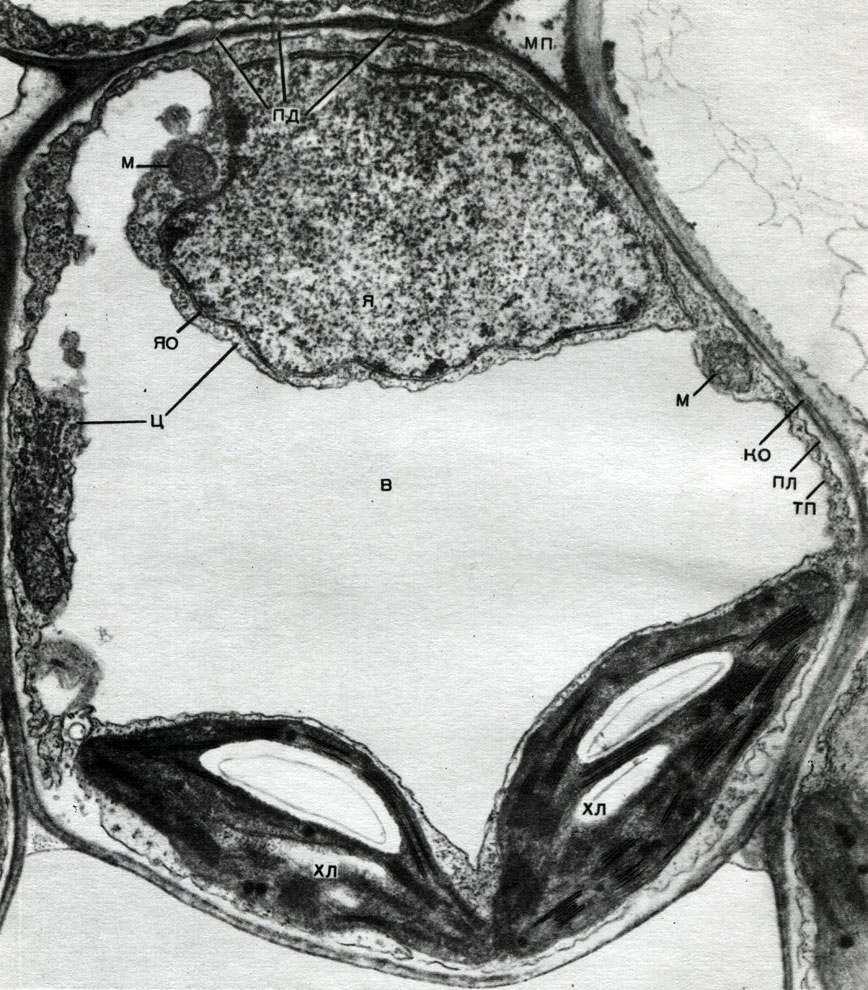
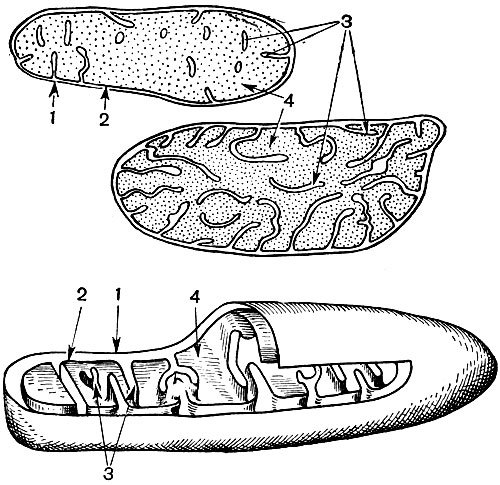
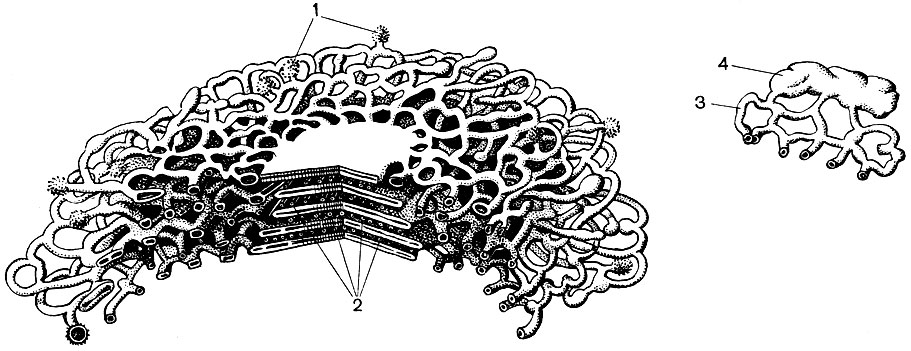
Les mitochondries sont de petits corps de forme ronde ou oblongue, de 0,5 à 1,5 microns, c'est-à-dire de la taille d'une bactérie. Leur nombre dans une cellule est généralement grand, environ 100 à 3000. Il existe cependant des cellules avec un petit nombre de mitochondries. Ainsi, le sperme de l'algue fucus ne contient que 4 mitochondries, et l'algue unicellulaire micromonas en contient une. Les mitochondries sont visibles au microscope optique, mais leur structure fine ne peut être étudiée qu'au microscope électronique (Tableau 10, schéma de la structure - Fig. 31). Les mitochondries sont des formations construites à partir de membranes lipoprotéiques immergées dans une substance basique - une matrice. La membrane mitochondriale est formée de deux membranes entre lesquelles il y a un espace.
La membrane interne de la coquille donne de nombreuses invaginations à l'intérieur, ce sont des crêtes. Il y a une matrice entre eux. La membrane interne de la membrane mitochondriale et les crêtes qu'elle forme sont construites à partir de l'ordre d'enzymes localisées. En raison des plis - crêtes, la surface de travail des membranes à l'intérieur des mitochondries est très grande. Un certain nombre d'enzymes se trouvent dans la matrice mitochondriale, c'est-à-dire entre les crêtes.
L'association de ces enzymes assure la respiration intracellulaire et le stockage de l'énergie libérée lors de la respiration sous forme d'ATP. Le travail des mitochondries est étroitement lié aux processus se produisant dans le hyaloplasme, où se déroulent les premières étapes de la dégradation du glucose et d'autres substances en acide pyruvique. Dans les mitochondries, sa division supplémentaire a lieu. L'acide pyruvique pénètre dans les mitochondries et ici, étape par étape, il est oxydé en dioxyde de carbone et en eau, et l'oxygène est simultanément consommé. Il s'agit de la respiration intracellulaire, dans laquelle la cellule, qui se divise et s'oxyde, produit beaucoup d'énergie, qu'elle peut ensuite utiliser pour une variété de ses besoins.
La première étape de la dégradation de la molécule de glucose, au cours de laquelle elle est divisée en deux et qui a lieu dans l'hyaloplasme, ne donne à la cellule que deux molécules d'ATP.
À la suite de la deuxième étape, conduisant à la «combustion» complète du glucose, 36 molécules d'ATP supplémentaires sont formées. Par conséquent, les mitochondries par leur fonction sont les centrales électriques de la cellule, des machines pour obtenir l'essentiel de l'énergie. La dégradation même des produits de dégradation du glucose se produit dans la matrice mitochondriale, tandis que l'ATP se forme en raison de réactions se déroulant sur ses membranes internes, qui comprennent des enzymes respiratoires et des enzymes qui assurent la formation d'ATP. Le nombre de crêtes dans les mitochondries peut varier. Plus il y en a, plus l'activité biochimique des mitochondries est élevée.
Nous parlons ici du glucose en tant que substance, décomposant dont la cellule produit de l'énergie. Le glucose est une substance centrale, mais pas la seule. Sa molécule a un squelette de six atomes de carbone reliés les uns aux autres. À la suite d'une longue et complexe chaîne de réactions, sa molécule est broyée, oxydée et, à la fin, divisée en six molécules d'une substance inorganique - le dioxyde de carbone (CO 2), dont chaque molécule ne contient qu'un seul atome de carbone, et elle est extrêmement oxydée. En additionnant toutes les réactions successives d'oxydation du glucose et en excluant tous les produits intermédiaires, vous pouvez obtenir la réaction totale de ce processus:
L'amidon est facilement converti en glucose, après quoi il subit la dégradation décrite ci-dessus. Les protéines et les graisses donnent divers acides organiques, qui sont convertis en produits intermédiaires de la dégradation du glucose puis oxydés de la même manière que ce dernier, et à l'aide des mêmes enzymes.
L'oxydation biologique complète de la matière organique est similaire à sa combustion. Dans les deux cas, le résultat est le dioxyde de carbone, l'eau et l'énergie libérés. Cependant, lors de la combustion, cette énergie est libérée sous forme de chaleur, et complètement à la fois; lors de l'oxydation biologique, l'énergie des liaisons chimiques est libérée par portions, et sa partie principale est liée, passant dans l'énergie de la liaison chimique phosphate de l'ATP. En conséquence, la cellule reçoit un concentré d'énergie sous une telle forme, qui ensuite au bon moment et au moment approprié peut être utilisé pour créer de nouvelles liaisons chimiques, pour synthétiser de nouvelles substances, ainsi que pour produire d'autres types de travail - électrique, mécanique, ainsi que des travaux sur le transport de substances d'environnement en cellule, de cellule en environnement et de cellule en cellule.
Plus l'activité vitale de la cellule est active, plus son besoin d'énergie est grand et plus elle contient de mitochondries. Ils peuvent être inégalement répartis au sein d'une même cellule: il y en a plus dans la partie de la cellule qui travaille actuellement plus activement.
Les mitochondries sont capables de synthétiser certaines des substances dont elles sont elles-mêmes composées. Grâce à cela, les mitochondries peuvent se multiplier.
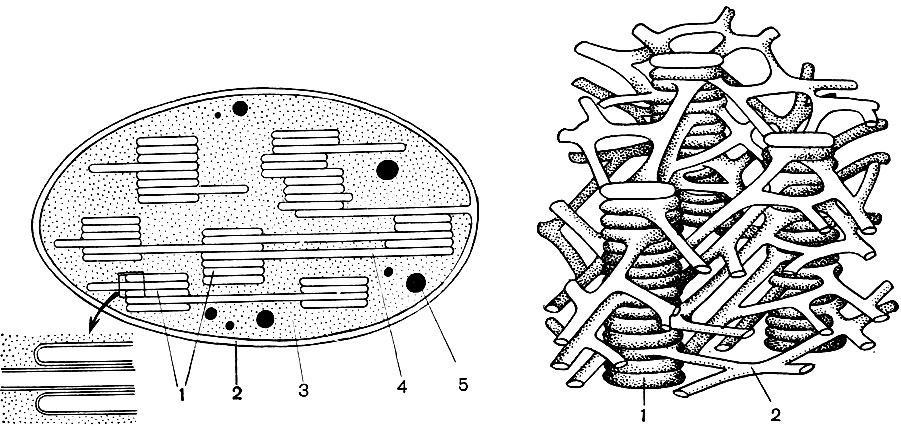
Le réticulum endoplasmique est un organoïde du cytoplasme, dans lequel un grand nombre de substances sont synthétisées (tableau 10). Le réticulum endoplasmique est un système de canaux qui imprègnent le cytoplasme et qui se rétrécissent dans certaines zones, se dilatent dans d'autres, formant soit des citernes, soit des sacs plats, soit des tubes de ramification. Les parois de toutes ces formations sont construites de membranes qui contiennent des enzymes.
Comme dans d'autres formations membranaires de la cellule, les enzymes du réticulum sont disposées de manière ordonnée. Dans ce cas, les enzymes voisines effectuent des réactions séquentielles (opérations de travail) et leur groupe réalise toute la chaîne de réactions conduisant à la création d'une substance particulière.
Distinguer le réticulum endoplasmique agranulaire (lisse) et granuleux. Sur la surface externe des canaux du réticulum granulaire, il existe de nombreux petits organites - ribosomes, dont la fonction est la synthèse de molécules protéiques. Le réticulum endoplasmique agranulaire, qui prévaut quantitativement sur le réticulum granulaire dans la cellule végétale, ne porte pas de ribosomes.
Le réticulum endoplasmique, en plus d'être un transporteur pour de nombreux types de conversion enzymatique de substances, principalement pour leur synthèse, est également un système d'autoroutes le long duquel les substances se déplacent à travers la cellule. Le réticulum part de la membrane externe de la membrane du noyau et, en se ramifiant, s'approche de divers organites du cytoplasme, ainsi que du plasmalemme. Ainsi, il relie toutes les parties de la cellule les unes aux autres. De plus, les canaux du réticulum endoplasmique traversent les plasmodesmes, reliant le réticulum des cellules voisines.
En outre, les membranes du réticulum endoplasmique démembrent le cytoplasme en de nombreux compartiments, en raison desquels la cellule ne peut pas être représentée comme une masse homogène dans laquelle une variété de substances sont mélangées. Les conditions dans un compartiment peuvent être complètement différentes des autres; les processus qui s'y déroulent ne peuvent avoir lieu qu'en lui, tandis que les uns dans les autres se déroulent des processus.
Enfin, les membranes du réticulum endoplasmique sont les surfaces le long desquelles se propagent les biocourants, signaux qui modifient la perméabilité sélective des membranes et donc l'activité des enzymes. Grâce à cela, certaines réactions chimiques sont déclenchées, d'autres sont inhibées - le métabolisme est soumis à une régulation et se déroule de manière coordonnée.
Bon nombre des substances synthétisées dans la cellule doivent être concentrées et libérées de la cellule soit dans l'environnement externe, soit dans la vacuole intracellulaire. De plus, une cellule concentre également les substances qui y pénètrent à partir d'autres cellules, par exemple, si elle les stocke en réserve. Ce travail est effectué par des dictyosomes. Habituellement, dans une cellule végétale, il y a plusieurs dictyosomes, et leur ensemble complet est appelé appareil de Golge (ou complexe) et la cellule donnée. Chaque dictyosome est un système de membranes empilées (Tableau 10 - coupe, Fig. 32 - diagramme volumétrique). Les cavités entre les membranes qui forment cet organoïde ressemblent à des fentes étroites, puis des sacs plats - des citernes, puis des bulles. Leur forme change au cours du fonctionnement de l'organoïde et dépend apparemment du degré de remplissage des espaces intermembranaires avec les substances sécrétées et accumulées. Les vésicules formées et envahies sont séparées de l'organoïde. Apparemment, de nombreuses vacuoles cellulaires, entourées de membranes - tonoplaste, sont le produit de l'activité de l'appareil de Golgi; ce sont des bulles qui s'en détachent puis grossissent.
L'appareil de Golgi est spécialement développé dans les cellules excrétrices (sécrétoires), dans lesquelles diverses substances sont déposées ou à partir desquelles diverses substances sont excrétées. Il synthétise et libère des substances qui forment la paroi cellulaire.
Les lysosomes sont des corps arrondis plutôt petits (environ 0,5 microns de diamètre) - un autre organoïde du cytoplasme. Ils sont recouverts d'une membrane - une membrane lipoprotéique. Le contenu des lysosomes sont des enzymes qui digèrent les protéines, les glucides, les acides nucléiques et les lipides. La membrane du lysosome empêche la libération d'enzymes de l'organoïde dans le hyaloplasme, sinon ce dernier serait digéré par ces enzymes.
On peut penser que les lysosomes sont des produits de l'activité de l'appareil de Golgi, des bulles qui s'en détachent, dans lesquelles cet organoïde a accumulé des enzymes de digestion.
Les parties de la cellule qui meurent au cours de son développement sont détruites par les enzymes lysosomales. Dans une cellule décédée, les lysosomes sont détruits, les enzymes se trouvent dans le cytoplasme et la cellule entière, à l'exception de la membrane, subit une digestion.
Les ribosomes sont de très petits organites, d'environ 250 A de diamètre. Ils sont de forme presque sphérique. Certains d'entre eux sont attachés aux surfaces externes (hyaloplasmiques) des membranes qui forment les canaux du réticulum endoplasmique granulaire; certains sont à l'état libre dans le hyaloplasme. Une cellule peut contenir jusqu'à 5 millions de ribosomes. Ce sont des "appareils de synthèse des protéines. Par conséquent, il y en a surtout beaucoup dans les cellules qui forment activement des protéines - dans les cellules en croissance, dans les cellules qui sécrètent des substances protéiques. Les ribosomes se trouvent également dans les mitochondries et les chloroplastes, où ils synthétisent certaines des protéines à organites.
Les organites appelés microtubules se trouvent dans de nombreuses cellules. Leur nom même parle de leur forme - ce sont des tubes avec un canal à l'intérieur. Leur diamètre extérieur est d'environ 250A. Parfois, ce sont des tubes doubles - deux simples, couchés côte à côte et ayant une paroi commune qui sépare leurs cavités. Les parois des microtubules sont construites à partir de molécules de protéines. On pense que les microtubules sont associés à l'activité contractile (motrice) du cytoplasme et de ses formations. À partir d'eux, comme à partir des pièces de construction, les structures contractiles du flagelle, un organoïde, à l'aide duquel se déplacent certaines algues unicellulaires et coloniales, ainsi que des cellules qui servent à la reproduction de nombreuses plantes inférieures, sont apparemment construites. Pendant la division cellulaire, les filets de broche sont formés à partir de microtubules, ce qui sera discuté ci-dessous. Pendant la période de division, les microtubules se rassemblent en groupes et forment ces filaments. En fin de division, les filaments se désintègrent à nouveau en microtubules séparés. Dans les cellules ou parties de celles-ci, dépourvues de membrane dense, les microtubules remplissent éventuellement une fonction de support, constituant le squelette interne de la cellule.
Les plastes sont des organites inhérents uniquement aux cellules végétales. Ce sont généralement de grands corps, clairement visibles au microscope optique.
Il existe 3 types de plastes: incolores - leucoplastes, verts - chloroplastes, colorés dans d'autres couleurs - chromoplastes. Chaque type de plastides a sa propre structure et a ses propres fonctions inhérentes. Cependant, des transitions de plastes d'un type à un autre sont possibles. Ainsi, le verdissement des tubercules de pomme de terre est causé par le réarrangement de leurs leucoplastes en chloroplastes. Dans la culture de racine de carotte, les leucoplastes passent dans les chromoplastes. Les trois types de plastes sont formés à partir de proplastes.
Les proplastes sont des corps incolores, semblables aux mitochondries, mais un peu plus gros qu'eux. On les trouve en grande quantité dans les cellules méristématiques. Les leucoplastes se trouvent dans les cellules des parties non colorées des plantes (fruits, graines, racines, épiderme des feuilles). Leur forme est indéfinie. Le plus souvent, il existe des leucoplastes dans lesquels de l'amidon se dépose (il est formé de sucres). Il existe des leucoplastes qui stockent des protéines. Les moins courants sont les leucoplastes remplis de graisse; ils se forment lors du vieillissement des chloroplastes. Il n'y a pas de différences significatives entre les leucoplastes et les proplastes.
Les chloroplastes sont des plastes de plantes supérieures dans lesquelles le processus de photosynthèse a lieu, c'est-à-dire l'utilisation de l'énergie des rayons lumineux pour la formation de substances organiques à partir de substances inorganiques (dioxyde de carbone et eau) avec la libération simultanée d'oxygène dans l'atmosphère Les chloroplastes ont la forme d'une lentille biconvexe, leur taille est d'environ 4 - 6 mk. Ils se trouvent dans les cellules narenchymateuses des feuilles et d'autres parties vertes des plantes supérieures. Leur nombre dans une cellule varie de 25 à 30.
La structure du chloroplaste est donnée dans les tableaux 11 et 12 (vue en coupe) et le schéma de la figure 33. A l'extérieur, le chloroplaste est recouvert d'une membrane constituée de deux membranes lipoprotéiques. En dessous, dans la substance principale (stroma), de nombreuses formations - lamelles - sont disposées de manière ordonnée. Ils forment des sachets plats qui se superposent dans des selles régulières. Ces stoïciens, qui ressemblent à des pièces de monnaie empilées dans une colonne, sont appelés grains. Des lamelles plus longues les traversent, de sorte que tous les grains de chloroplaste sont connectés en un seul système. La composition des membranes formant les granules comprend un pigment vert - la chlorophylle. C'est ici que se déroulent les réactions lumineuses de la photosynthèse - l'absorption des rayons lumineux par la chlorophylle et la conversion de l'énergie lumineuse en énergie d'électrons excités. Les électrons excités par la lumière, c'est-à-dire ceux qui ont un excès d'énergie, renoncent à l'énergie pour décomposer l'eau et synthétiser l'ATP. Lorsque l'eau se décompose, de l'oxygène et de l'hydrogène se forment. L'oxygène est libéré dans l'atmosphère et l'hydrogène est lié par la protéine ferrédoxine. La ferrédoxine est ensuite réoxydée, libérant cet hydrogène en un agent réducteur, abrégé en NADP. NADP se présente sous la forme réduite - NADP-H 2. Ainsi, le résultat des réactions lumineuses de la photosynthèse est la formation d'ATP, de NADP-H 2 et d'oxygène, et de l'eau et de l'énergie lumineuse sont consommées.
L'ATP accumule beaucoup d'énergie - il est ensuite utilisé pour les synthèses, ainsi que pour d'autres besoins de la cellule. Le NADP-H 2 est un accumulateur d'hydrogène, et il le cède ensuite facilement. Par conséquent, le NADP-H 2 est un agent réducteur chimique. Un grand nombre de biosynthèses sont associées à la réduction et le NADPH 2 agit comme un fournisseur d'hydrogène dans ces réactions.
De plus, à l'aide d'enzymes stromales chloroplastiques, c'est-à-dire à l'extérieur des granules, des réactions sombres se produisent: l'hydrogène et l'énergie contenue dans l'ATP sont utilisés pour réduire le dioxyde de carbone atmosphérique (CO2) et l'inclure dans la composition des substances organiques. La première matière organique, formée à la suite de la photosynthèse, subit un grand nombre de réarrangements et donne naissance à toute la variété des substances organiques synthétisées dans la plante et constituant son corps. Un certain nombre de ces transformations ont lieu sur place, dans le stroma chloroplastique, où se trouvent des enzymes pour la formation des sucres, des graisses, ainsi que tout ce qui est nécessaire à la synthèse des protéines. Les sucres peuvent alors soit se déplacer du chloroplaste vers d'autres structures cellulaires, et de là vers d'autres cellules végétales, soit former de l'amidon, dont les grains sont souvent visibles dans les chloroplastes. Les graisses sont également déposées dans les chloroplastes ou sous forme de gouttes, ou sous forme de substances plus simples, précurseurs de graisses, sont libérées du chloroplaste.
La complication des substances est associée à la création de nouvelles liaisons chimiques et nécessite généralement de l'énergie. Sa source est tout de même la photosynthèse. Le fait est qu'une proportion importante des substances formées à la suite de la photosynthèse se décomposent à nouveau dans le hyaloplasme et les mitochondries (dans le cas d'une combustion complète, en substances qui servent de matière de départ à la photosynthèse - CO2 et H2O). À la suite de ce processus, qui est essentiellement l'inverse de la photosynthèse, l'énergie précédemment accumulée dans les liaisons chimiques des substances décomposées est libérée et - à nouveau grâce à l'ATP - est dépensée pour la formation de nouvelles liaisons chimiques des molécules synthétisées. Ainsi, une partie importante de la production de la photosynthèse n'est nécessaire que pour lier l'énergie de la lumière et, après l'avoir transformée en produit chimique, l'utiliser pour la synthèse de substances complètement différentes.
Et seule une partie de la matière organique formée lors de la photosynthèse est utilisée comme matériau de construction pour ces synthèses.
La production de photosynthèse (biomasse) est colossale. Pendant un an dans le monde, il est d'environ 1010 tonnes. Les substances organiques créées par les plantes sont la seule source de vie non seulement pour les plantes, mais aussi pour les animaux, car ces derniers transforment des substances organiques prêtes à l'emploi, se nourrissant directement de plantes ou d'autres animaux qui, à leur tour, ils se nourrissent de plantes. Ainsi, la photosynthèse est au cœur de toute vie moderne sur Terre. Toutes les transformations de substances et d'énergie chez les plantes et les animaux sont des réarrangements, des recombinaisons et des transferts de matière et d'énergie des produits primaires de la photosynthèse. La photosynthèse est importante pour tous les êtres vivants et le fait qu'un de ses produits est de l'oxygène libre, provenant d'une molécule d'eau et rejeté dans l'atmosphère. On pense que tout l'oxygène de l'atmosphère provient de la photosynthèse. Il est essentiel à la respiration des plantes et des animaux.
Les chloroplastes peuvent se déplacer dans la cellule. En basse lumière, ils sont situés sous la paroi cellulaire qui fait face à la lumière. Ce faisant, ils se tournent vers la lumière avec leur plus grande surface. Si la lumière est trop intense, ils se tournent vers elle et s'alignent le long des ombres parallèles aux rayons lumineux. A un éclairage moyen, les chloroplastes occupent une position intermédiaire entre les deux extrêmes. Dans tous les cas, un résultat est obtenu: les chloroplastes sont dans les conditions d'éclairage les plus favorables à la photosynthèse. Un tel mouvement des chloroplastes (phototaxie) est une manifestation de l'un des types d'irritabilité chez les plantes.
Les chloroplastes ont une certaine autonomie dans le système cellulaire. Ils ont leurs propres ribosomes et un ensemble de substances qui déterminent la synthèse d'un certain nombre de protéines propres au chloroplaste. Il existe également des enzymes dont le travail conduit à la formation de lipides qui composent les lamelles et de chlorophylle. Comme nous l'avons vu, le chloroplaste dispose également d'un système de production d'énergie autonome. Grâce à tout cela, les chloroplastes sont capables de construire indépendamment leurs propres structures. On pense même que les chloroplastes (comme les mitochondries) proviennent de certains organismes inférieurs qui se sont installés dans une cellule végétale et sont d'abord entrés en symbiose avec elle, puis sont devenus sa partie intégrante, un organoïde.
Chez les plantes inférieures, la photosynthèse est également réalisée par des structures membranaires spécialisées, bien que moins développées que dans le chloroplaste. Dans les bactéries photosynthétiques, les membranes contenant de la chlorophylle forment un réseau qui imprègne le corps bactérien. Dans les algues bleu-vert, les membranes photosynthétiques sont fusionnées en bulles plates. Dans les algues vertes et autres, le système de ces membranes est séparé du reste de la cellule par une membrane de recouvrement et forme un organoïde spécial - un chromatophore. Le nombre de chromatophores dans une cellule est faible; souvent une cellule ne contient qu'un seul chromatophore. Leur forme est très différente chez les algues d'espèces différentes.
Dans Spirogyra, le chromatophore a la forme d'un ruban en spirale le long des parois cellulaires; dans Clostridium, ce sont des cylindres nervurés; le zygnema a des corps étoilés.
Les chromoplastes proviennent soit des proplastes, soit des chloroplastes, soit des leucoplastes. Leur structure de membrane interne est beaucoup plus simple que celle des chloroplastes. Gran n'est pas, le stroma contient beaucoup de pigment jaune ou orange. Les chromoplastes se trouvent dans les cellules des pétales, des fruits et des racines.
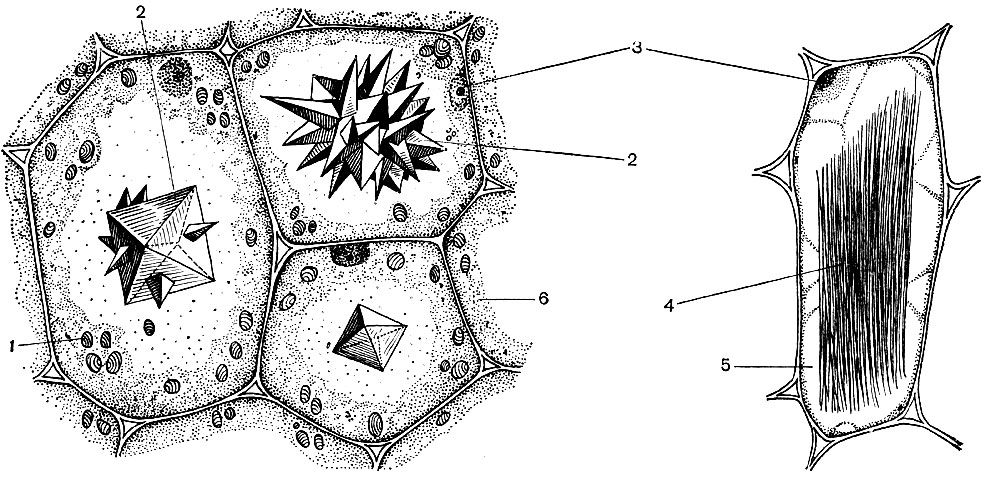
Une cellule végétale typique a une grande vacuole remplie de contenu liquide. Souvent, la vacuole occupe presque tout le volume de la cellule, de sorte que le cytoplasme n'est qu'une mince couche adjacente à la membrane cellulaire. Les jeunes cellules ont plusieurs petites vacuoles qui, à mesure que les cellules se développent, se développent et fusionnent en une seule. Le contenu de la vacuole - sève cellulaire - est une solution aqueuse de très nombreuses substances: sucres, acides aminés, autres acides organiques, pigments (colorants), vitamines, tanins, alcaloïdes, glycosides, sels inorganiques (nitrates, phosphates, chlorures), et parfois protéines.
Toutes ces substances sont les déchets de la cellule. Certains d'entre eux sont stockés dans du jus vacuolaire (cellulaire) comme substances de réserve et, au fil du temps, réintègrent le cytoplasme pour être utilisés. D'autres sont des déchets métaboliques retirés du cytoplasme. Ainsi, l'acide oxalique est éliminé dans la vacuole; les cristaux d'oxalate de calcium sont souvent déposés dans la sève vacuolaire - parfois sous la forme de monocristaux, dans d'autres cas sous la forme d'un conglomérat de cristaux de ce sel - à multiples facettes (druses) ou aciculaire (raffids) - illustré à la figure 34.
La grande majorité des plantes (à l'exception des organismes procaryotes) chaque cellule vivante a un noyau ou plusieurs noyaux. Une cellule dépourvue de noyau ne peut vivre que peu de temps. Les cellules anucléées des tubes tamis sont des cellules vivantes. Mais ils ne vivent pas longtemps. Dans tous les autres cas, les cellules non nucléaires sont mortes.

Le noyau se trouve toujours dans le cytoplasme. La forme du noyau peut être différente - ronde, ovale, fortement allongée, irrégulièrement multi-lobée. Dans certaines cellules, les contours du noyau changent au cours de son fonctionnement et des lobes de différentes tailles se forment à sa surface. Les tailles des noyaux ne sont pas les mêmes dans les cellules de différentes plantes et dans différentes cellules de la même plante. Les noyaux relativement grands se trouvent dans les jeunes cellules méristématiques, dans lesquelles ils peuvent occuper jusqu'à 3/4 du volume de la cellule entière. Les tailles relatives et parfois absolues des noyaux dans les cellules développées sont beaucoup plus petites que chez les jeunes.
À l'extérieur, le noyau est recouvert d'une coquille composée de deux membranes, entre lesquelles il y a un espace - l'espace périnucléaire. La coque est interrompue par des pores. L'extérieur des deux membranes de l'enveloppe donne des excroissances qui passent directement dans les parois du réticulum endoplasmique du cytoplasme. Les pores et la connexion directe du réticulum endoplasmique avec l'espace périnucléaire assurent un contact étroit entre le noyau et le cytoplasme.
Le contenu du noyau est une substance basique granulaire (suc nucléaire ou nucléoplasme), dans laquelle sont placées des structures plus denses - les chromosomes et le nucléole. Le nucléole est l'appareil de synthèse du matériel des ribosomes et le lieu de leur assemblage à partir de ce matériel.
Les chromosomes sont construits à partir d'un grand nombre de molécules d'acide désoxyribonucléique (ADN) associées à des molécules de protéines histones. Les molécules d'ADN sont des doubles brins longs et complexes. Chaque molécule se compose de deux brins, torsadés en spirale l'un autour de l'autre. Un fil, à son tour, est une chaîne d'un grand nombre de soi-disant nucléotides. Le nucléotide est un composé de base azotée, d'hydrate de carbone (désoxyribose) et d'acide phosphorique.
Chacun des nucléotides contient l'une des quatre bases azotées suivantes: adénine, guanine, cytosine ou thymine. En conséquence, 4 nucléotides différents sont distingués dans l'ADN: l'adénine (A), la guanine (G), la cytosine (C) et la thymine (T). Les nucléotides sont liés entre eux par leurs groupes phosphate, formant ainsi une longue chaîne. Deux chaînes, torsadées l'une autour de l'autre et formant une molécule d'ADN, sont liées entre elles par une interaction chimique (les soi-disant liaisons hydrogène) des bases azotées de leurs nucléotides. Les bases forment des paires - une base d'une chaîne, l'autre de la seconde. Le diagramme de la figure 35 donne une idée de l'ordre dans lequel les nucléotides sont liés en une chaîne, et les deux chaînes - l'une avec l'autre.
Bien que toutes les molécules d'acide désoxyribonucléique soient construites selon le plan unique décrit, leur composition qualitative spécifique est différente; ils diffèrent également par la taille des molécules. Une molécule d'ADN contient jusqu'à 50-100 mille paires de bases, mais le nombre de paires de celles-ci dans chaque molécule est différent du nombre de paires de nucléotides dans toute autre molécule. De plus, il existe 4 types différents de nucléotides, et chaque molécule d'ADN a son propre pourcentage inhérent de nucléotides de chaque type donné. En d'autres termes, le rapport quantitatif A: G: C: T pour chaque molécule d'ADN est différent. Et enfin, dans chaque molécule d'ADN, l'ordre d'alternance des nucléotides A, G, C, T n'est caractéristique que pour cette molécule. La permutation de seulement deux paires de 50 000 paires de bases, même si elles sont adjacentes, modifie considérablement les propriétés de la molécule entière; le même, en remplaçant au moins une paire, disons De - G à A - T ou G - Iz; la même chose - l'absence d'une paire parmi les mêmes 50 000. En réalité, les molécules d'ADN peuvent différer les unes des autres non pas dans un lien, pas dans une ou deux paires de nucléotides, mais dans un grand nombre d'entre elles à la fois. Le nombre de permutations possibles de paires de nucléotides dans une molécule d'ADN est infini et, en conséquence, le nombre de molécules différentes est infini, chacune ayant ses propres propriétés. La séquence de nucléotides dans une molécule d'ADN est un enregistrement crypté de la composition d'une protéine particulière qui est caractéristique d'une cellule donnée. Dans chaque molécule d'ADN, les enregistrements de la composition de plusieurs protéines sont séquentiellement ajustés, et dans toutes les molécules d'ADN de tous les chromosomes du noyau - des enregistrements de la composition de toutes les protéines d'une cellule qui peuvent y être synthétisées au cours de sa vie. L'essence de ces enregistrements codés est la suivante.
Les molécules de chaque protéine sont des chaînes d'acides aminés reliées en série. Il y a environ 20 acides aminés différents, et la nature d'une protéine est déterminée par quels acides aminés sa molécule se compose, quelle est leur quantité totale dans la molécule et dans quelle séquence ils sont connectés les uns aux autres. Par exemple, une section d'ADN responsable de la composition d'une certaine protéine (chaque section de ce type est appelée un gène) est un enregistrement d'une séquence spécifique d'acides aminés qui forment une molécule d'une protéine donnée, leur nombre total. Tous les 3 nucléotides consécutifs de la chaîne d'ADN désignent (codent) un acide aminé de la molécule protéique correspondante. Les 3 nucléotides suivants codent pour le prochain acide aminé, etc. Le nombre de combinaisons différentes possibles de 3 nucléotides parmi quatre types différents est plus que suffisant pour coder 20 acides aminés. Il est intéressant de noter que les mêmes acides aminés dans le monde vivant sont codés par les mêmes combinaisons de nucléotides. En conséquence, dans un gène, sa composition nucléotidique code la composition en acides aminés de toutes les protéines qui peuvent être synthétisées dans une cellule donnée.
Les protéines cellulaires sont des enzymes. Les enzymes déterminent le cours de toutes les réactions qui constituent l'essence de l'activité vitale des cellules. La formation et la transformation de toutes les autres substances de la cellule dépendent de la présence de certaines enzymes, que ce soit des graisses, des glucides, des alcaloïdes, des résines, etc. Ceci, à son tour, forme toutes les propriétés d'une cellule qui la distinguent des cellules d'autres plantes dans les noyaux cellulaires qui codait pour la composition d'un autre ensemble d'enzymes. C'est pourquoi les informations sur la composition de toutes les protéines qui peuvent être formées dans une cellule sont des informations sur toutes les propriétés d'une cellule et d'un organisme. Nous verrons plus loin que cette information est héréditaire, c'est-à-dire qu'elle se transmet entièrement de cellule en cellule lors de leur reproduction et de la plante mère aux plantes filles.
Ces informations sont stockées dans les chromosomes. Cependant, la réalisation de ces informations - la synthèse des protéines - ne se produit pas en eux. Les gènes chromosomiques agissent uniquement comme initiateurs de cette synthèse. Lorsqu'une cellule a besoin de former une protéine particulière, le gène dans lequel la composition de cette protéine est cryptée est activé. Cela signifie que des molécules du soi-disant acide ribonucléique informationnel (i-ARN) sont formées sur la section du brin d'ADN qui compose ce gène.
La structure et la composition de ces molécules, qui sont des chaînes uniques de nucléotides, reflètent la structure nucléotidique du gène sur lequel elles se sont formées. Ainsi, les informations sur la composition de la future protéine sont copiées.
La formation de molécules d'ARNm (beaucoup d'entre elles sont formées sur chaque gène) signifie également la multiplication des informations, en quelque sorte, l'impression de nombreuses matrices identiques qui reflètent la structure du même gène et portent ainsi des informations sur la composition en acides aminés d'une protéine donnée dans la séquence nucléotidique de leurs molécules. Ces empreintes génétiques passent du noyau au cytoplasme. Ici, les informations contenues dans les matrices - molécules d'i-ARN, sont déchiffrées, les informations sont réalisées, elles sont traduites du langage de la séquence nucléotidique dans le langage de la séquence d'acides aminés, c'est-à-dire la synthèse de molécules protéiques d'une composition donnée.
Dans le décodage et la synthèse, en plus de la molécule d'i-ARN, un grand nombre de molécules de divers acides ribonucléiques de transport (ARN-t), des ribosomes et un certain nombre d'enzymes sont impliqués. Les acides aminés se lient au t-ARN - molécule par molécule. Chacun des 20 types d'acides aminés a son propre ARN-t. Ainsi, la molécule d'ARN-t a des groupes chimiques qui peuvent reconnaître son acide aminé, en le choisissant parmi tous les acides aminés disponibles. Cela se produit à l'aide d'enzymes spéciales. Ayant reconnu son acide aminé, le t-ARN entre en connexion avec lui. Le ribosome est attaché au début de la molécule d'i-ARN, qui, se déplaçant le long de l'i-ARN, se connecte les uns aux autres dans une chaîne polypeptidique exactement les acides aminés dont l'ordre est crypté par la séquence nucléotidique de l'i-ARN donné. Cette lecture de l'information, ce décodage de celle-ci se produit grâce à un mécanisme spécial disponible dans l'ARN de transport, dont les molécules alignent exactement les acides aminés qui sont «désignés», «écrits» par la séquence nucléotidique de la molécule d'i-ARN. Le ribosome est un appareil de liaison chimique d'acides aminés dans une molécule protéique.
Ainsi, le schéma général est le suivant. Sur le gène activé - une section de l'une des chaînes d'une molécule d'ADN - des chaînes nucléotidiques, des molécules d'i-ARN, sont synthétisées, dont la composition reflète précisément la composition du gène, et porte donc un enregistrement de la composition de la protéine codée par ce gène. Dans le cytoplasme du ribosome, sur la base d'informations transférées du gène par les molécules d'ARNm, à l'aide d'ARN de transport, différents acides aminés sont combinés dans une séquence donnée, entraînant la formation d'une molécule de la protéine dont la composition est codée dans ce gène.
La formation de molécules d'i-ARN sur le gène activé sert à la fois de commande et de programme spécifique pour la synthèse d'une protéine strictement définie. La même molécule d'ARNm est utilisée comme modèle pour créer plusieurs des mêmes molécules protéiques. Cependant, il est plutôt de courte durée, par conséquent, pour une synthèse à long terme de nouvelles molécules de la même protéine, la formation de copies identiques mais nouvelles de molécules d'ARNm sur le même gène est nécessaire. Avec la transition d'un gène à un état inactif - et cela se produit lorsque le besoin de la cellule d'une protéine donnée disparaît - il est bloqué, cesse de former l'i-ARN et bientôt la synthèse de cette protéine s'arrête. Au cours de la vie d'une cellule, elle a besoin de diverses protéines. A chaque fois, les gènes qui les définissent sont activés et la composition correspondante de molécules d'i-ARN se forme.
Chaque ribosome produit de nombreuses molécules de protéines différentes au cours de sa vie. Il peut fonctionner sur la base de n'importe quel i-ARN, et le résultat, la nature de la protéine créée par celui-ci, ne dépend que de la composition de l'i-ARN en contact avec lequel le ribosome travaillait cette fois.
Ainsi, le noyau cellulaire remplit les fonctions interdépendantes suivantes. Il stocke des informations sur la composition de toutes les protéines qui peuvent être synthétisées dans une cellule donnée au cours de sa vie. (À l'exception de certaines protéines de mitochondries et de chloroplastes. Leur composition est codée dans leur propre ADN de ces organites, où se trouvent ces ADN. Ici, sur place, toutes les étapes de décodage des informations contenues dans ces ADN ont lieu, y compris la synthèse de ces protéines à l'aide de propres ribosomes.)
Le noyau stocke des informations sur toutes les propriétés de la cellule et de l'organisme. Le noyau organise la synthèse de chacune de ces protéines au bon moment. Lors de la division cellulaire, accompagnée de la division nucléaire, toutes ces informations sont transférées intégralement à chacun des noyaux nouvellement formés, à chaque nouvelle cellule. Cela est possible du fait qu'avant la division, tout le matériel génétique des chromosomes se dédouble automatiquement, deux copies identiques se forment et l'une d'entre elles se retrouve dans chacun des nouveaux noyaux. Le noyau de chaque cellule contient un ensemble complet de gènes caractéristiques d'un organisme donné. Cependant, tous les gènes ne fonctionnent pas au cours de la vie de diverses cellules spécialisées. Dans certaines cellules, une partie des gènes fonctionne, dans d'autres - une autre, dans la troisième - une troisième. C'est pourquoi les cellules de différents tissus d'un organisme diffèrent les unes des autres. Une partie importante des gènes reste dans un état passif, «stockant uniquement des informations» de la naissance à la mort d'une cellule ou d'un organisme. De plus, les gènes d'une cellule donnée ne fonctionnent pas simultanément: certains sont actifs à une période de la vie d'une cellule, à un certain stade de son développement; d'autres - de l'autre, etc.
Pourquoi certains gènes restent inactifs, comment d'autres s'allument et s'éteignent est un problème très complexe et important qui fait actuellement l'objet de recherches approfondies. Les protéines-histones semblent jouer un rôle important dans le blocage et le déblocage des gènes. Ils font partie des chromosomes, en conjonction avec l'ADN. Peut-être, l'activation, la «libération» d'un gène se produit lorsqu'une molécule d'histone est détachée de la section correspondante d'ADN, exposant ainsi ses chaînes, leur permettant de se démêler et de commencer à fonctionner chimiquement. L'attachement de l'histone conduit au blocage du gène. Cependant, la manière dont l'attachement et le détachement d'histones sont contrôlés n'est pas assez claire.
Le matériel génétique, comme dit, se trouve dans les chromosomes. Par conséquent, le travail du noyau pour stocker des informations héréditaires, pour les dupliquer et les transférer de cellule en cellule, pour organiser la synthèse de diverses protéines au cours de la vie d'une cellule est avant tout le travail des chromosomes. Bien qu'ils existent tout au long de la vie d'une cellule, ils ne se distinguent dans le noyau sous la forme de structures claires que lors de la division cellulaire. À ce moment, la condensation du matériau des chromosomes et du chromosome peut être détectée par des méthodes de microscopie en tant que formations individuelles clairement définies.
Les chromosomes ont différentes formes. Ce sont soit des bâtons droits ou courbes, soit divers crochets, corps ovales, boules (Fig. 36). Leur taille varie considérablement. Chaque cellule végétale d'une espèce donnée contient dans son noyau le même ensemble (ou ensembles) de chromosomes à partir d'un nombre strictement défini de chromosomes différents, mais strictement définis. Chez toutes les plantes supérieures et certaines plantes inférieures, deux générations alternent au cours de leur cycle de vie: avec des cellules contenant un seul (haploïde, n) ensemble de chromosomes dans leurs noyaux, et avec des cellules dont les noyaux ont un double (diploïde, 2 n) ensemble de chromosomes. Un ensemble haploïde se compose de n chromosomes différents, un de chaque type. La valeur numérique est strictement constante pour toutes les cellules haploïdes de toutes les plantes d'une espèce donnée. Ainsi, pour les fraises des bois, ce nombre est de 7, pour les pois - également 7, mais pour les haricots - 11, pour un pommier - 17, pour une poire de terre - 51, pour la canne à sucre - 60. Un ensemble diploïde est composé de deux haploïdes pliés ensemble. Il contient deux chromosomes de chaque type. Si un pois a 7 chromosomes différents dans un ensemble haploïde, alors un ensemble diploïde a 7 paires différentes de chromosomes et deux chromosomes dans chaque paire sont identiques. Il est facile de calculer que pour les plantes répertoriées, l'ensemble diploïde (2n) pour les fraises est de 14, pour les pois - 14, pour les haricots - 22, pour un pommier - 34, pour une poire de terre - 102 et pour la canne à sucre - 120 chromosomes.
Dans le cycle de vie des mousses et des fougères, les organismes alternent avec les cellules diploïdes et haploïdes. Chez les fougères, ces organismes mènent une vie indépendante et isolée. Dans les mousses, une plante diploïde vit sur une plante haploïde. Dans un organisme diploïde, des cellules haploïdes se forment - des spores. Chacun d'eux, en germination, donne un organisme haploïde, c'est-à-dire un organisme construit à partir de cellules haploïdes. Dans celui-ci, des cellules sexuelles haploïdes se forment - des gamètes.
Au moment de la fécondation, les gamètes mâles et femelles fusionnent, formant une cellule - un zygote, qui a un noyau - le résultat de la fusion des noyaux des deux gamètes. De chacun des gamètes, le noyau du zygote reçoit un ensemble haploïde de chromosomes et, par conséquent, il en a un double ensemble diploïde. Un organisme se développe à partir d'un zygote, dont chaque cellule possède un ensemble diploïde de chromosomes.

Dans un ensemble diploïde, deux chromosomes de chaque paire sont de forme et de structure interne identiques et contiennent des gènes qui contrôlent l'apparence de traits homogènes (Fig. 36). Ils sont appelés chromosomes homologues. L'un d'eux provient de l'ensemble haploïde du gamète paternel, l'autre de l'ensemble maternel. Par conséquent, dans les organismes dioïques, l'un d'eux porte des gènes qui déterminent le développement des caractéristiques sous sa juridiction selon le type paternel, le second - selon le type maternel.
Les chromosomes homologues d'une autre paire déterminent de la même manière le développement d'une autre série de caractères, la troisième paire-troisième rangée, etc. Bien que les organismes paternels et maternels appartiennent à la même espèce, leurs propriétés héréditaires ne sont pas identiques - ils ont aussi des différences individuelles. Par conséquent, certains gènes d'un chromosome homologue ne sont pas identiques aux gènes correspondants du second.
L'ensemble haploïde, qui fait partie du diploïde et issu du gamète paternel, porte l'hérédité paternelle avec ses traits individuels, et l'ensemble haploïde du gamète maternel porte celui maternel. L'interaction complexe de gènes homogènes, mais pas toujours identiques, de deux ensembles haploïdes, qui forment ensemble un diploïde, détermine quels traits apparaîtront dans la progéniture diploïde, qui, par essence, est un hybride d'un père et d'une mère.
Chez les gymnospermes et les angiospermes, l'alternance des générations se produit, en principe, comme dans les mousses et les fougères, mais leur phase haploïde est fortement réduite et n'est souvent représentée que par un groupe de cellules. Elle ne vit pas de façon indépendante, mais dans le corps d'une plante haploïde. Chez les angiospermes, la génération haploïde femelle est enfermée dans le sac embryonnaire situé dans l'ovule et la génération mâle est à l'intérieur du grain de pollen.
Même avant la division cellulaire, chaque molécule d'ADN dans chaque chromosome se fixe une copie d'elle-même - une seconde de la même molécule. En conséquence, tout le matériel héréditaire de la cellule double et chaque chromosome se compose désormais de deux parties égales - les chromatides. De plus, la "tâche" de la cellule est de diviser chaque chromosome en chromatides, de les répartir strictement également entre les futures cellules filles: une chromatide de chaque chromosome doit être envoyée à chacune d'elles. Cela se fait comme suit. Immédiatement avant la division cellulaire, les chromosomes sont fortement condensés et contractés. Ensuite, ils sont situés dans un plan - le long de l'équateur du noyau, avec une chromatide de chacun d'eux faisant face à un pôle de la cellule, l'autre - à l'opposé. L'enveloppe nucléaire disparaît et le nucléole se dissout également. Des filaments apparaissent entre les pôles de la cellule, qui forment ensemble une forme de fuseau. Les filaments sont collectés à partir de microtubules se connectant les uns aux autres. La broche se compose de deux types de filets. Certaines sont des cellules continues allant d'un pôle à l'autre. D'autres tirent, chacun connectant un pôle à l'une des chromatides. Dans le chromosome, il y a un site - le kinétochore, auquel les fils de traction sont attachés - l'un d'un pôle, l'autre de l'autre. De plus, les chromatides divergent. Les chromatides, qui constituaient auparavant chaque chromosome, sont séparés les uns des autres et, tirés vers le haut par des filaments, divergent vers les pôles opposés de la cellule. En conséquence, un ensemble complet de chromatides est collecté à chaque pôle, qui sont maintenant des chromosomes. Après cela, les fils de la broche se désintègrent, une enveloppe nucléaire se forme autour de chaque ensemble de chromosomes, les chromosomes gonflent considérablement (se déspiralisent). Un nucléole apparaît dans chaque noyau. Le noyau acquiert la structure caractéristique des noyaux des cellules non en division. Dans le plan médian de la cellule, un septum se forme, divisant la cellule en deux cellules filles.
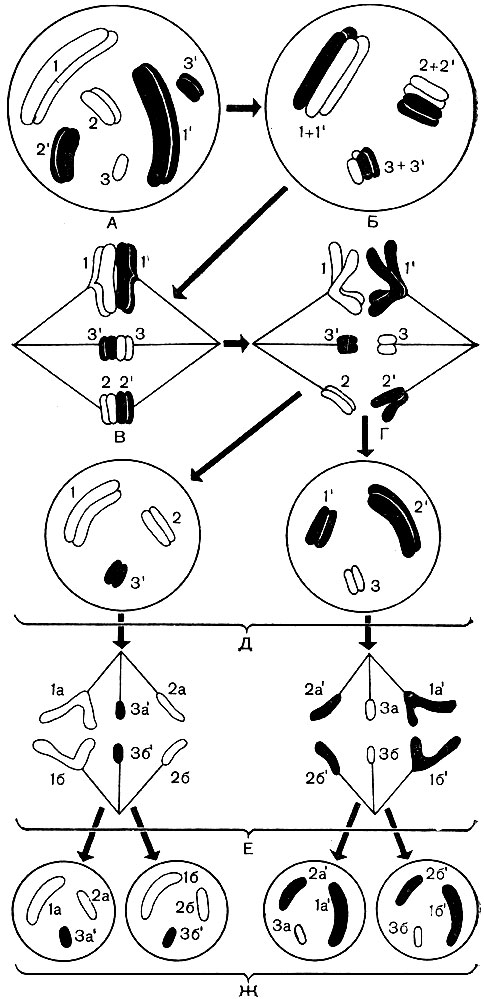
Figure: 38. Schéma du comportement des chromosomes de la cellule "conditionnelle" au cours de la méiose. A - représente un noyau de cellule diploïde contenant 3 paires de chromosomes (1 et 1 "- la 1ère paire de chromosomes homologues, 2 et 2" - la 2ème paire, 3 et 3 "- la 3ème paire). 2n \u003d 6,3 les chromosomes provenant de l'organisme maternel sont légers (nombres sans trait); leurs chromosomes homologues appariés provenant de l'organisme paternel sont noircis (nombres avec un trait). Les chromosomes homologues sont désignés par un nombre commun. Chaque chromosome est composé de deux chromatides. B - fusion de chromosomes homologues À ce stade, les chromosomes homologues échangent des sections séparées les uns avec les autres. En conséquence, une certaine redistribution du matériel héréditaire maternel et paternel entre les chromosomes (croisement) se produit. C - des filaments de fuseau se forment, se fixant aux chromosomes, la membrane nucléaire disparaît. D - les chromosomes homologues divergent vers des pôles opposés. cellules, aux pôles il y a un chromosome homologue de chaque paire, le nombre total de chromosomes à chaque pôle est la moitié de celui du noyau d'origine A. D - image il y a deux noyaux avec un ensemble haploïde de chromosomes dans chacun; l'un d'eux a eu plus de chromosomes de grand-mère (léger, maternel par rapport au noyau A) et moins grand-père (noirci, paternel par rapport au noyau A), dans l'autre - au contraire; ainsi, les nouveaux noyaux ne sont pas complètement identiques les uns aux autres dans la composition de leur substance héréditaire; leur différence est aussi due au croisement qui se produit avec les chromosomes au stade B.E - G - division mitotique de chacun des noyaux haploïdes D: division longitudinale de chaque chromosome, divergence des chromosomes vers les pôles, formation de deux noyaux haploïdes à partir de chaque noyau D. En conséquence, 4 cellules sont apparues avec des noyaux haploïdes au lieu d'une cellule avec un ensemble diploïde de chromosomes
Les organites ne sont pas répartis de manière strictement égale entre les cellules filles, mais leurs éléments constitutifs sont ensuite synthétisés dans les cellules, de nouvelles copies d'organelles de chaque espèce sont assemblées et leur nombre dans chaque cellule est rétabli à la normale. Les cellules elles-mêmes se développent. Dans les chromosomes, le matériel héréditaire est dupliqué, après quoi les chromosomes sont constitués, comme avant la division cellulaire, de deux chromatides. La cellule est prête pour une nouvelle division.
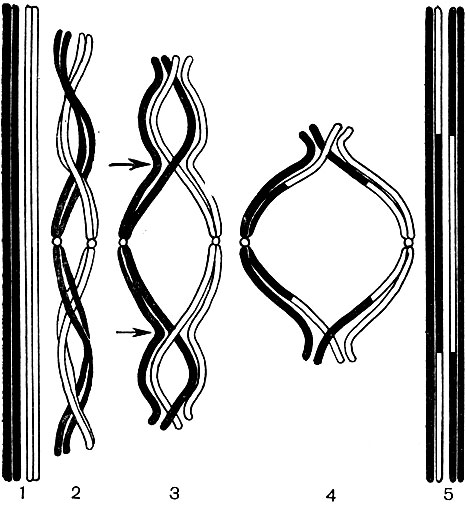
Figure: 38A. Schéma de croisement pendant la méiose au stade B de la Fig. 38. 1 - deux chromosomes homologues, dont chacun se compose de deux chromatides soeurs; 2 - les chromosomes homologues sont entrelacés les uns avec les autres; 3 - chaque chromosome est divisé en ses chromatides constituantes; les chromosomes commencent à se repousser et à diverger, mais à certains points (sur la figure - en quatre) ils restent connectés; les chromatides soeurs sont doublées dans la boucle centrale et les chromatides non soeurs dans les deux boucles extrêmes; les points où le franchissement est possible sont indiqués par des flèches; 4 - les chromatides ont échangé des zones, et les chromosomes, ayant subi une sorte d'hybridation mutuelle, divergent; 5 - deux chromosomes divergents, mais déjà "hybrides" (contrairement à ceux représentés sur 1)
Lorsque la génération diploïde de cellules est remplacée par une génération haploïde, la soi-disant division nucléaire de réduction, la méiose, se produit. Au cours de la méiose (Fig. 38), les chromosomes homologues de chaque paire se rapprochent, se rejoignent étroitement sur leur longueur et se tordent. Des sections séparées sont échangées entre les chromosomes homologues contigus. En conséquence, certains des gènes des chromosomes paternels sont transférés dans la composition des chromosomes maternels, et les gènes correspondants des chromosomes maternels occupent les places vacantes dans les chromosomes paternels (phénomène de croisement - Fig. 38A). Apparence ces chromosomes et d'autres ne changent pas en conséquence, mais leur composition qualitative devient différente. L'héritage paternel et maternel est redistribué et mixte. En outre, l'enveloppe nucléaire et le nucléole se dissolvent, un appareil à fuseau est formé, le même que pendant la mitose. Les chromosomes homologues sont séparés et, à l'aide de fils de broche, divergent vers les pôles de la cellule. Un pôle a un ensemble haploïde de chromosomes (un chromosome homologue de chaque paire), l'autre a un deuxième ensemble haploïde.
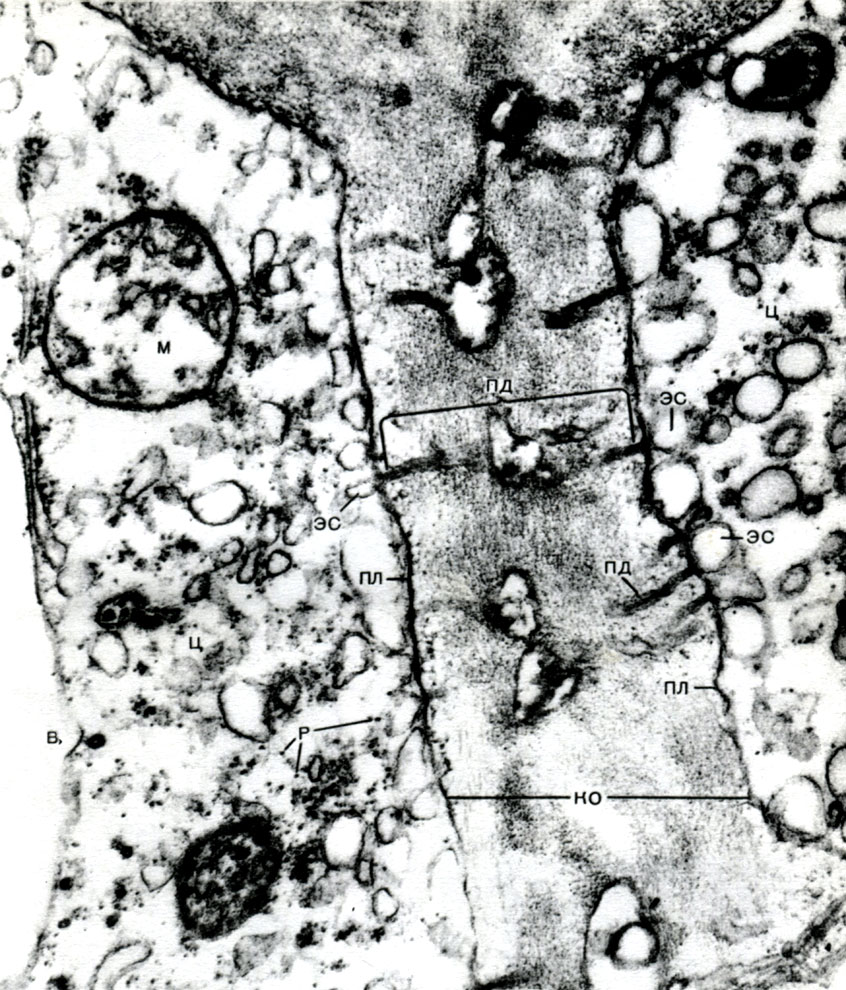
Après la méiose, les noyaux haploïdes formés se divisent par mitose. Dans ce cas, chaque chromosome de l'ensemble haploïde est divisé en deux chromatides, elles divergent et des cellules haploïdes filles se forment.
La méiose diffère de la mitose de deux manières fondamentales. Tout d'abord, lors de la méiose, chaque paire de chromosomes homologues s'agglutine et une partie du matériel héréditaire est échangée entre eux, ce qui n'est pas le cas lors de la mitose. Deuxièmement, pendant la méiose, les chromosomes homologues divergent vers les pôles de la cellule, un de chaque paire, et pendant la mitose, chaque chromosome est divisé en chromatides et les chromatides divergent vers les pôles, un de chaque chromosome. À la suite de la méiose, les cellules haploïdes sont formées à partir d'une cellule diploïde.
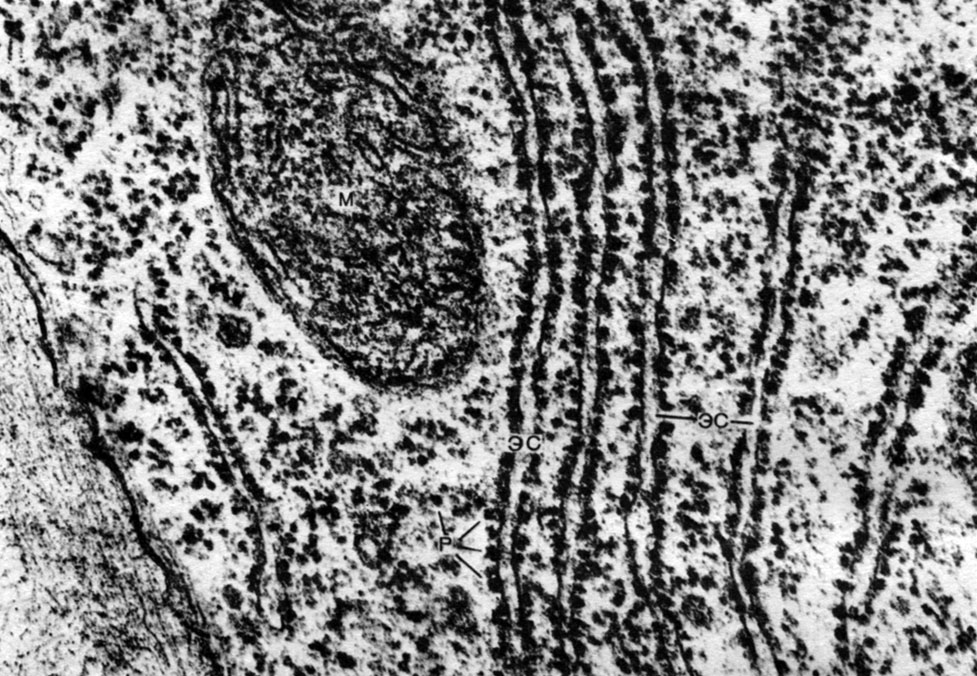
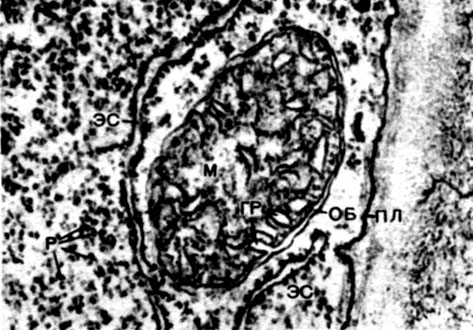
Tableau 10. Structure des cellules végétales. Appareil de Golgi dans le cytoplasme des poils racinaires en développement d'un radis (Raphanus sativus). Micrographie électronique (grossissement x 52000) par M.F Danilova: ag - appareil de Golgi, des coupes de sacs plats et de bulles sont visibles
Dans les cellules de certains tissus, au cours de leur développement, une mitose incomplète se produit: le matériel chromosomique dans les noyaux double, les chromosomes se divisent en deux, mais au lieu de former deux noyaux, ils restent dans le noyau d'origine. À partir de ce moment, il ne contient pas un diploïde, mais un ensemble tétraploïde (quadruple) de chromosomes. Le processus menant à cette duplication des chromosomes dans un noyau est appelé endomitose - mitose interne. Si cela arrive deux fois à un noyau, alors il devient un ensemble octoploïde (multiplié par huit), etc. Les cellules dont les noyaux portent plus de deux ensembles de chromosomes sont appelées polyploïdes, c'est-à-dire multiploïdes. La polyploïdie cellulaire augmente dans certains cas leur viabilité, puisque chaque gène est dupliqué par plusieurs autres gènes du même. Les gènes de qualité unique agissent à l'unisson et les dommages causés à l'un d'entre eux n'entraînent pas la perte du trait qu'il détermine, car il est compensé par le travail des gènes homogènes restants. Dans de nombreux cas, les cellules polyploïdes sont plus grandes et plus riches en contenu que les cellules diploïdes. Les variétés de plantes cultivées polyploïdes ont été sélectionnées, qui ont des qualités économiques accrues.
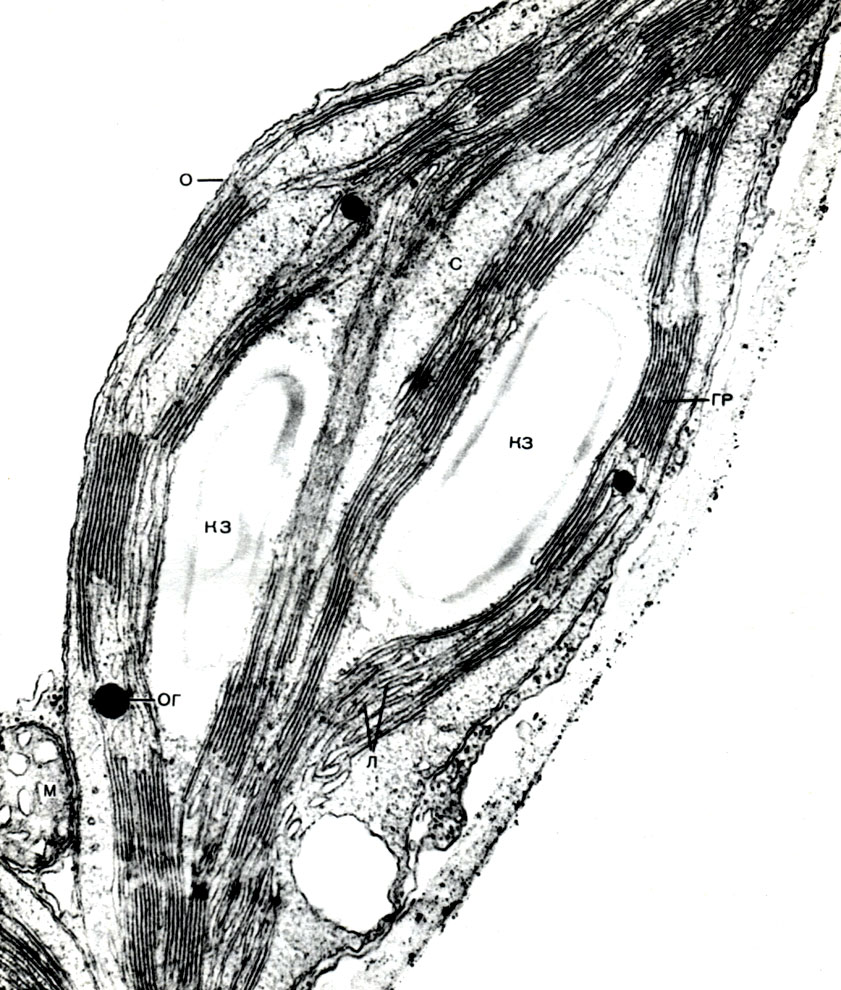
La science qui étudie la cellule s'appelle la cytologie. La cellule est la base de la structure de tous les êtres vivants et de tous les processus de la vie; par conséquent, la plupart des problèmes biologiques généraux les plus importants peuvent être résolus avec l'implication de la cytologie et des méthodes de ses recherches.
Avec l'aide de la cytologie, de nombreux problèmes pratiquement importants sont résolus: la lutte contre les maladies des plantes (fongiques, bactériennes), la sélection de nouvelles variétés de plantes cultivées, le dépassement de la stérilité (infertilité) des variétés hybrides.
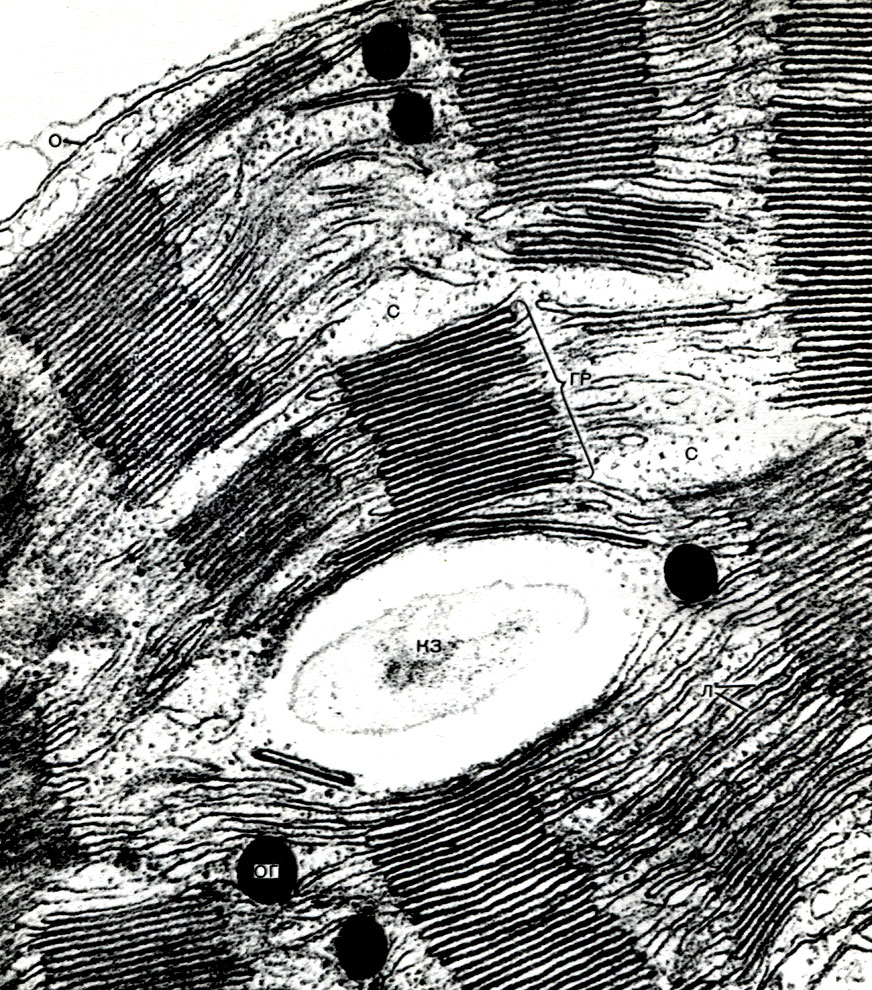
La cytologie moderne étudie ces problèmes en utilisant une variété de méthodes - microscopiques, biochimiques, biophysiques, génétiques - et en interagissant étroitement avec d'autres sciences biologiques connexes.