Cazul 2: Închiderea rădăcinii și crearea țesutului keratinizat. Pacientul este localizat. structura celulei vegetale, a țesuturilor, a organelor vegetative și de reproducere ... există un tip columnar (palisat) (Figura 9), spongios. Celulele aproape tuturor plantelor multicelulare nu sunt omogene, ci sunt colectate în T. În plantele inferioare T. ... un sistem (parenchim purtător de clorofilă, T. spongios și columnar) ... pe care se dezvoltă organismul plantei. La aceasta. Test tematic pe tema „Structura și funcția celulelor”, nota 10. Opțiunea 1. Partea A Într-un singur organism vegetal, toate celulele care au o structură similară și. gaz și pătrunde prin stomate către spațiile intercelulare ale țesutului spongios, și de acolo până la. Palisada (celule de sus, dens împachetate) și spongioase (de jos, slăbite. Slăbiciunea țesutului spongios joacă un rol important în schimbul gazelor frunzești. himerele plantelor) ca urmare a mutațiilor somatice este posibilă. formează un anumit model și formă, apoi cade, în timp ce. Dateiformat: PDF / Adobe Acrobat Iată țesutul educațional care dă naștere lamei frunzei și ... Țesutul spongios este țesutul principal, ale cărui celule au o formă rotunjită ... dacă faceți o adaptare simplă, așa cum se arată în imagine .. Întregul proces de respirație are loc în celulele organismului plantei. de la hrănirea naturală la laptele de vacă cu suplimente pe bază de plante .. Matrice pentru săruri de calciu din interiorul celulelor - lizozomi și mitocondrii .. În zonele cu var, se poate forma țesut osos și în jurul acestuia se va afla. Când este diagnosticată, nefrocalcinoza se distinge de rinichiul spongios.
ȚESUTUL SPONGOS (parenchim spongios), partea inferioară liberă a pulpei frunzei. Se caracterizează printr-o formă neregulată a celulei și spații intercelulare mari. țesătură din bumbac imprimat cu model oriental. [edit] ... în care un agent de contrast este injectat în țesutul spongios al procesului spinos ... țesut vegetal format din celule foarte asemănătoare cu un cambium. Figura prezintă o parte a secțiunii transversale a unei frunze de fistic cu un foarte. Celulele țesutului spongios de ventilație au adesea destul. Produsele secreției interne a organismelor vegetale sunt localizate sau în. Într-un singur organism vegetal, toate celulele sunt similare ca structură și. gaz și pătrunde prin stomate în spațiile intercelulare ale țesutului spongios, și de acolo până la. Țesutul este o colecție de celule care sunt similare ca structură și funcție .. celule ale parenchimului spongios; 5 - celule de pază ale stomatelor, un decalaj între fiecare pereche de ele. Citoplasma este localizată în toate celulele prezentate în figură. Celulele mesofile aparțin principalului parenchim purtător de clorofilă. Celulele țesutului spongios sunt mai mult sau mai puțin izodiametrice și, de regulă ,. Iată țesutul educativ care dă creșterea lamei frunzei și ... Țesutul spongios este țesutul principal, ale cărui celule au o formă rotunjită, ... dacă efectuați o adaptare simplă, așa cum se arată în figură .. Întregul proces de respirație are loc în celulele organismului plantei. Majoritatea celulelor vegetale au cavități - vacuole. Celulele mecanice ale țesutului au o membrană îngroșată. 15.… II - țesut spongios.… Schițați planta de cartof, indicând fructul și tuberculul din figură. Țesutul este un grup de celule care au o origine comună, efectuează una sau. Un exemplu al primului este clorenchima coloană, clorenchima spongioasă. Profesori Chestionare ale celor mai buni tutori. Găsiți un tutore Completați formularul de solicitare și dvs. Puneți o întrebare despre educația modernă, exprimați o idee sau rezolvați o problemă urgentă.
5 septembrie 2014. Probka, sau fellema (lat. Felema) este țesutul tegumentar secundar al axialului. Așa cum s-a menționat mai sus, celulele fellomului sunt depuse. În ele s-a găsit doar mezofilă uniform spongioasă. (omogen). (pe scară largă. Desenele anatomice sunt realizate cu ajutorul unui aparat de desen .. volumul unei celule, numărul de celule pe unitatea de suprafață a frondei. În toate plantele, aceste țesuturi sunt parenchimate în forma generală a celulelor lor. epidermă, 4 - țesut acvifer, 5 - parenchim coloanei, 6 - parenchim spongios ... pentru a face un desen schematic, marcând țesutul acvifer ,. desenarea copacilor cu un creion. cum să lipiți peste un ou cu spumă. Figura 1 - Elemente structura anatomică epiderma frunzei. Partea interioară a frunzei este umplută cu celule libere de țesut spongios. Țesutul spongios la astfel de pacienți a fost foarte des înlocuit pe toată lungimea sa. Celulele endoteliale conservate au prezentat semne de umflare. 5 septembrie 2014. Probka, sau fellema (lat. Felema) este țesutul tegumentar secundar al axialului. După cum sa menționat mai sus, celulele fella sunt depuse.Țesutul conjunctiv este format din diferite tipuri de celule. Figura 9.3.2.1. ... Țesutul osos spongios este format din elemente osoase subțiri. ȚESUTUL SPONGOS (parenchim spongios), partea inferioară liberă a pulpei frunzei. Se caracterizează printr-o formă neregulată a celulelor și spații intercelulare mari. Micrografie a substanței spongioase, care prezintă trabeculele sale (roz) și medulla (albastru). ... Țesutul osos spongios (substanță spongioasă, substanță trabeculară, lat. Celule, condroblast, condrocit. Celulele structurale nespecializate ale parenchimului sunt totuși active din punct de vedere metabolic: multe. Sistemul de aer intercelular în parenchimul spongios este dezvoltat în special. 5 .. Cifre în biologie. Arătate palisada (celule superioare, strâns împachetate) și spongioase .. formează un anumit model și formă, apoi cad, în timp ce. Celulele mesofile aparțin principalului parenchim purtător de clorofilă. Sunt cu pereți subțiri și. , sunt foarte slab aranjate ... Desen aleatoriu. Desenele au fost efectuate cu ajutorul unui aparat de desen RA-4M. Celulele epidermei superioare și inferioare ale farmaciei de mușețel de gradul "Karagandinskaya" sunt mici. În parenchimul spongios sunt recipiente cu ulei esențial. 30 ianuarie 2009. Acest lucru se observă când înlocuirea formelor vechi de celule ale osteocitelor și. Determinată pe modelul osos cu buclă mare de roentgenogramă ... Intern în raport cu osul spongios al procesului alveolar. Celulele parenchimului nespecializate structural sunt totuși active din punct de vedere metabolic: multe. Sistemul spațiilor intercelulare ale aerului din parenchimul spongios este dezvoltat în special. cinci. Desene în biologie.
Celula vegetală (N.I. Arronet)
O plantă, ca orice organism viu, este formată din celule și fiecare celulă este generată și de o celulă. Celula este unitatea cea mai simplă și obligatorie a unei ființe vii, este elementul ei, baza structurii, dezvoltării și a întregii activități vitale a organismului.
Există plante construite dintr-o singură celulă. Acestea includ alge unicelulare și ciuperci unicelulare. De obicei, acestea sunt organisme microscopice, dar există și organisme unicelulare destul de mari (lungimea algelor unicelulare acetabularia ajunge la 7 cm). Majoritatea plantelor pe care le întâlnim în viața noastră de zi cu zi sunt organisme multicelulare construite dintr-un număr mare de celule. De exemplu, într-o frunză a unei plante lemnoase există aproximativ 20.000.000 de ele. Dacă un copac are 200.000 de frunze (ceea ce este o cifră foarte realistă), atunci numărul de celule din toate acestea este de 4.000.000.000.000. Arborele în ansamblu conține încă 15 celule din nou. ...
Plantele, cu excepția unora inferioare, sunt formate din organe, fiecare dintre care își îndeplinește propria funcție în corp. De exemplu, la plantele cu flori, organele sunt rădăcină, tulpină, frunză, floare. Fiecare organ este de obicei construit din mai multe țesuturi. Țesutul este o colecție de celule similare ca structură și funcție. Celulele fiecărui țesut au propria lor specialitate. Efectuând lucrări în specialitatea lor, ele contribuie la viața întregii plante, care constă în combinație și interacțiune tipuri diferite munca diferitelor celule, organe, țesuturi.
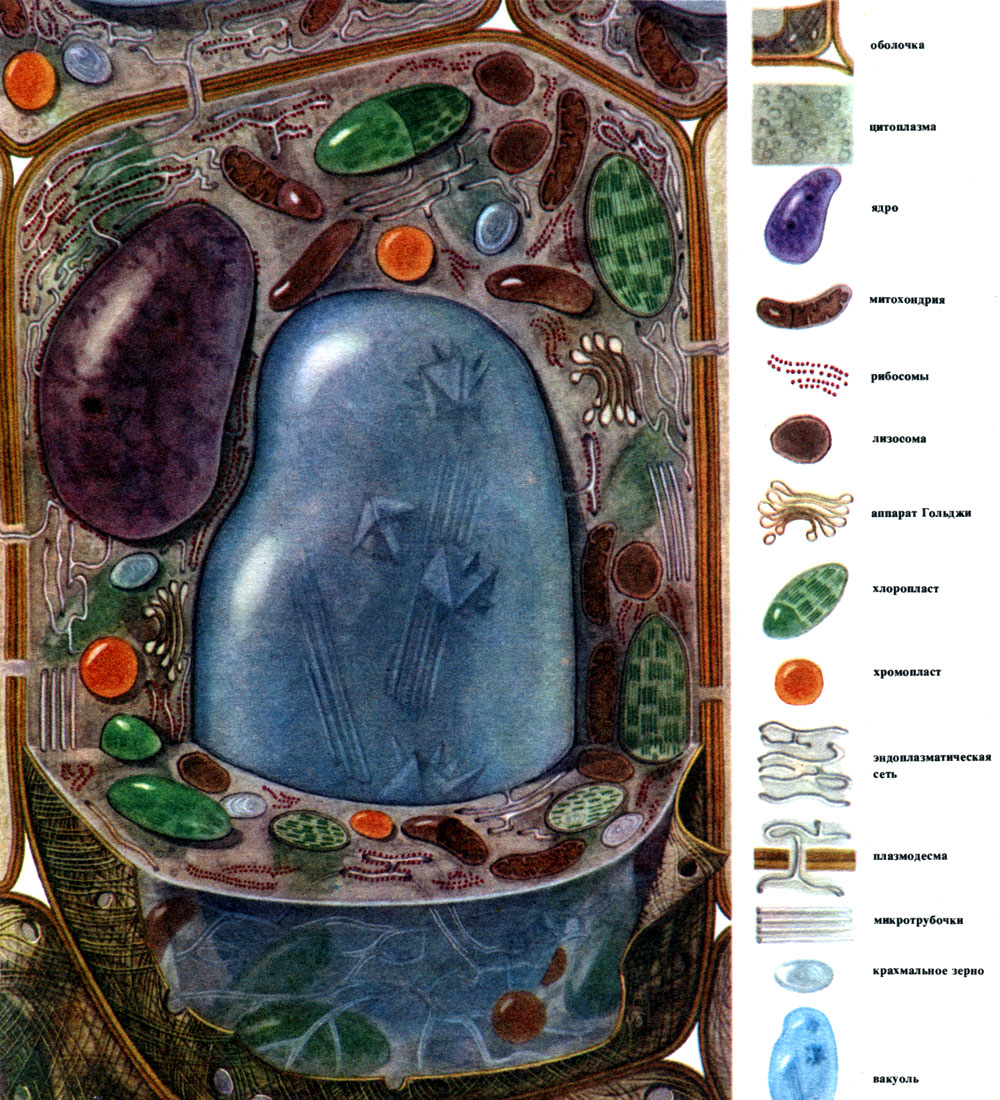
Principalele componente, cele mai frecvente din care sunt construite celulele, sunt nucleul, citoplasma cu numeroase organite de diferite structuri și funcții, coaja și vacuolul. Membrana acoperă exteriorul celulei, sub ea se află citoplasma, în ea se află nucleul și unul sau mai multe vacuole. Atât structura, cât și proprietățile celulelor diferitelor țesuturi diferă brusc datorită specializării lor diferite. Componentele principale și organele enumerate, care vor fi discutate în continuare, sunt dezvoltate în ele în grade diferite, au o structură inegală și, uneori, una sau alta componentă poate fi complet absentă.

Principalele grupuri de țesuturi din care sunt construite organele vegetative (care nu sunt direct legate de reproducere) ale unei plante superioare sunt următoarele: tegumentare, de bază, mecanice, conductoare, excretoare, meristematice. Fiecare grup include de obicei mai multe țesuturi cu o specializare similară, dar fiecare construit în felul său dintr-un anumit tip de celule. Țesuturile din organe nu sunt izolate unele de altele, ci constituie sisteme de țesuturi în care alternează elementele țesuturilor individuale. Deci, lemnul este un sistem de țesături mecanice și conductive, și uneori de bază.


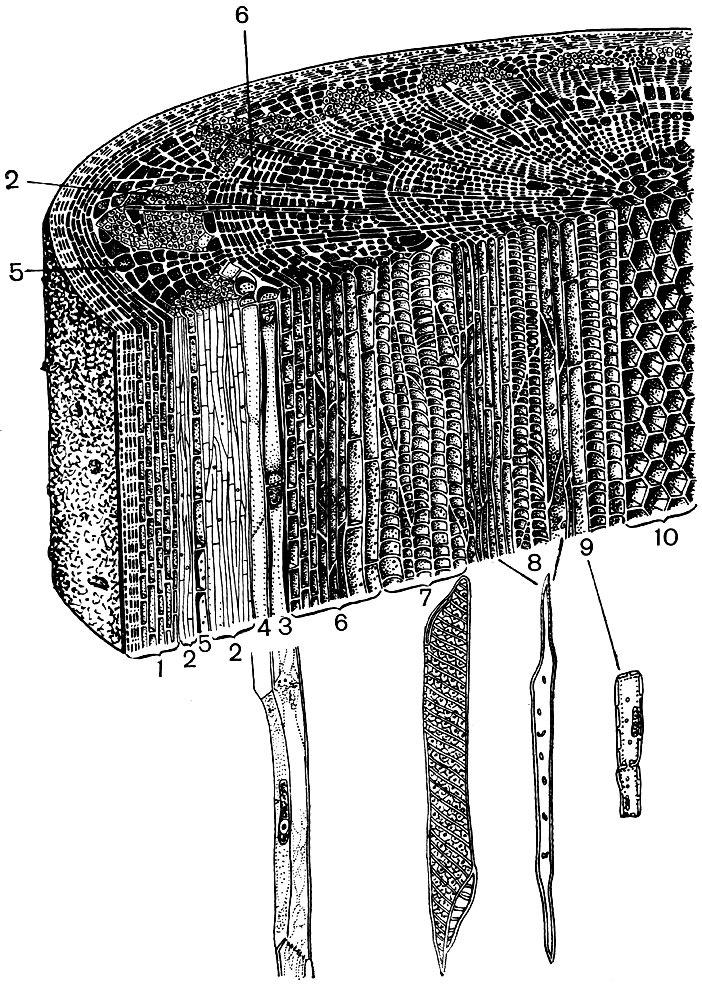
Luați, de exemplu, cele două organe ale unei plante superioare - frunza și tulpina. Și vom vedea cât de diverse sunt structura și funcționarea celulelor care își formează țesuturile (figurile 22 și 23). Vom vedea, de asemenea, modul în care structura celulelor fiecărui țesut este adaptată pentru a-și îndeplini funcțiile speciale.


Acoperirea țesuturilor - epidermă, plută, scoarță - protejează organele plantelor de efectele adverse: de la uscare, supraîncălzire, hipotermie, energie radiantă, deteriorări mecanice, umiditate excesivă, de la pătrunderea organismelor străine, cum ar fi bacteriile. Epiderma acoperă atât suprafețele frunzelor, lăstarii tineri, cât și petalele. De regulă, acesta este un strat de celule. Acei pereți celulari cu care se ating sunt de obicei sinuoși, iar proeminența unuia umple concavitatea celui vecin. Acest lucru promovează o conexiune puternică a celulelor, care, ca rezultat, formează un singur strat dens. Pereții exteriori ai celulelor care se învecinează cu aerul sunt mai groși.

Celulele epidermice secretă substanțe care se depun pe ele în exterior sub forma unui film (de exemplu, cutina, care formează o cuticula densă și ceară; cutina pătrunde, de asemenea, pereții exteriori ai celulelor epidermice). O înveliș exterior gros și un film exterior de cutină și ceară asigură proprietățile izolante și protectoare ale acestei țesături. Pătrunderea gazelor, a apei, a bacteriilor este dificilă prin stratul său. Cu toate acestea, aerul trebuie să intre în mod necesar în părțile verzi ale plantei, iar vaporii de apă și oxigenul trebuie să iasă afară. Schimbul de gaze, precum și de vapori de apă, între atmosferă și țesuturile interne ale organelor tinere, acoperite cu epidermă, se efectuează cu ajutorul stomatelor.
Stomele sunt formate din celule speciale ale epidermei, dintre care două (așa-numitele celule de pază ale stomatelor) există o deschidere de o dimensiune reglabilă - decalajul stomatal. Prin intermediul acestuia se realizează conexiunea dintre atmosferă și spațiile intercelulare aeriene cu grosimea frunzei sau a altui organ acoperit de epidermă. Celulele de închidere ale stomatelor sub influența unei modificări a condițiilor de umiditate sau de lumină își schimbă forma, închizându-se între ele sau deschizându-se. Procedând astfel, ele deschid sau, respectiv, închid decalajul stomatal. În lumină, când planta fotosinteză și are nevoie de un aflux de dioxid de carbon din atmosferă, golurile stomatale sunt deschise. Se închid noaptea; celulele de pază închid golurile stomatelor în timpul fierbinte al zilei, ceea ce protejează planta de o mare pierdere de apă, de ofilire.
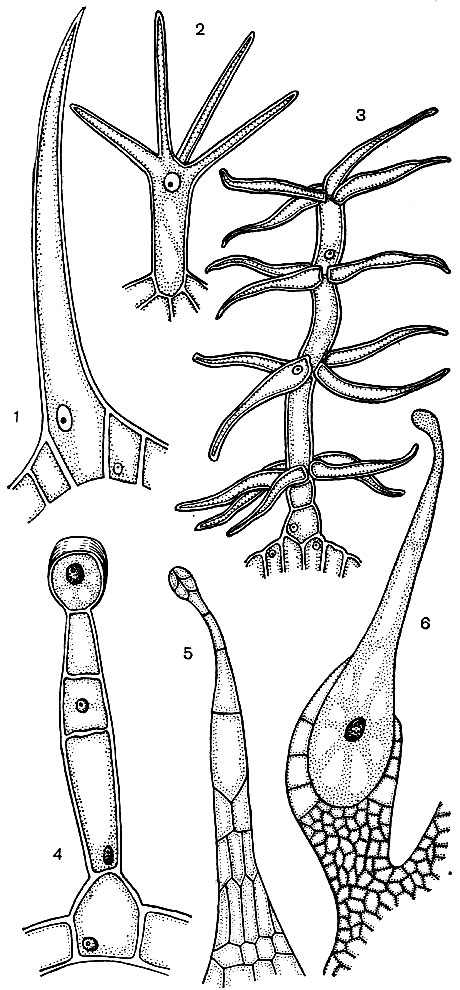
Celulele epidermice formează adesea excrescențe - fire de păr (Fig. 24). Uneori acestea sunt formațiuni multicelulare, în alte cazuri fiecare păr este un proces al unei celule care se află în stratul comun al epidermei. Acestea joacă un rol protector, de susținere (de exemplu, la plantele cățărătoare sau târâtoare), excretor. Firele de rădăcină sunt importante - excrescențe tubulare ale celulelor epidermice ale rădăcinii. Măresc suprafața de aspirație a acestuia din urmă. Se estimează că se dezvoltă până la 14 miliarde de fire de rădăcină pe 14 milioane de rădăcini laterale ale unei plante de secară. Suprafața rădăcinilor este de 232 m 2, iar suprafața firelor de păr este de 400 m 2. Epiderma multor semințe sau fructe formează fire de păr care favorizează dispersarea semințelor și, astfel, a plantelor. Adesea o persoană folosește fire de păr. Un exemplu în acest sens este bumbacul, care reprezintă firele epidermice ale semințelor de bumbac; firele de păr au învelișuri groase de celuloză și ating o lungime de 70 mm.
În tulpina plantelor perene de sub epidermă, pentru a o înlocui, se dezvoltă un țesut protector mai grosier - un dop. Celulele sale mor, pierd protoplastul și constau doar din pereți groși care înconjoară o cavitate plină cu aer sau substanțe rășinoase. Pereții sunt impregnați cu suberină, ceea ce îi face impermeabili și etanși, termoizolanți, precum și puternici și rezistenți. De asemenea, se pot lignifica. Un exemplu al acestei țesături este scoarța de mesteacăn. Este destul de subțire, dar pluta de plută atinge o grosime de câteva zeci de centimetri. Pluta este întreruptă în unele locuri de lenticele - formațiuni ale unui alt țesut permeabil la apă și aer. Prin linte, părțile interne ale trunchiului sunt schimbate cu mediul înconjurător. Lintea se dezvoltă la nivelul stomatelor.
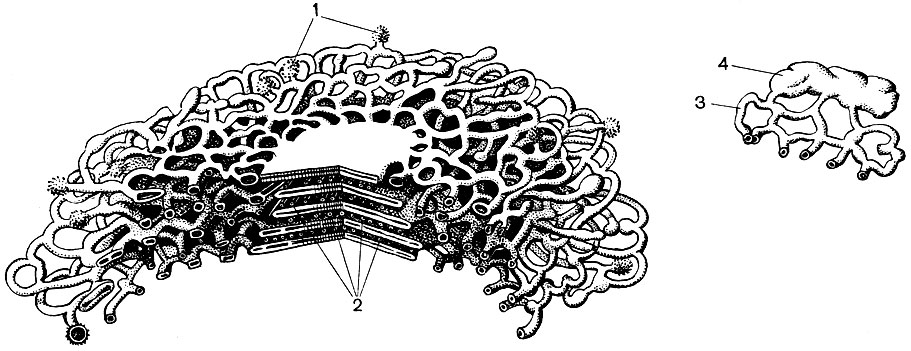
Celulele Bast sunt localizate în tulpină sub țesuturile tegumentare. Bast este un sistem de elemente din mai multe țesături: conductiv, de susținere, de bază. Cel mai important element al bastului este tuburile de sită. Sunt construite din celule alungite, alungite de-a lungul tulpinii, articulate între ele la capete. Acestea sunt celule vii, dar fără nuclee, elementele citoplasmei în care se află lângă perete. Cojile de la joncțiunea acestor celule au numeroase găuri, astfel încât pereții despărțitori sunt ca o sită. Ca urmare, celulele adiacente comunică între ele și se dovedesc astfel unite în tuburi lungi care se întind prin vene și pețiolele frunzelor, de-a lungul tulpinii și rădăcinii.
Prin sistemul de tuburi de sită, produsele formate în părțile verzi ale plantelor se deplasează către toate celelalte părți ale sale și le alimentează. Principalul produs transportat este zaharoza. Tuburile de sită sunt un element al țesutului conductiv. Există celule satelit lângă celulele tuburilor de sită. Acestea conțin nuclee, iar protoplastul lor are legături directe cu citoplasma celulelor de sită non-nucleare. În bast, există, de asemenea, zone constând din celule parenchimatoase (adică, celule a căror lungime, lățime și înălțime sunt de dimensiuni apropiate). În ele se depun diverse substanțe - amidon, uleiuri, rășini. Acestea sunt celulele țesutului subiacent.
În plus, bastonul conține elemente ale țesăturii de susținere - fibre bast. Acestea sunt celule foarte lungi, cu pereți îngroșați; lungimea unei celule poate depăși lățimea ei de mii de ori. La in, lungimea fibrelor subțiri alungite de bast ajunge la 4 cm, iar la ramie - chiar și 35 cm. De obicei, acestea sunt celule moarte, fără protoplast. Acestea îndeplinesc o funcție mecanică, creând puterea tulpinii.
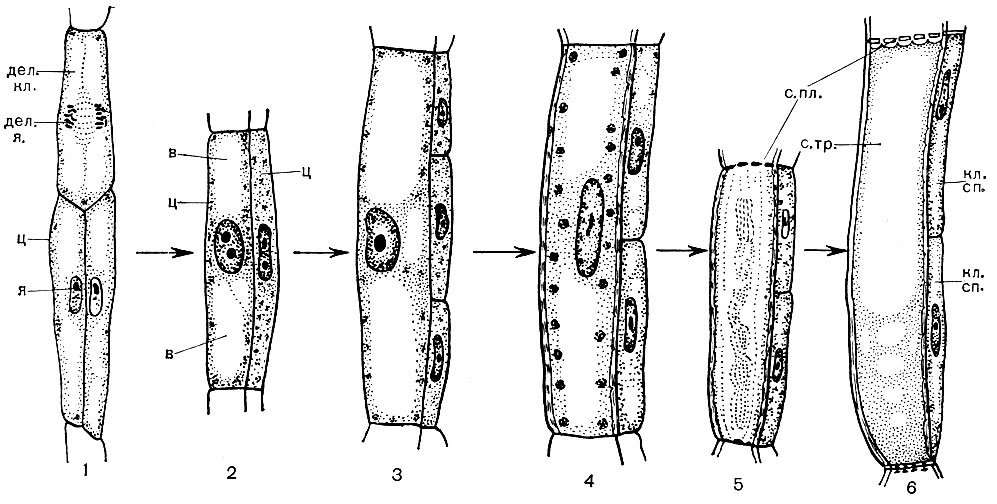
Majoritatea celulelor specializate sunt incapabile de reproducere. Cu toate acestea, planta crește pe tot parcursul vieții sale și se formează noi celule pe parcursul vieții sale. Se dezvoltă din celulele țesuturilor educaționale - meristematice. Reproducerea prin diviziune este o specializare a celulelor meristematice, funcția lor în organism. Celulele care provin din acestea se dezvoltă, transformându-se într-una sau alta celule specializate - celule de tuburi de sită, celule de dop, fibre etc. Țesutul educațional este situat în diferite părți ale plantei (de exemplu, în punctele de creștere - în vârfurile lăstarilor, rădăcinilor). De obicei, există mai multe straturi de țesut educativ în grosimea tulpinii. Sub stratul de plută există celule felogen, care, împărțind, completează numărul de celule de plută și crustă. Un strat de celule cambium este situat în interiorul bastului. Cele din celulele pe care le generează, care se formează în afara cambiului, se dezvoltă în celule bast (Fig. 25). Odată ajuns în interior, adică mai aproape de axa trunchiului, dau naștere celulelor lemnoase.
Lemnul este format din elemente de țesături conductive, de susținere și de bază. Primele includ fibre de lemn - celule moarte îndelungate cu pereți lignificați; la a doua - vase, care sunt rezultatul fuziunii mai multor celule; la a treia - celule ale parenchimului lemnos. Celulele care diferențiază de elementele cambiale în celule vasculare cresc în lungime și lățime. Pereții lor laterali sunt îngroșați și lignificați. În pereți, totuși, rămân porii, închiși doar de o coajă subțire. Partițiile de la joncțiunile celulelor adiacente dispar, protoplastul moare. Ca urmare, se formează vase lungi, formate din niște pereți. Sistemul conductiv se extinde prin rădăcină și tulpină în frunză. Prin astfel de vase, un flux ascendent de apă și săruri dizolvate în acesta este efectuat de la rădăcini la toate organele plantei.
În centrul tulpinii sunt celule de bază - celule parenchimale rotunde sau poliedrice. Acestea sunt elementele țesăturii principale. Uneori sunt goale și conțin aer; uneori sunt umplute cu substanțe nutritive de rezervă, diverse cristale, taninuri. Pereții lor pot fi lignificați. Razele radiale pătrund în lemn și în bast. Celulele lor sunt derivați de cambium și au o funcție de stocare. La nivelul bastului, aceste raze se extind în mod vizibil.
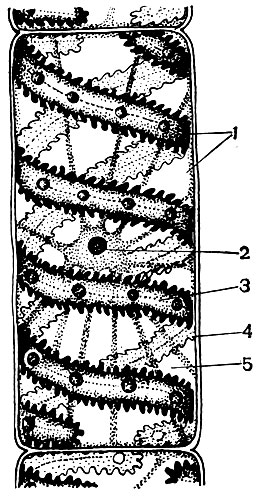
În pulpa frunzei dintre epiderma superioară și inferioară, există țesutul principal - celule cu membrane subțiri și un număr mare de plastide verzi - cloroplaste. Fotosinteza are loc în aceste celule. Straturile superioare sunt compuse în mare parte din celule alungite, strâns adiacente una cu cealaltă - acest parenchim coloanei. Straturile inferioare se slăbesc între celule, există spații intercelulare - spații umplute cu aer; este un parenchim spongios. Parenchimul este pătruns cu vene ramificate, care sunt construite din mănunchiuri de țesuturi mecanice (diverse fibre) și conductive (tuburi de sită și acvifere).
![]()
Această prezentare scurtă și schematică a planului structurii tulpinii, frunzei arată cât de diverse sunt dimensiunile, forma, structura, funcțiile celulelor aceleiași plante. Dacă luați diferite plante, atunci vă puteți întâlni cu o varietate și mai mare de celule. Cu toate acestea, de regulă, celulele țesuturilor cu același nume, chiar și la plantele care sunt foarte îndepărtate una de alta, sunt similare, deoarece aceste celule îndeplinesc funcții similare. Celulele epidermice ale unei frunze de mesteacăn și, de exemplu, păpădia sunt mai asemănătoare între ele decât celula epidermică a unei păpădii și a celulei sale. Trăsăturile distinctive ale celulei sunt asociate în primul rând cu funcția sa specială. Celulele specializate în funcția mecanică au membrane îngroșate și adesea lignificate. Specializarea în fotosinteză duce la apariția cloroplastelor în celule. Specializarea în funcția de conducere este asociată cu alungirea celulelor, cu pierderea protoplastului, cu modificări ale membranelor celulare la joncțiuni, datorită cărora comunică cavitățile celulelor adiacente. Celulele specializate în funcția de protecție se caracterizează prin diferite modificări ale pereților exteriori, prezența firelor de păr și capacitatea de a produce substanțe de protecție.
Specializarea în funcția de stocare a substanțelor nutritive poate duce la o creștere a dimensiunii celulelor, la apariția vacuolelor foarte mari în ele. În celulele meristematice, care se împart intens, sunt dezvoltate în special acele structuri intracelulare care asigură sinteza substanțelor - părțile constitutive ale celulei.
În funcție de natura specializării, unele caracteristici ale structurii și activității celulei sunt foarte puternic dezvoltate, în timp ce altele, dimpotrivă, sunt puțin dezvoltate sau complet absente. După cum am văzut, unele celule, devenind specializate, mor și este în starea moartă când încep să își îndeplinească funcția specială într-un organism multicelular (celule de plută, fibre de lemn, vase).
Celulele algelor filamentoase multicelulare și ale plantelor unicelulare sunt foarte diverse și deosebite (figurile 26 și 27). Mai mult, celula oricăreia dintre acestea din urmă este foarte diferită de celulele plantelor multicelulare. Ea singură trebuie să îndeplinească mai multe funcții, care la plantele multicelulare sunt împărțite între celule din diferite țesuturi.
În același timp, chiar și celule foarte diferite au asemănări profunde în structură și funcție. Pentru organismele multicelulare, acest lucru se datorează, în primul rând, faptului că toate celulele corpului (dacă excludem reproducerea vegetativă, în care un întreg complex multicelular este separat imediat de planta mamă ca un individ nou sau rudimentul său) sunt descendenți ai aceleiași celule progenitoare. Prin urmare, indiferent cât de specializate sunt celulele, ele au o origine comună și, prin urmare, sunt rude.
În al doilea rând, trăsăturile comune în structura celulelor plantelor din diferite specii sunt asociate cu faptul că toate plantele sunt legate într-un grad sau altul. Toate plantele au evoluat prin evoluție din strămoșii unicelulari comuni. Caracteristicile comune sunt moștenite de celulele plantelor moderne de la celulele antice progenitoare. Acest lucru este, de asemenea, asociat cu prezența trăsăturilor comune ale structurii și activității celulelor vegetale și animale.
În al treilea rând, asemănarea este asociată cu faptul că toate celulele vii, indiferent de funcția specială pe care o pot îndeplini în corp, trebuie să își asigure în primul rând propria viață. Celulele absorb substanțele nutritive, le prelucrează, câștigă energie și își construiesc propriul corp, respiră, scapă de substanțe inutile, luptă împotriva diferitelor daune, reacționează la schimbările din condițiile externe, reconstruind activitatea vitală și cresc. Toate aceste procese în diferite celule se desfășoară într-un mod similar și cu ajutorul unor structuri de același tip, care sunt comune în ceea ce privește structura nu numai pentru celule de plante diferite, ci și pentru celule de plante și animale. Trebuie spus că performanța oricărei funcții speciale a unei celule se bazează pe proprietățile sale generale. Una sau alta caracteristică, o abilitate inerentă tuturor celulelor, se dezvoltă deosebit de puternic într-o celulă specializată și asigură că celula își îndeplinește funcția principală, specială. Aceste caracteristici comune, fără de care este imposibil să îndeplinești această funcție specială, sunt păstrate în celulă, iar restul se pot pierde. Celulele specializate moarte sunt un caz extrem al acestui lucru. Funcția specială a acestor celule este legată de membrana lor; protoplastul este necesar numai atâta timp cât creează o coajă; după aceea, se stinge și întreaga celulă constă numai dintr-o coajă neînsuflețită, care funcționează pentru nevoile plantei.
Având în vedere caracteristicile generale ale structurii și activității vitale a celulelor vegetale, este convenabil să vorbim despre o anumită celulă tipică care a absorbit trăsăturile comune ale diferitelor celule specializate. O astfel de celulă, deși nu există într-o „formă pură”, poate fi chiar descrisă (Tabelul 7, Fig. 28). Dintre celulele reale, acestea sunt mai asemănătoare cu o astfel de celule „tipice” ale parenchimului frunzelor (Tabelul 8, Fig. 29).
Să trecem la descrierea structurii și lucrării unei celule vegetale.
Tabelul 5. Angiospermele: stânga sus - floarea magnoliei; sus dreapta - flori de măr; mai jos este un lotus.
Tabelul 6. Angiospermele: sus (de la stânga la dreapta) - clopote, pelargoniu, șofran (crocus); jos stânga - aster; în dreapta jos - brusture.
În exterior, celula vegetală este acoperită cu o membrană care nu este aceeași ca grosime și structură în diferite celule. Substanțele care o formează sunt produse în citoplasmă și depozitate în afara acesteia, creând treptat o coajă. Aceste substanțe sunt în principal polizaharide cu molecule mari - pectină, hemiceluloză și celuloză în cantități mici. Ele formează așa-numita coajă primară. Este destul de elastic, deoarece celula crește, se întinde și crește și, prin urmare, nu interferează cu creșterea celulei. Cu toate acestea, creează o anumită rezistență a celulei și este capabilă să o protejeze de deteriorarea mecanică. Există celule cărora le lipsește o astfel de membrană primară - acestea sunt unele celule care servesc la reproducere asexuată și sexuală (zoospori și gamete de alge și ciuperci inferioare, gameti masculi de plante superioare). Multe celule au nu numai o membrană primară, ci și secundară. Se formează sub primar și este construit în principal din celuloză. Celuloza este o polizaharidă ale cărei molecule formează cele mai fine filamente - microfibrilele. În înveliș, filamentele de celuloză sunt scufundate într-o substanță amorfă formată din compuși de pectină. În unele celule, aceste microfibrile de celuloză sunt situate pe lungimea celulei, în inele; din această cauză, astfel de celule se pot întinde în lungime, dar nu și în lățime (de exemplu, celulele vaselor stem). Alții au fire longitudinal; celulele cu o astfel de membrană sunt elastice atunci când sunt întinse, dar foarte rigide în întinderea longitudinală. În altele, acestea sunt situate oblic, formând o spirală (fire de păr epidermice din semințe de bumbac, fibre de bast). Toate acestea seamănă cu structurile din beton armat, iar firele de celuloză joacă rolul tijelor de fier aici, iar substanțele pectinice joacă rolul cimentului. Celulele cu membrană secundară sunt foarte durabile. Ele formează țesuturile mecanice de susținere ale plantei. Uneori, membrana secundară joacă, de asemenea, rolul unui depozit de produse nutritive: substanțele care o formează pot fi transformate în altele mai simple, care sunt consumate ca alimente.
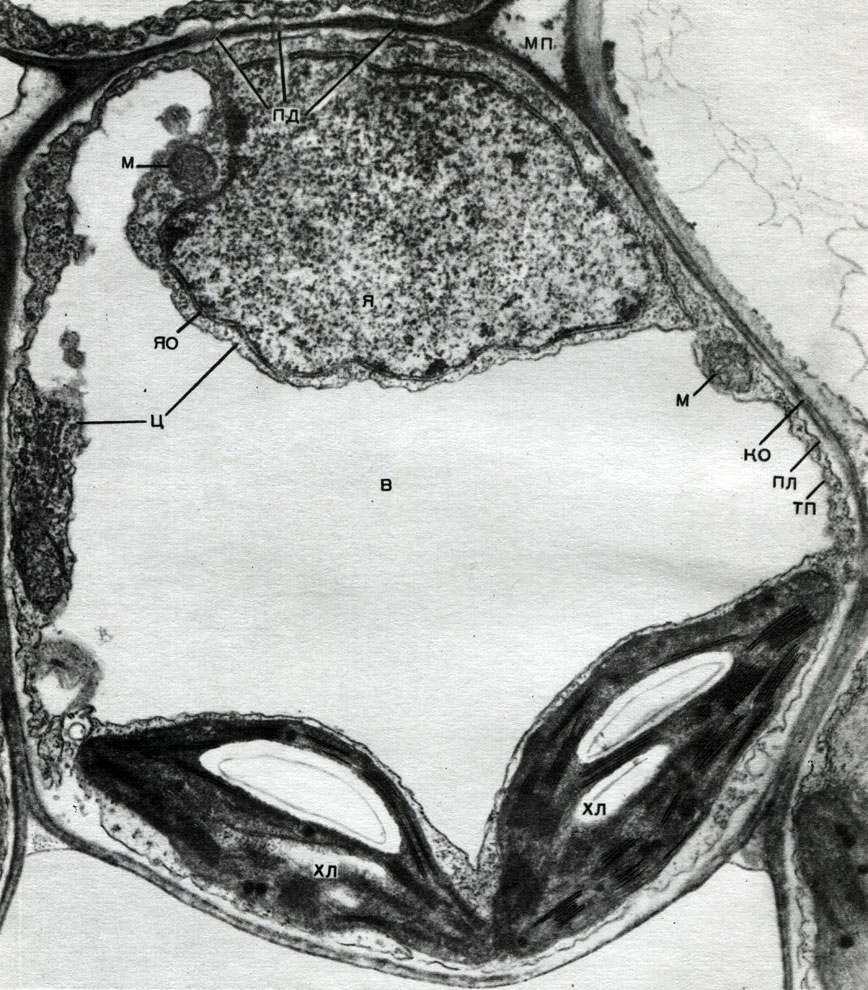
În cochilie există locuri neîngroșate - pori (în cochilia primară, se numesc câmpuri de vizuină), prin care se realizează comunicarea între celulele vecine. Corzi subțiri de citoplasmă trec prin câmpurile porilor și porii. Acestea sunt plasmodesme care leagă citoplasma celulelor învecinate (Tabelul 9). Acestea schimbă substanțe între celulele vecine. Plasmodesmata, împreună cu elemente ale țesutului conducător, conectează celulele și țesuturile corpului într-un singur întreg.
Schimbul de substanțe și răspândirea excitării permit celulelor să influențeze dezvoltarea și munca reciprocă, iar fiecare țesut afectează funcțiile vitale ale tuturor celorlalte țesuturi. Acest lucru creează coordonarea muncii tuturor părților unui singur organism, a întregii plante.
În multe celule, membrana celulară cu vârsta este impregnată cu substanțe care o consolidează și mai mult. Acumularea de lignină în ea duce la lignificarea cojii. Pereții celulelor lemnoase sunt lignificate și, adesea, fibre de bast, coji de celule de piele de semințe, uneori pericarp (Fig. 30) și chiar petale. Celulele cu pereți lignificați nu sunt doar mai puternice, ci și mai puțin permeabile microbilor și apei. Membranele altor celule - țesuturile tegumentare, precum și pe suprafețele rănilor - devin plute, îmbibate în suberină. Conținutul celulelor se stinge atunci, deoarece suberina este impermeabilă la apă și gaze, dar țesutul din astfel de celule moarte protejează părțile vii interioare ale plantei de influențe externe dăunătoare. Așa cum s-a menționat deja, membranele exterioare ale celulelor epidermice sunt de obicei impregnate sau acoperite cu cutină și ceară și acest lucru menține celulele împotriva evaporării excesive a apei, a infecției. Carcasa poate fi, de asemenea, impregnată cu minerale, în principal săruri de calciu.
Cu toate acestea, există momente în viața celulelor în care membrana lor ar trebui să devină mai puțin durabilă decât înainte. Acest lucru se întâmplă, de exemplu, în timpul germinării semințelor.
Membrana celulei polizaharidice este o trăsătură caracteristică a structurii unei celule vegetale, care o distinge de o celulă animală.
Membrana sau peretele celular este o formațiune de protecție. Există citoplasmă sub membrană. Stratul său cel mai exterior, adiacent membranei, este membrana celulară de suprafață - plasmalema. Este o combinație de straturi de molecule asemănătoare grăsimilor și proteinelor. Astfel de membrane se numesc membrane lipoproteice („lipos” - grăsime, „protein” - proteină). O membrană de acest design delimitează citoplasma de vacuole; această membrană se numește tonoplast. Multe organite celulare sunt construite din membrane lipoproteice. Cu toate acestea, în fiecare caz, membrana este construită din substanțe asemănătoare grăsimilor (lipide) și proteine \u200b\u200binerente acestei membrane particulare. Diversitatea calitativă a lipidelor și, în special, a proteinelor este colosală, de unde marea varietate de membrane care diferă prin proprietăți, atât în \u200b\u200bcadrul aceleiași celule, cât și în celule diferite.
Plasmalemma reglează intrarea substanțelor în celulă și ieșirea lor din aceasta, asigură pătrunderea selectivă a substanțelor în și din celulă. Rata de penetrare prin membrana diferitelor substanțe este diferită. Apa și substanțele gazoase pătrund bine prin ea. Substanțele liposolubile pătrund, de asemenea, cu ușurință, probabil datorită faptului că are un strat lipidic. Se presupune că stratul lipidic al membranei este pătruns cu pori. Acest lucru permite substanțelor insolubile în grăsimi să treacă prin membrană. Porii poartă o sarcină electrică, astfel încât pătrunderea ionilor prin ele nu este complet liberă. În unele condiții, sarcina porilor se schimbă, iar aceasta reglează permeabilitatea membranelor pentru ioni. Cu toate acestea, membrana nu este la fel de permeabilă la diferiți ioni cu aceeași sarcină și la diferite molecule neîncărcate de dimensiuni apropiate. Aceasta este manifestarea celei mai importante proprietăți a membranei - selectivitatea permeabilității acesteia: pentru unele molecule și ioni este mai bine permeabilă, pentru altele este mai proastă.
O substanță se deplasează în celulă, difuzându-se în ea, dacă concentrația sa în exterior este mai mare decât concentrația din interior. În caz contrar, ar trebui să difuzeze în afara celulei. Dar, după ce au pătruns în celulă, moleculele acestei substanțe pot fi implicate imediat în metabolism și pot fi transformate în alte substanțe. Apoi, concentrația acestei substanțe în celulă scade din nou și o nouă porțiune a moleculelor sale se difuzează în celulă. Drept urmare, celulele care consumă în mod activ această substanță sau alta, acestea din urmă intră intens din celulele din jur.
Un alt mecanism pentru absorbția sporită a unei substanțe de către o celulă este legarea acesteia de către proteine \u200b\u200bsau alte substanțe. Prin legare, substanța părăsește soluția intracelulară și nu interferează cu difuzarea ulterioară a aceleiași substanțe. Continuă să intre în celulă și se acumulează în ea într-o formă legată. Deci, în celulele unor alge marine, iodul se acumulează în concentrații de un milion de ori mai mari decât concentrația sa în apa de mare.
În toate aceste cazuri, vorbim despre mișcarea substanțelor prin difuzie. Cu toate acestea, membranele celulare au și mecanisme pentru deplasarea substanțelor prin ele însele de la o concentrație mai mică la una mai mare. Printre proteinele membranei, există substanțe proteice, a căror activitate este de a muta diferite substanțe de pe o parte a membranei pe cealaltă. Se numesc enzime de transport. Probabil, o astfel de enzimă de transport formează un intermediar cu substanța transportată; acesta din urmă trece prin membrană, apoi se descompune în părțile sale originale, după care enzima se întoarce înapoi, iar substanța transferată rămâne de cealaltă parte a membranei. Detaliile acestui proces sunt necunoscute, dar chiar și faptul unui astfel de transport activ de substanțe este fără îndoială. Spre deosebire de transportul difuz, pasiv, enzimatic necesită cheltuieli de energie. Probabil, tocmai datorită acestui transport activ de ioni, celulele epidermei radiculare sunt capabile să absoarbă substanțele anorganice necesare plantei din sol și apoi să le transfere prin plantă din celulă în celulă.
Odată cu modificările chimice sau fizice din mediul extern și intern, membranele celulare își schimbă permeabilitatea, precum și gradul și natura selectivității sale. Mecanismele de reglare a mișcării substanțelor în și din celulă se bazează pe aceasta. O modificare a permeabilității membranelor pentru nutrienți se reflectă în intensitatea proceselor metabolice din celulă, asupra naturii sintezelor care apar în ea și pe parcursul întregii sale vieți. Modificarea permeabilității membranelor pentru ionii de sodiu și potasiu este mecanismul de propagare a excitației celulare - apariția și mișcarea biocurentului. Un număr mare de cationi, în principal ioni de potasiu, sunt atașați la suprafața membranei. Prin urmare, poartă o încărcătură pozitivă din exterior. Sub influența stimulilor, ionii de potasiu sunt separați de zona afectată, sarcina zonei scade și devine electronegativă față de zonele învecinate ale membranei. Această electronegativitate este, la rândul său, un iritant pentru zonele învecinate, în același mod îndepărtându-și sarcina, depolarizându-le. Unda de electronegativitate se răspândește pe membrană - acesta este curentul biologic. Apoi ionii de potasiu stau din nou pe membrană, dându-i sarcina inițială - o undă de depolarizare este urmată de o undă de restaurare. Suprafețele plasmodesmelor care trec prin pereții celulari și care leagă citoplasma celulelor vecine sunt, de asemenea, formate din astfel de membrane. De asemenea, biocurentul se mișcă de-a lungul lor, răspândindu-se de la celulă la celulă. Biocurenții celulelor vegetale sunt încă slab înțelese. Cu toate acestea, este clar că acestea sunt o metodă de semnalizare utilizată în celula vegetală pentru a declanșa unele reacții chimice și a inhiba altele. Faptul este că toate procesele chimice care apar în celulă și care constituie esența activității sale vitale sunt efectuate cu participarea catalizatorilor biologici - enzime. Fiecare reacție este posibilă numai atunci când enzima care o declanșează este activă. Majoritatea enzimelor devin active sub acțiunea anumitor cationi anorganici: K., Na. , Ca .., Mg .., Mn ... Biocurentul, modificând permeabilitatea membranei pentru substanțele încărcate, creează condiții pentru pătrunderea lor prin membrane și contactul cu enzimele. Astfel, biocurentul activează anumite enzime și astfel reglează și direcționează metabolismul în celulă. Reglarea metabolismului de către biocurenți este doar una dintre multele modalități de a regla metabolismul intracelular.
Ceea ce s-a spus despre permeabilitatea membranei de suprafață a celulei - plasmalema - se aplică și altor membrane intracelulare, inclusiv celor din care sunt construite multe organite celulare.
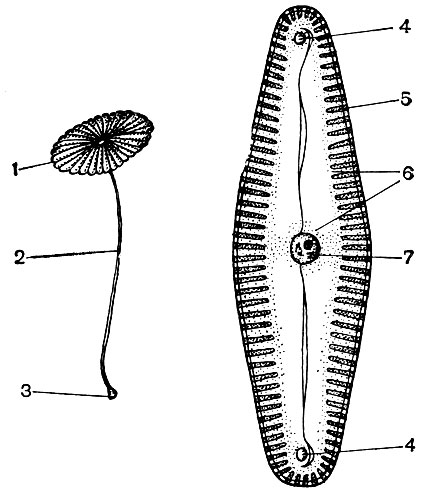
Citoplasma, considerată odată o soluție coloidală omogenă de substanțe proteice, este de fapt structurată complex. Odată cu dezvoltarea tehnicilor de cercetare microscopice, au devenit clare detalii din ce în ce mai subtile ale structurii citoplasmei. În citoplasmă, au fost descoperite diferite organite (organite) - structuri, fiecare dintre acestea îndeplinind anumite funcții fiziologice și biochimice. Cele mai importante organite ale citoplasmei sunt mitocondriile, reticulul endoplasmatic (reticulul endoplasmic), aparatul Golgi, ribozomii, plastidele, lizozomii. Celulele motile (zoospori și gamete de alge, spermatozoizi de coadă de cal, ferigi, cicade, unele alge unicelulare și coloniale) au organite de mișcare - flageli.
În special o mulțime de fapte noi despre structura fină a citoplasmei au adus și sunt produse prin utilizarea unui microscop electronic, care permite studierea detaliilor structurii organelor. Metodele biofizice și biochimice moderne fac posibilă izolarea anumitor organite ale citoplasmei în formă pură și apoi studierea lor compoziție chimică și funcțiile lor. În afara celulei, în medii cu compoziție complexă, multe organite sunt capabile să facă treaba pe care o fac atunci când sunt în celulă.
Partea citoplasmei în care sunt scufundate organitele și care până acum pare fără structură se numește substanța de bază a citoplasmei sau hialoplasmei. Hialo-plasma nu este în nici un caz un material de umplutură pasiv, ci o parte activă a citoplasmei. O serie de procese chimice vitale au loc în ea, conține multe proteine \u200b\u200benzimatice, cu ajutorul cărora se efectuează aceste procese.
Viața celulară constă într-o muncă chimică continuă, care în totalitate se numește metabolism. În esență, celula este o fabrică chimică care produce o gamă largă de produse și extrage independent energia necesară pentru ao produce. Produsele sale sunt substanțe de care ea însăși are nevoie pentru a-și menține propria viață (pentru a-și construi corpul în timpul creșterii și dezvoltării, pentru a-și înlocui părțile uzate) și pentru a crea celule fiice în timpul reproducerii și pentru nevoile altor celule din corp.
Toate reacțiile chimice care au loc în celulă pot fi împărțite în două grupe. Ca urmare, unele dintre aceste substanțe sau acele substanțe se descompun în molecule mai mici. Ca rezultat al altor substanțe moleculare mici, sunt sintetizate substanțe cu molecule mai mari. O moleculă din orice substanță este formată din atomi care sunt ținuți împreună prin legături chimice, adică printr-o anumită cantitate de energie chimică potențială concentrată. Când molecula este ruptă, legăturile se rup și energia lor chimică este eliberată. Pentru sinteză, adică pentru formarea unei molecule mai mari din cele mici, este necesar să se creeze noi legături chimice. Este necesar să se pună o parte din energie în ele. Dimpotrivă, reacțiile de descompunere continuă în cele din urmă cu eliberarea de energie, deoarece în timpul lor legăturile dintre atomi sunt rupte. Sensul biologic al reacțiilor de descompunere care au loc în celulă este acela că eliberează energie chimică, care este apoi utilizată de celulă pentru reacții de sinteză și pentru producerea altor tipuri de lucrări (electrice, mecanice, lucrări la transportul substanțelor). Deoarece celula a sintetizat diferite substanțe de-a lungul vieții sale, trebuie să descompună continuu alte substanțe. Energia eliberată nu este utilizată imediat pentru sinteze. În primul rând, este stocat prin formarea de substanțe speciale - acumulatori de energie chimică - acid adenozin trifosforic (ATP) și compuși înrudiți. La momentul potrivit și la punctul adecvat al celulei, ATP este divizat și renunță la energie pentru sinteza substanței necesare celulei.
Toate reacțiile chimice din celulă - atât sinteză, cât și descompunere - sunt efectuate cu ajutorul enzimelor. Enzimele sunt substanțe proteice care accelerează cursul reacțiilor. Această accelerație este atât de mare încât fără enzime astfel de reacții nu ar fi posibile deloc în celulă. Există cazuri în care, datorită enzimei, reacția este accelerată de 1011 ori. Aceasta înseamnă că o reacție care se încheie cu participarea enzimei în 0,01 sec, fără ea, ar avea loc 31 de ani. Este clar că astfel de reacții fără enzimă ar fi pur și simplu nerealiste.
În plus, datorită enzimelor, cursul reacțiilor chimice din celulă este controlat și reglat. Activitatea enzimelor dintr-o celulă se modifică în funcție de necesitatea acesteia pentru o anumită substanță sau energie, adică ca rezultat al activității acestor enzime. Atunci când este nevoie de o substanță, acele enzime, datorită cărora se formează, sunt activate sau sintetizate din nou. Formarea substanțelor, eliberarea și stocarea energiei este rezultatul reacțiilor succesive, rezultatul muncii unui întreg lanț de enzime. Toate structurile părților vii ale celulei sunt construite tocmai din enzime și din substanțe care țin aceste enzime împreună. În acest caz, enzimele care participă la reacții adiacente, care se desfășoară secvențial, sunt situate una lângă alta. Ei transferă moleculele substanțelor pe care le convertesc între ele, ca și cum ar fi de-a lungul unei benzi transportoare, și fiecare dintre ele își efectuează propria operație de lucru asupra moleculei.
Hialoplasma conține enzime care descompun moleculele de glucoză în molecule mai simple de acid piruvic. Energia eliberată în acest caz este stocată prin formarea moleculelor de ATP. Același proces are loc în nucleul celular. Cu toate acestea, cea mai mare parte a energiei este produsă în organite speciale ale citoplasmei - mitocondriile, deoarece există o descompunere mai profundă a substanțelor.
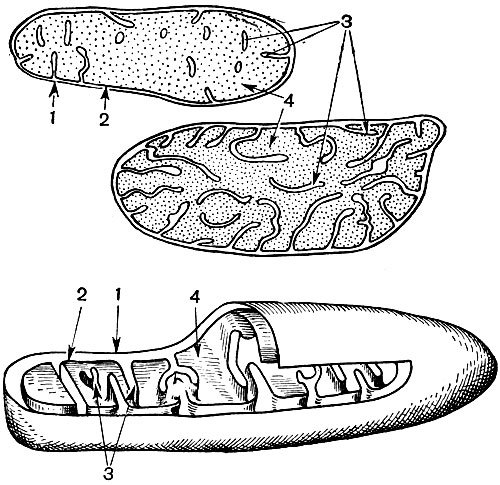
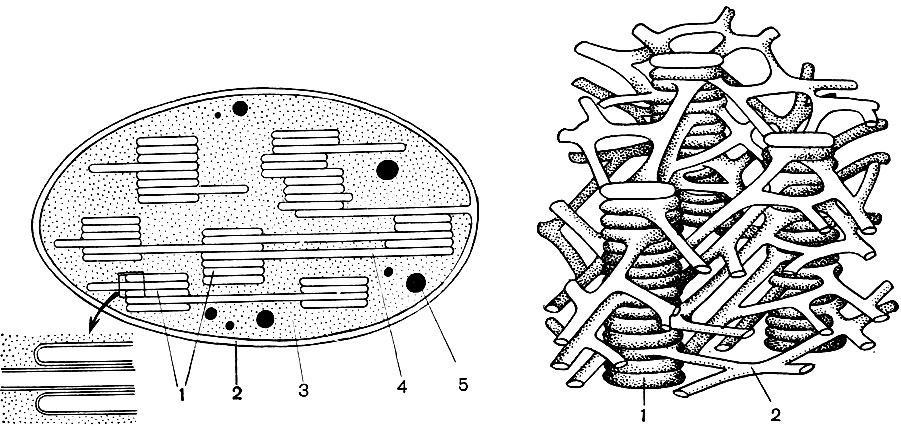
Mitocondriile sunt corpuri mici de formă rotundă sau alungită, de 0,5 - 1,5 microni, adică de mărimea unei bacterii. Numărul lor într-o celulă este de obicei mare, aproximativ 100 - 3000. Există, totuși, celule cu un număr mic de mitocondrii. Deci, sperma algelor fucus conține doar 4 mitocondrii, iar alga unicelulară micromonas conține una. Mitocondriile sunt vizibile la microscopul cu lumină, dar structura lor fină poate fi studiată doar cu microscopul electronic (Tabelul 10, diagrama structurii - Fig. 31). Mitocondriile sunt formațiuni construite din membrane lipoproteice scufundate într-o substanță de bază - o matrice. Membrana mitocondrială este formată din două membrane, între care există un decalaj.
Membrana interioară a cochiliei dă numeroase invaginații în interior, acestea sunt criste. Există o matrice între ele. Atât membrana interioară a membranei mitocondriale, cât și crestele formate de aceasta sunt construite din ordinea enzimelor localizate. Datorită pliurilor - cristae, suprafața de lucru a membranelor din interiorul mitocondriilor este foarte mare. Un număr de enzime se găsesc în matricea mitocondrială, adică între creste.
Combinația acestor enzime efectuează respirația intracelulară și stocarea energiei eliberate în timpul respirației sub formă de ATP. Activitatea mitocondriilor este strâns legată de procesele care au loc în hialoplasmă, unde au loc primele etape ale descompunerii glucozei și a altor substanțe în acid piruvic. În mitocondrii, are loc divizarea sa suplimentară. Acidul piruvic pătrunde în mitocondrii și aici, pas cu pas, este oxidat în dioxid de carbon și apă, iar oxigenul este consumat în același timp. Aceasta este respirația intracelulară, în care celula, divizând și substanțe oxidante, produce multă energie, pe care o poate folosi apoi pentru o varietate de nevoi.
Prima etapă a descompunerii moleculei de glucoză, în timpul căreia este divizată în jumătate și care are loc în hialoplasmă, conferă celulei doar două molecule ATP.
Ca urmare a celei de-a doua etape, care duce la „arderea” completă a glucozei, se formează încă 36 de molecule ATP. Prin urmare, mitocondriile prin funcția lor sunt centralele electrice ale celulei, mașini pentru obținerea volumului mare de energie. Însăși descompunerea produselor de descompunere a glucozei are loc în matricea mitocondrială, în timp ce ATP se formează datorită reacțiilor care au loc pe membranele sale interne, care includ enzime respiratorii și enzime care asigură formarea ATP. Numărul cristelor din mitocondrii poate varia. Cu cât sunt mai mulți, cu atât este mai mare activitatea biochimică a mitocondriilor.
Vorbim aici despre glucoză ca substanță, descompunând celula care produce energie. Glucoza este centrală, dar nu singura astfel de substanță. Molecula sa are un schelet de șase atomi de carbon conectați între ei. Ca urmare a unui lanț lung și complex de reacții, molecula sa este zdrobită, oxidată și, în cele din urmă, împărțită în șase molecule ale unei substanțe anorganice - dioxid de carbon (CO 2), fiecare moleculă din care conține doar un atom de carbon și este extrem de oxidată. Prin adăugarea tuturor reacțiilor succesive de oxidare a glucozei și excluderea tuturor produselor intermediare, puteți obține reacția totală a acestui proces:
Amidonul se transformă cu ușurință în glucoză, după care suferă degradarea descrisă mai sus. Proteinele și grăsimile dau diverși acizi organici, care sunt transformați în produse intermediare ale descompunerii glucozei și apoi se oxidează în același mod ca acesta din urmă și cu ajutorul acelorși enzime.
Oxidarea biologică completă a materiei organice este similară arderii sale. În ambele cazuri, rezultatul este dioxidul de carbon, apa și energia eliberată. Cu toate acestea, în timpul arderii, această energie este eliberată sub formă de căldură și complet simultan; în timpul oxidării biologice, energia legăturilor chimice este eliberată în porțiuni, iar partea sa principală este legată, trecând în energia legăturii chimice fosfat a ATP. Ca rezultat, celula primește un concentrat de energie într-o astfel de formă, care apoi la momentul potrivit și la punctul adecvat poate fi utilizat pentru a crea noi legături chimice, pentru a sintetiza substanțe noi, precum și pentru a efectua alte tipuri de lucrări - electrice, mecanice, precum și pentru transportul substanțelor de la mediu la celulă, de la celulă la mediu și de la celulă la celulă.
Cu cât activitatea vitală a celulei este mai activă, cu atât este mai mare necesitatea ei de energie și cu atât mai multă mitocondrie conține. Ele pot fi distribuite inegal în aceeași celulă: sunt mai multe în partea celulei care funcționează în prezent mai activ.
Mitocondriile sunt capabile să sintetizeze unele dintre substanțele din care sunt ele însele compuse. Datorită acestui fapt, mitocondriile se pot înmulți.
Reticulul endoplasmatic este un organoid al citoplasmei, în care sunt sintetizate foarte multe substanțe (Tabelul 10). Reticulul endoplasmatic este un sistem de canale care pătrund în citoplasmă și care se îngustează în unele zone, se extind în altele, formând fie cisterne, fie pungi plate, fie tuburi ramificate. Pereții tuturor acestor formațiuni sunt construiți din membrane care includ enzime.
Ca și în alte formațiuni membranare ale celulei, enzimele din reticul sunt dispuse în mod ordonat. În acest caz, enzimele vecine efectuează reacții succesive (operațiuni de lucru), iar grupul lor efectuează întregul lanț de reacții care conduce la crearea acestei substanțe sau a acelei substanțe.
Distingeți între reticulul endoplasmatic agranular (neted) și granular. Pe suprafața exterioară a canalelor reticulului granular, există numeroase organite mici - ribozomi, a căror funcție este sinteza moleculelor de proteine. Reticulul endoplasmatic agranular, care prevalează cantitativ asupra celui granular din celula vegetală, nu transportă ribozomi.
Reticulul endoplasmatic, pe lângă faptul că este un transportor pentru multe tipuri de conversie enzimatică a substanțelor, în principal pentru sinteza lor, este, de asemenea, un sistem de autostrăzi de-a lungul căruia substanțele se deplasează prin celulă. Reticulul începe de la membrana exterioară a membranei nucleului și, ramificându-se, se apropie de diferite organite ale citoplasmei, precum și de plasmalemă. Astfel, el conectează toate părțile celulei între ele. În plus, canalele reticulului endoplasmatic trec prin plasmodesme, conectând reticulul celulelor vecine.
Mai mult, membranele reticulului endoplasmatic dezmembrează citoplasma în numeroase compartimente, datorită cărora celula nu poate fi reprezentată ca o masă omogenă în care sunt amestecate o varietate de substanțe. Condițiile dintr-un compartiment pot fi foarte diferite de oricare altul; procesele care au loc în el pot avea loc numai în el, în timp ce unul în celălalt au loc alte procese.
În cele din urmă, membranele reticulului endoplasmatic sunt acele suprafețe de-a lungul cărora se propagă biocurenții, care sunt semnale care modifică permeabilitatea selectivă a membranelor și, prin urmare, activitatea enzimelor. Datorită acestui fapt, unele reacții chimice sunt puse în mișcare, altele sunt inhibate - metabolismul este supus reglementării și se desfășoară în mod coordonat.
Multe dintre substanțele sintetizate în celulă trebuie concentrate și eliberate din celulă fie în mediul extern, fie în vacuola intracelulară. În plus, o celulă concentrează și substanțe care intră în ea din alte celule, de exemplu, dacă le stochează în rezervă. Această lucrare este realizată de dictiozomi. De obicei, într-o celulă vegetală există mai mulți dictiozomi, iar întregul lor set se numește aparatul Golge (sau complex) și celula dată. Fiecare dictiozom este un sistem de membrane stivuite (Tabelul 10 - secțiune, Fig. 32 - diagramă volumetrică). Cavitățile dintre membranele care formează acest organoid arată ca niște fante înguste, apoi saci plate - cisterne, apoi bule. Forma lor se schimbă în timpul funcționării organoidului și, aparent, depinde de gradul de umplere a spațiilor intermembranare cu substanțele secretate și acumulate. Veziculele formate și crescute sunt separate de organoid. Aparent, multe vacuole celulare, înconjurate de membrane - tonoplast, sunt produsul activității aparatului Golgi; acestea sunt bule desprinse de el și apoi mărite.
Aparatul Golgi este dezvoltat în special în celulele excretoare (secretoare) în care sunt depuse diferite substanțe sau din care sunt excretate diferite substanțe. Sintetizează și eliberează substanțe care formează membrana celulară.
Lizozomii sunt corpuri rotunjite destul de mici (aproximativ 0,5 microni în diametru) - un alt organoid al citoplasmei. Sunt acoperite cu o membrană - o membrană lipoproteică. Conținutul lizozomilor sunt enzime care digeră proteinele, carbohidrații, acizii nucleici și lipidele. Membrana lizozomală împiedică eliberarea enzimelor din organoid în hialoplasmă, altfel aceasta din urmă ar fi digerată de aceste enzime.
Se poate crede că lizozomii sunt produse ale activității aparatului Golgi, bule desprinse de acesta, în care acest organoid a acumulat enzime digestive.
Acele părți ale celulei care mor în timpul dezvoltării sale sunt distruse de enzimele lizozomale. Într-o celulă decedată, lizozomii sunt distruși, enzimele se găsesc în citoplasmă și întreaga celulă, cu excepția membranei, suferă digestie.
Ribozomii sunt organite foarte mici, cu diametrul de aproximativ 250A. Au o formă aproape sferică. Unele dintre ele sunt atașate la suprafețele exterioare (hialoplasmatice) ale membranelor care formează canalele reticulului endoplasmatic granular; unele sunt în stare liberă în hialoplasmă. O celulă poate conține până la 5 milioane de ribozomi. Ele sunt „aparate pentru sinteza proteinelor. Prin urmare, există mai multe dintre ele în celulele care formează în mod activ proteine \u200b\u200b- în celulele în creștere, în celulele care secretă substanțe proteice. Ribozomii se găsesc și în mitocondrii și cloroplaste, unde sintetizează unele dintre proteinele din care acestea organite.
Organele numite microtubuli se găsesc în multe celule. Chiar numele lor vorbește despre forma lor - acestea sunt tuburi cu un canal în interior. Diametrul lor exterior este de aproximativ 250A. Uneori, acestea sunt tuburi duble - două simple, întinse unul lângă altul și având un perete comun care le separă cavitățile. Pereții microtubulilor sunt construiți din molecule de proteine. Se crede că microtubulii sunt asociați cu activitatea contractilă (motoră) a citoplasmei și a formațiunilor sale. Din ele, ca și din părțile de construcție, aparent sunt construite structurile contractile ale flagelului, un organoid, cu ajutorul căruia se deplasează unele alge unicelulare și coloniale, precum și celule care servesc la reproducerea multor plante inferioare. În timpul diviziunii celulare, firele axului sunt formate din microtubuli, care vor fi discutate în continuare. În perioada de diviziune, microtubulii se adună în grupuri și formează aceste filamente. La sfârșitul divizării, filamentele se dezintegrează din nou în microtubuli separați. În celule sau părți ale acestora care nu au o membrană densă, microtubulii îndeplinesc, eventual, o funcție de susținere, alcătuind scheletul interior al celulei.
Plastidele sunt organite inerente numai în celulele vegetale. De obicei, acestea sunt corpuri mari, clar vizibile la microscopul cu lumină.
Există 3 tipuri de plastide: incolore - leucoplaste, verzi - cloroplaste, colorate în alte culori - cromoplaste. Fiecare tip de plastide are propria structură și își desfășoară propriile funcții inerente. Cu toate acestea, sunt posibile tranziții de plastide de la un tip la altul. Astfel, ecologizarea tuberculilor de cartof este cauzată de rearanjarea leucoplastelor lor în cloroplaste. În cultura de rădăcină de morcov, leucoplastele trec în cromoplaste. Toate cele trei tipuri de plastide sunt formate din proplastide.
Proplastidele sunt corpuri incolore, asemănătoare cu mitocondriile, dar oarecum mai mari decât ele. Se găsesc în cantități mari în celule meristematice. Leucoplastele se găsesc în celulele părților necolorate ale plantelor (fructe, semințe, rădăcini, epidermă de frunze). Forma lor este nedeterminată. Cel mai adesea, există leucoplaste în care se depune amidon (este format din zaharuri). Există leucoplaste care stochează proteine. Cele mai puțin frecvente sunt leucoplastele umplute cu grăsime; se formează în timpul îmbătrânirii cloroplastelor. Nu există diferențe semnificative între leucoplaste și proplastide.
Cloroplastele sunt plastide ale plantelor superioare în care are loc procesul de fotosinteză, adică utilizarea energiei razelor de lumină pentru a forma substanțe organice din substanțe anorganice (dioxid de carbon și apă) cu eliberarea simultană de oxigen în atmosferă. Cloroplastele au forma unei lentile biconvexe, dimensiunea lor este de aproximativ 4 - 6 mk. Se găsesc în celulele narenchimale ale frunzelor și în alte părți verzi ale plantelor superioare. Numărul lor într-o celulă variază de la 25 la 30.
Structura cloroplastului este dată în tabelele 11 și 12 (vedere în secțiune) și în diagrama din figura 33. În exterior, cloroplastul este acoperit cu o membrană formată din două membrane lipoproteice. Sub aceasta, în substanța principală (stroma), sunt comandate numeroase formațiuni - lamele. Ele formează pungi plate care se așează una peste alta în scaune obișnuite. Acești stoici, care seamănă cu monedele stivuite într-o coloană, se numesc boabe. Lamele mai lungi trec prin ele, astfel încât toate boabele de cloroplast sunt conectate într-un singur sistem. Compoziția membranelor care formează granulele include un pigment verde - clorofilă. Aici au loc reacțiile luminoase ale fotosintezei - absorbția razelor de lumină de către clorofilă și conversia energiei luminii în energia electronilor excitați. Electronii excitați de lumină, adică având energie în exces, renunță la energie pentru a descompune apa și a sintetiza ATP. Când apa se descompune, se formează oxigen și hidrogen. Oxigenul este eliberat în atmosferă, iar hidrogenul este legat de proteina feredoxină. Ferredoxina este apoi reoxidată, eliberând acest hidrogen la un agent reducător, prescurtat ca NADP. NADP intră în forma redusă - NADP-H 2. Astfel, rezultatul reacțiilor la lumină ale fotosintezei este formarea de ATP, NADP-H 2 și oxigen, iar apa și energia luminii sunt consumate.
ATP acumulează multă energie - este apoi utilizat pentru sinteze, precum și pentru alte nevoi ale celulei. NADP-H 2 este un acumulator de hidrogen și apoi îl dă cu ușurință. Prin urmare, NADP-H 2 este un agent chimic de reducere. Un număr mare de biosinteze sunt asociate cu reducerea, iar NADPH 2 acționează ca furnizor de hidrogen în aceste reacții.
Mai mult, cu ajutorul enzimelor stromale cloroplastice, adică în afara granulelor, apar reacții întunecate: hidrogenul și energia conținută în ATP sunt utilizate pentru a reduce dioxidul de carbon atmosferic (CO2) și a-l încorpora în substanțele organice. Prima materie organică, formată ca urmare a fotosintezei, suferă un număr mare de rearanjări și dă naștere întregii varietăți de substanțe organice sintetizate în plantă și alcătuind corpul acesteia. O serie dintre aceste transformări au loc chiar acolo, în stroma cloroplastului, unde există enzime pentru formarea de zaharuri, grăsimi, precum și tot ceea ce este necesar pentru sinteza proteinelor. Zaharurile se pot muta apoi de la cloroplast la alte structuri celulare și de acolo la alte celule vegetale sau pot forma amidon, ale cărui boabe pot fi adesea văzute în cloroplaste. Grăsimile sunt, de asemenea, depozitate în cloroplaste fie sub formă de picături, fie sub formă de substanțe mai simple, precursori ai grăsimilor, și părăsesc cloroplastul.
Complicația substanțelor este asociată cu crearea de noi legături chimice și de obicei necesită energie. Sursa sa este aceeași fotosinteză. Faptul este că o porțiune semnificativă a substanțelor formate ca urmare a fotosintezei se descompune din nou în hialoplasmă și mitocondrie (în cazul arderii complete, la substanțe care servesc drept materie primă pentru fotosinteză - C0 2 și H 2 0). Ca urmare a acestui proces, care este în esență inversul fotosintezei, energia acumulată anterior în legăturile chimice ale substanțelor descompuse este eliberată și - din nou prin ajutorul ATP - este cheltuită pe formarea de noi legături chimice ale moleculelor sintetizate. Astfel, o parte semnificativă a producției de fotosinteză este necesară numai pentru a lega energia luminii și, după ce a transformat-o în chimică, o folosește pentru sinteza substanțelor complet diferite.
Și doar o parte din materia organică formată în timpul fotosintezei este utilizată ca material de construcție pentru aceste sinteze.
Producția de fotosinteză (biomasă) este colosală. Timp de un an pe glob, este vorba de aproximativ 1010 tone. Substanțele organice create de plante sunt singura sursă de viață nu numai pentru plante, ci și pentru animale, deoarece acestea din urmă procesează substanțe organice gata preparate, hrănindu-se fie direct cu plante, fie cu alte animale care, la rândul lor, se hrănesc cu plante. Astfel, fotosinteza se află în centrul întregii vieți moderne de pe Pământ. Toate transformările de substanțe și energie din plante și animale sunt rearanjări, recombinări și transferuri de materie și energie ale produselor primare ale fotosintezei. Fotosinteza este importantă pentru toate viețuitoarele și faptul că unul dintre produsele sale este oxigenul liber, provenit dintr-o moleculă de apă și eliberat în atmosferă. Se crede că tot oxigenul din atmosferă provine din fotosinteză. Este esențial pentru respirație atât pentru plante cât și pentru animale.
Cloroplastele se pot mișca în jurul celulei. În condiții de lumină slabă, acestea sunt situate sub peretele celular care este orientat spre lumină. Procedând astfel, se transformă în lumină cu suprafața lor mai mare. Dacă lumina este prea intensă, se întorc spre ea și se aliniază de-a lungul umbrelor paralele cu razele de lumină. La iluminare medie, cloroplastele ocupă o poziție intermediară între cele două extreme. În orice caz, se obține un rezultat: cloroplastele se află în cele mai favorabile condiții de iluminare pentru fotosinteză. O astfel de mișcare a cloroplastelor (fototaxis) este o manifestare a unuia dintre tipurile de iritabilitate la plante.
Cloroplastele au o anumită autonomie în sistemul celular. Ei au propriile ribozomi și un set de substanțe care determină sinteza unui număr de proteine \u200b\u200bproprii ale cloroplastului. Există, de asemenea, enzime a căror activitate duce la formarea lipidelor care alcătuiesc lamelele și a clorofilei. După cum am văzut, cloroplastul are și un sistem autonom de producere a energiei. Datorită tuturor acestor lucruri, cloroplastele sunt capabili să-și construiască în mod independent propriile structuri. Există chiar o părere că cloroplastele (cum ar fi mitocondriile) provin din unele organisme inferioare care s-au instalat într-o celulă vegetală și au intrat mai întâi în simbioză cu aceasta și apoi au devenit partea sa integrală, un organoid.
La plantele inferioare, fotosinteza este, de asemenea, efectuată de specialiști, deși nu la fel de dezvoltată ca în cloroplastul, structurile de membrană. La bacteriile fotosintetice, membranele care conțin clorofilă formează o rețea care pătrunde în corpul bacterian. În algele albastru-verzui, membranele fotosintetice sunt fuzionate în bule plate. La algele verzi și alte alge, sistemul acestor membrane este separat de restul celulei printr-o membrană de acoperire și formează un organoid special - un cromatofor. Numărul de cromatofori dintr-o celulă este mic; adesea o celulă conține un singur cromatofor. Forma lor este foarte diferită la algele din diferite specii.
În Spirogyra, cromatoforul are forma unei panglici în spirală de-a lungul pereților celulari; în Clostridium, aceștia sunt cilindri cu nervuri; zygnema are corpuri stelate.
Cromoplastele apar fie din proplastide, fie din cloroplaste, fie din leucoplaste. Structura lor membranară interioară este mult mai simplă decât cea a cloroplastelor. Gran nu este, stroma conține mult pigment galben sau portocaliu. Cromoplastele se găsesc în celulele petalelor, fructelor și culturilor radiculare.
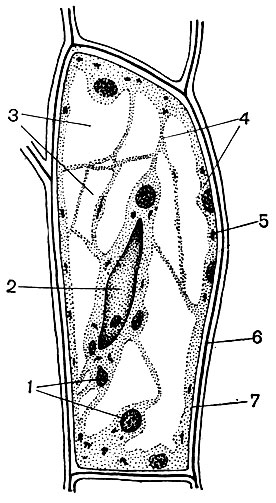
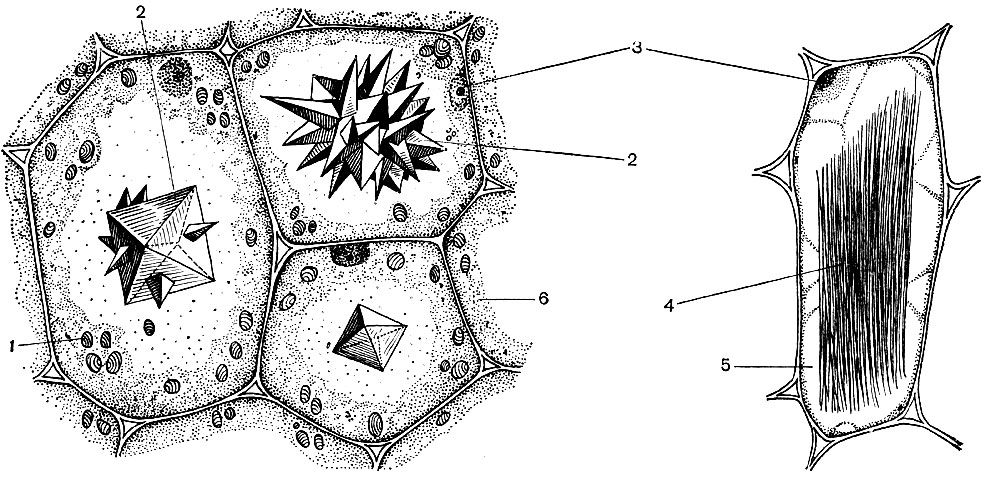
O celulă tipică de plantă are un vacuol mare umplut cu conținut lichid. Adesea vacuolul ocupă aproape întregul volum al celulei, astfel încât citoplasma este doar un strat subțire adiacent membranei celulare. Celulele tinere au mai multe vacuole mici, care, pe măsură ce celulele se dezvoltă, cresc și se îmbină într-unul singur. Conținutul vacuolului - seva celulară - este o soluție apoasă de foarte multe substanțe: zaharuri, aminoacizi, alți acizi organici, pigmenți (coloranți), vitamine, taninuri, alcaloizi, glicozide, săruri anorganice (nitrați, fosfați, cloruri) și, uneori, proteine.
Toate aceste substanțe sunt produse reziduale ale celulei. Unele dintre ele sunt depozitate în seva vacuolară (celulară) ca substanțe de rezervă și, în timp, reintră în citoplasmă pentru utilizare. Altele sunt deșeuri metabolice eliminate din citoplasmă. Deci, acidul oxalic este îndepărtat în vacuol; cristalele de oxalat de calciu sunt adesea depuse în seva vacuolară - uneori sub formă de cristale unice, în alte cazuri sub forma unui conglomerat de cristale ale acestei sări - multifacetate (drusen) sau asemănătoare acelor (raffida), prezentate în figura 34.
Marea majoritate a plantelor (cu excepția organismelor procariote) fiecare celulă vie are un nucleu sau mai mulți nuclei. O celulă lipsită de un nucleu nu poate trăi decât pentru o perioadă scurtă de timp. Celulele anucleate ale tuburilor de sită sunt celule vii. Dar nu trăiesc mult. În toate celelalte cazuri, celulele non-nucleare sunt moarte.

Nucleul se află întotdeauna în citoplasmă. Forma nucleului poate fi diferită - rotundă, ovală, puternic alungită, neregulată multilobată. În unele celule, contururile nucleului se schimbă în timpul funcționării sale și pe suprafața sa se formează lobi de diferite dimensiuni. Dimensiunile nucleelor \u200b\u200bnu sunt aceleași în celulele diferitelor plante și în celulele diferite ale aceleiași plante. Nucleii relativ mari se găsesc în celule tinere, meristematice, în care pot ocupa până la 3/4 din volumul întregii celule. Mărimile relative și uneori absolute ale nucleelor \u200b\u200bdin celulele dezvoltate sunt mult mai mici decât la cele tinere.
În exterior, miezul este acoperit cu o coajă formată din două membrane, între care există un decalaj - spațiul perinuclear. Coaja este întreruptă de pori. Exteriorul celor două membrane ale membranei dă ieșiri care trec direct în pereții reticulului endoplasmatic al citoplasmei. Atât porii, cât și conexiunea directă a reticulului endoplasmatic cu spațiul perinuclear asigură un contact strâns între nucleu și citoplasmă.
Conținutul nucleului este o substanță bazică granulară (suc nuclear sau nucleoplasmă), în care sunt plasate structuri mai dense - cromozomi și un nucleol. Nucleolul este aparatul pentru sintetizarea materialului ribozomilor și locul asamblării acestora din acest material.
Cromozomii sunt construiți dintr-un număr mare de molecule de acid dezoxiribonucleic (ADN) combinate cu molecule de proteină histonică. Moleculele de ADN sunt fire duble lungi, complexe. Fiecare moleculă este formată din două fire, răsucite spiralat una în jurul celeilalte. Un fir, la rândul său, este un lanț dintr-un număr imens de așa-numitele nucleotide. Nucleotida este un compus de bază azotată, carbohidrați (dezoxiriboză) și acid fosforic.
Fiecare dintre nucleotide conține una dintre următoarele patru baze azotate: adenină, guanină, citozină sau timină. În consecință, în ADN se disting 4 nucleotide diferite: adenină (A), guanină (G), citozină (C) și timină (T). Nucleotidele sunt legate între ele prin grupările lor fosfat, formând astfel un lanț lung. Două lanțuri, răsucite unul în jurul celuilalt și formând o moleculă de ADN, sunt legate între ele prin interacțiune chimică (așa-numitele legături de hidrogen) ale bazelor azotate ale nucleotidelor lor. Bazele formează perechi - o bază dintr-un lanț, cealaltă din al doilea. Diagrama din Figura 35 oferă o idee despre ordinea în care nucleotidele sunt legate într-un lanț și cele două lanțuri - între ele.
Deși toate moleculele de acid dezoxiribonucleic sunt construite conform planului unic descris, compoziția lor calitativă specifică este diferită; ele diferă și în ceea ce privește mărimea moleculelor. O moleculă de ADN conține până la 50-100 de mii de perechi de baze, dar numărul de perechi al acestora din fiecare moleculă este diferit de numărul de perechi de nucleotide din orice altă moleculă. În plus, există 4 tipuri diferite de nucleotide și fiecare moleculă de ADN are propriul procent inerent de nucleotide de fiecare tip dat. Cu alte cuvinte, raportul cantitativ A: G: C: T pentru fiecare moleculă de ADN este diferit. Și, în cele din urmă, în fiecare moleculă de ADN, ordinea alternanței nucleotidelor A, G, C, T este caracteristică numai pentru această moleculă. Permutarea a doar două perechi de 50 de mii de perechi de baze, chiar dacă adiacente, schimbă dramatic proprietățile întregii molecule; la fel, înlocuind cel puțin o pereche, să spunem De la - G la A - T sau G - Iz; la fel - absența unei perechi dintre aceleași 50 de mii. În realitate, moleculele ADN pot diferi unele de altele nu într-o singură legătură, nu într-una sau două perechi de nucleotide, ci într-un număr imens de ele simultan. Numărul posibilelor permutări de perechi de nucleotide într-o moleculă de ADN este infinit și, în consecință, numărul de molecule diferite este infinit, fiecare dintre ele având propriile sale proprietăți. Secvența de nucleotide dintr-o moleculă de ADN este o înregistrare criptată a compoziției unei anumite proteine \u200b\u200bcare este caracteristică unei celule date. De-a lungul fiecărei molecule de ADN, înregistrările compoziției mai multor proteine \u200b\u200bse încadrează succesiv și în toate moleculele de ADN ale tuturor cromozomilor nucleului - înregistrări ale compoziției tuturor proteinelor celulei care pot fi sintetizate în ea pe parcursul vieții sale. Esența acestor înregistrări codate este următoarea.
Moleculele fiecărei proteine \u200b\u200bsunt lanțuri de aminoacizi conectați în serie. Există aproximativ 20 de aminoacizi diferiți, iar natura unei proteine \u200b\u200beste determinată de aminoacizii din care constă molecula sa, care este cantitatea lor totală în moleculă și în ce secvență sunt conectați între ei. De exemplu, o secțiune de ADN responsabilă de compoziția unei anumite proteine \u200b\u200b(fiecare astfel de secțiune se numește genă) este o înregistrare a unei secvențe specifice de aminoacizi care formează o moleculă a unei proteine \u200b\u200bdate, numărul lor total în ea. La fiecare 3 nucleotide consecutive ale lanțului ADN desemnează (codifică) un aminoacid al moleculei proteice corespunzătoare. Următoarele 3 nucleotide codifică următorul aminoacid și așa mai departe. Numărul posibilelor combinații diferite de 3 nucleotide dintre patru tipuri diferite este mai mult decât suficient pentru a codifica 20 de aminoacizi. Este interesant faptul că aceiași aminoacizi din întreaga lume vie sunt codificați de aceleași combinații de nucleotide. Ca rezultat, într-o genă, compoziția sa de nucleotide codifică compoziția de aminoacizi a tuturor proteinelor care pot fi sintetizate într-o celulă dată.
Proteinele celulare sunt enzime. Enzimele determină cursul tuturor reacțiilor care alcătuiesc esența activității vitale a celulelor. Formarea și transformarea tuturor celorlalte substanțe ale celulei depinde de prezența anumitor enzime, fie că sunt grăsimi, carbohidrați, alcaloizi, rășini etc. Aceasta, la rândul său, formează toate proprietățile unei celule care o disting de celulele altor plante din nucleele celulare. care a codificat compoziția unui alt set de enzime. De aceea, informațiile despre compoziția tuturor proteinelor care se pot forma într-o celulă sunt informații despre toate proprietățile unei celule și ale unui organism. Mai departe vom vedea că aceste informații sunt ereditare, adică sunt transmise pe deplin de la celulă la celulă în timpul reproducerii lor și de la planta mamă la plantele fiice.
Aceste informații sunt stocate în cromozomi. Cu toate acestea, realizarea acestor informații - sinteza proteinelor - nu are loc în ele. Genele cromozomiale acționează doar ca inițiatori ai acestei sinteze. Când o celulă trebuie să formeze o anumită proteină, gena în care este criptată compoziția acestei proteine \u200b\u200beste activată. Aceasta înseamnă că moleculele așa-numitului acid ribonucleic informațional (i-ARN) sunt formate pe secțiunea catenei ADN care alcătuiește această genă.
Structura și compoziția acestor molecule, care sunt lanțuri unice de nucleotide, reflectă structura nucleotidică a genei pe care au fost formate. Astfel, informațiile despre compoziția viitoarei proteine \u200b\u200bsunt copiate.
Formarea moleculelor de ARN m (multe dintre ele se formează pe fiecare genă) înseamnă, de asemenea, multiplicarea informațiilor, așa cum ar fi, imprimarea multor matrice identice care reflectă structura aceleiași gene și, prin urmare, poartă informații despre compoziția aminoacizilor unei proteine \u200b\u200bdate în secvența nucleotidică a moleculelor lor. Aceste amprente genetice trec de la nucleu la citoplasmă. Aici, informațiile conținute în matrice - molecule de i-ARN, sunt descifrate, informațiile sunt realizate, sunt traduse din limbajul secvenței de nucleotide în limbajul secvenței de aminoacizi, adică sinteza moleculelor de proteine \u200b\u200bdintr-o compoziție dată.
În decodare și sinteză, pe lângă molecula de i-ARN, sunt implicați un număr mare de molecule ale diferiților acizi ribonucleici de transport (ARN-t), ribozomi și o serie de enzime. Aminoacizii se leagă de ARN t - moleculă cu moleculă. Fiecare dintre cele 20 de tipuri de aminoacizi are propriul său ARN-t. Deci, molecula t-ARN are grupuri chimice care îi pot recunoaște aminoacidul, alegându-l dintre toți aminoacizii disponibili. Acest lucru se întâmplă cu ajutorul unor enzime speciale. După ce și-a recunoscut aminoacidul, t-ARN intră într-o legătură cu acesta. Ribozomul este atașat la începutul moleculei de i-ARN, care, deplasându-se de-a lungul i-ARN-ului, se conectează între ele într-un lanț polipeptidic exact acei aminoacizi a căror ordine este criptată de secvența nucleotidică a i-ARN-ului dat. Această citire a informațiilor, această decodificare a acesteia are loc datorită unui mecanism special disponibil în ARN-ul de transport, ale cărui molecule aliniază exact acei aminoacizi care sunt „desemnați”, „scrise” de secvența nucleotidică a moleculei de i-ARN. Ribozomul este un aparat pentru legarea chimică a aminoacizilor într-o moleculă proteică.
Deci, schema generală este următoarea. Pe gena activată - o secțiune a unuia dintre lanțurile unei molecule de ADN - sunt sintetizate lanțuri de nucleotide, molecule de i-ARN, a căror compoziție reflectă cu exactitate compoziția genei și, prin urmare, poartă o înregistrare a compoziției proteinei codificate de această genă. În citoplasma ribozomului, pe baza informațiilor transferate de la genă de moleculele m-ARN, folosind ARN-uri de transport, diferiți aminoacizi sunt combinați într-o secvență dată, rezultând formarea unei molecule a proteinei a cărei compoziție este codificată în această genă.
Formarea moleculelor de i-ARN pe gena activată servește atât ca comandă, cât și ca program specific pentru sinteza unei proteine \u200b\u200bstrict definite. Aceeași moleculă de ARNm este utilizată ca plan pentru a crea multe dintre aceleași molecule de proteine. Cu toate acestea, este destul de scurtă, prin urmare, pentru o sinteză pe termen lung a moleculelor noi ale aceleiași proteine, este necesar să se formeze copii identice, dar noi ale moleculelor de i-ARN pe aceeași genă. Odată cu trecerea unei gene la o stare inactivă - și acest lucru se întâmplă atunci când dispare nevoia celulei pentru o anumită proteină - aceasta este blocată, nu mai formează i-ARN și în curând sinteza acestei proteine \u200b\u200bse oprește. În timpul vieții unei celule, are nevoie de diverse proteine. De fiecare dată, genele care le definesc sunt activate și se formează compoziția corespunzătoare a moleculelor de i-ARN.
Fiecare ribozom produce multe molecule de proteine \u200b\u200bdiferite pe parcursul vieții sale. Poate funcționa pe baza oricărui i-ARN, iar rezultatul, natura proteinei create de acesta, depinde doar de compoziția i-ARN-ului în contact cu care a funcționat de data aceasta ribozomul.
Astfel, nucleul celular îndeplinește următoarele funcții interdependente. Stochează informații despre compoziția tuturor proteinelor care pot fi sintetizate într-o anumită celulă pe parcursul vieții sale. (Excepția face unele proteine \u200b\u200bale mitocondriilor și ale cloroplastelor. Compoziția lor este codificată în ADN-ul propriu al acestor organite, unde se află aceste ADN-uri. Aici, la fața locului, au loc toate etapele decodării informațiilor conținute în aceste ADN-uri, inclusiv sinteza acestor proteine \u200b\u200butilizând proprii ribozomi.)
Nucleul stochează informații despre toate proprietățile celulei și ale organismului. Nucleul organizează sinteza fiecăreia dintre aceste proteine \u200b\u200bla momentul potrivit. În timpul diviziunii celulare, însoțită de diviziune nucleară, toate aceste informații sunt transferate în întregime către fiecare dintre nucleii nou formați, către fiecare celulă nouă. Acest lucru este posibil datorită faptului că, înainte de divizare, tot materialul genetic al cromozomilor se autodublează, se formează două copii identice ale acestuia și unul dintre ei ajunge în fiecare dintre noii nuclei. Nucleul fiecărei celule conține un set complet de gene caracteristice unui anumit organism. Cu toate acestea, nu toate genele funcționează în timpul vieții diferitelor celule specializate. În unele celule, o parte a genelor funcționează, în altele - alta, în a treia - o treime. De aceea, celulele diferitelor țesuturi ale unui organism diferă una de alta. O parte semnificativă a genelor rămâne într-o stare pasivă, doar de „stocare a informațiilor”, de la naștere până la moartea unei celule sau a unui organism. În plus, genele dintr-o anumită celulă nu funcționează simultan: unele sunt active într-o perioadă a vieții unei celule, într-un anumit stadiu al dezvoltării acesteia; altele - pe de altă parte etc.
De ce unele gene rămân inactive, cum activează și dezactivează altele este o problemă foarte complexă și importantă care este acum investigată intens. Proteinele-histone par să joace un rol important în blocarea și deblocarea genelor. Acestea fac parte din cromozomi, fiind împreună cu ADN-ul. Poate că activarea, „eliberarea” unei gene are loc atunci când o moleculă de histonă este detașată de secțiunea corespunzătoare a ADN-ului, expunându-i astfel lanțurile, permițându-le să se desfacă și să înceapă să funcționeze chimic. Atașarea histonei duce la blocarea genei. Cu toate acestea, modul în care este controlată atașarea și detașarea histonelor nu este suficient de clar.
După cum sa spus, materialul genetic se găsește în cromozomi. Prin urmare, munca nucleului de a stoca informații ereditare, de a le copia și de a le transfera de la celulă la celulă, de a organiza sinteza diferitelor proteine \u200b\u200bîn timpul vieții unei celule este în primul rând opera cromozomilor. Deși există pe tot parcursul vieții unei celule, ele se disting în nucleu sub formă de structuri clare numai în timpul diviziunii celulare. În acest moment, condensarea materialului cromozomilor și a cromozomului poate fi detectată prin metode de microscopie ca formațiuni individuale, clar definite.
Cromozomii au forme diferite. Acestea sunt fie bețe drepte, fie curbate, fie diferite cârlige, corpuri ovale, bile (Fig. 36). Mărimea variază foarte mult. Fiecare celulă vegetală a unei specii date conține în nucleul său același set (sau seturi) de cromozomi dintr-un număr strict definit de cromozomi diferiți, dar strict definiți. În toate plantele superioare și unele inferioare, două generații alternează în timpul ciclului lor de viață: cu celule care conțin un singur set (haploid, n) de cromozomi în nucleii lor și cu celule ale căror nuclee au un set dublu (diploid, 2 n) de cromozomi. Un set haploid este format din n cromozomi diferiți, unul din fiecare tip. Valoarea numerică este strict constantă pentru toate celulele haploide din toate plantele unei specii date. Deci, pentru căpșunile sălbatice acest număr este 7, pentru mazăre - și 7, dar pentru fasole - 11, pentru un măr - 17, pentru o pere de pământ - 51, pentru trestia de zahăr - 60. Un set diploid este două haploide împăturite împreună. Conține doi cromozomi de fiecare tip. Dacă un bob de mazăre are 7 cromozomi diferiți într-un set haploid, atunci un set diploid are 7 perechi diferite de cromozomi și doi cromozomi în fiecare pereche sunt aceiași. Este ușor de calculat că pentru plantele enumerate setul diploid (2n) pentru căpșuni este 14, pentru mazăre - 14, pentru fasole - 22, pentru un măr - 34, pentru o pere de pământ - 102, iar pentru trestia de zahăr - 120 cromozomi.
În ciclul de viață al mușchilor și al ferigilor, organismele alternează cu celule diploide și haploide. În ferigi, aceste organisme duc o viață independentă, izolată. La mușchi, o plantă diploidă trăiește pe o plantă haploidă. Într-un organism diploid, se formează celule haploide - spori. Fiecare dintre ei, germinând, dă un organism haploid, adică un organism construit din celule haploide. În ea, se formează celule sexuale haploide - gameți.
În momentul fertilizării, gametii masculi și feminini fuzionează, formând o singură celulă - un zigot, care are un nucleu - rezultatul fuziunii nucleilor ambilor gameti. De la fiecare dintre gameți, nucleul zigotului primește un set haploid de cromozomi și, ca rezultat, are un set dublu, diploid. Un organism se dezvoltă dintr-un zigot, a cărui celulă are un set diploid de cromozomi.

Într-un set diploid, doi cromozomi ai fiecărei perechi sunt identici ca formă, structură internă și conțin gene care controlează aspectul trăsăturilor omogene (Fig. 36). Se numesc cromozomi omologi. Unul dintre ei provine din setul haploid al gametului patern, celălalt din cel matern. Prin urmare, în organismele dioice, unul dintre ele poartă gene care determină dezvoltarea caracteristicilor aflate sub jurisdicția sa în funcție de tipul patern, al doilea - în funcție de tipul matern.
Cromozomii omologi ai unei alte perechi determină în același mod dezvoltarea unei alte serii de caractere, a treia pereche-a treia linie etc. Deși organismele paterne și materne aparțin aceleiași specii, proprietățile lor ereditare nu sunt identice - au și diferențe individuale. Prin urmare, unele gene ale unui cromozom omolog nu sunt identice cu genele corespunzătoare ale celui de-al doilea.
Setul haploid, care face parte din diploid și provine din gametul patern, poartă ereditatea paternă cu trăsăturile sale individuale, iar setul haploid din gametul matern îl poartă pe cel matern. Interacțiunea complexă a genelor omogene, dar nu întotdeauna identice a două seturi haploide, care formează împreună un diploid, determină ce trăsături vor apărea în descendenții diploizi, care, în esență, este un hibrid de tată și mamă.
La gimnosperme și angiosperme, alternanța generațiilor se produce, în principiu, ca la mușchi și ferigi, dar faza lor haploidă este mult redusă și este adesea reprezentată doar de un grup de celule. Ea nu trăiește independent, ci în corpul unei plante haploide. În angiosperme, generația haploidă feminină este închisă în sacul embrionar situat în ovul, iar generația masculină este în interiorul bobului de polen.
Chiar înainte de divizarea celulară, fiecare moleculă de ADN din fiecare cromozom se atașează singură o copie a sa - o secundă a aceleiași molecule. Ca rezultat, întregul material ereditar al celulei se dublează, iar fiecare cromozom constă acum din două părți egale - cromatide. Mai mult, „sarcina” celulei este de a împărți fiecare cromozom în cromatide, distribuindu-le strict în mod egal între viitoarele celule fiice: o cromatidă din fiecare cromozom trebuie trimisă fiecăreia dintre ele. Acest lucru se face după cum urmează. Imediat înaintea diviziunii celulare, cromozomii sunt puternic condensați și contractați. Apoi, acestea sunt situate într-un plan - de-a lungul ecuatorului nucleului, cu o cromatidă din fiecare dintre ele orientată către un pol al celulei, celălalt - către opus. Învelișul nuclear dispare, iar nucleolul se dizolvă și el. Filamentele apar între polii celulei, care împreună formează o formă de fus. Filamentele sunt colectate din microtubuli care se conectează între ei. Fusul este format din două tipuri de fire. Unele sunt celule continue care merg de la un pol la altul. Alții trag, fiecare dintre care conectează un pol la una dintre cromatide. În cromozom există un site - cinetocorul, de care sunt atașate firele de tragere - unul de la un pol, celălalt de la celălalt. Mai mult, cromatidele diverg. Cromatidele, care anterior constituiau fiecare cromozom, sunt separate unele de altele și, trase în sus de filamente, diverg spre polii opuși ai celulei. Ca rezultat, un set complet de cromatide este colectat la fiecare pol, care sunt acum cromozomi. După aceasta, firele fusului se dezintegrează, se formează un înveliș nuclear în jurul fiecărui set de cromozomi, cromozomii se umflă foarte mult (despiralizează). În fiecare nucleu apare un nucleol. Nucleul capătă structura caracteristică nucleelor \u200b\u200bcelulelor care nu se divid. În planul median al celulei se formează un sept, împărțind celula în două celule fiice.
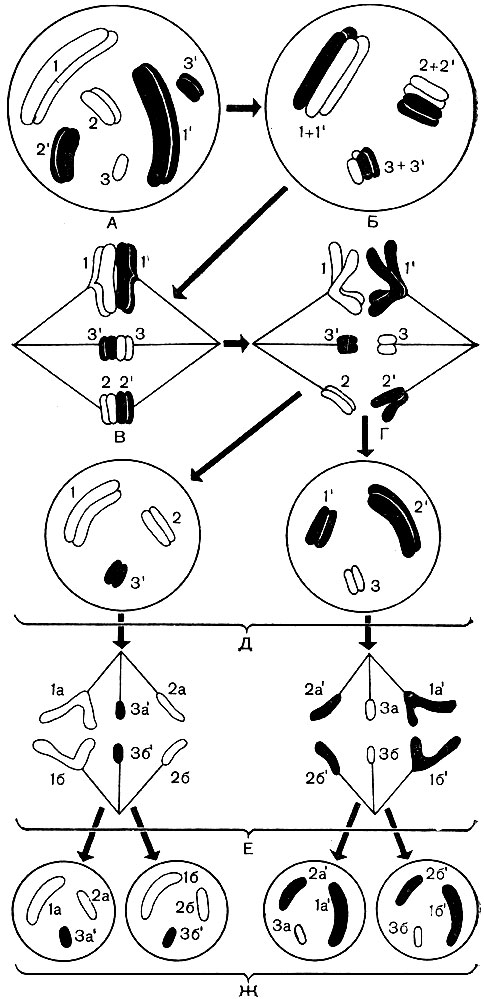
Figura: 38. Schema comportamentului cromozomilor celulei „condiționate” în timpul meiozei. A - descrie un nucleu celular diploid care conține 3 perechi de cromozomi (1 și 1 "- prima pereche de cromozomi omologi, 2 și 2" - a doua pereche, 3 și 3 "- a 3-a pereche). 2n \u003d 6,3 cromozomii originari din organismul matern sunt ușori (numere fără accident vascular cerebral); cromozomii lor omologi asociați originari din organismul patern sunt înnegriți (numere cu accident vascular cerebral). Cromozomii omologi sunt desemnați printr-un număr comun. Fiecare cromozom este format din două cromatide. B - fuziunea cromozomilor omologi În acest stadiu, cromozomii omologi schimbă secțiuni separate între ele. Ca rezultat, apare o anumită redistribuire a materialului ereditar matern și patern între cromozomi (încrucișare). C - se formează filamente fus, care se atașează la cromozomi, membrana nucleară dispare. D - cromozomii omologi diferă de poli opuși. celule, la poli există câte un cromozom omolog din fiecare pereche, numărul total de cromozomi la fiecare pol este jumătate din cel din nucleul original A. D - imagine există două nuclee cu un set haploid de cromozomi în fiecare; unul dintre ei a primit mai mulți cromozomi ai bunicii (ușori, materni în raport cu nucleul A) și mai puțin cromozomii bunicului (înnegriți, paterni în raport cu nucleul A), în celălalt - dimpotrivă; astfel, noii nuclei nu sunt complet identici între ei în compoziția substanței lor ereditare; diferența lor se datorează și încrucișării care are loc cu cromozomii în stadiul B. E - G - diviziune mitotică a fiecăruia dintre nucleii haploizi D: divizarea longitudinală a fiecărui cromozom, divergența cromozomilor la poli, formarea a doi nuclei haploizi din fiecare nucleu D. Ca urmare, au apărut 4 celule. cu nuclei haploizi în loc de o celulă cu un set diploid de cromozomi
Organele nu sunt distribuite strict în mod egal între celulele fiice, dar apoi părțile lor constitutive sunt sintetizate în celule, sunt asamblate noi copii ale organelor din fiecare specie, iar numărul lor în fiecare celulă este restabilit la normal. Celulele în sine cresc. În cromozomi, materialul ereditar este duplicat, după care cromozomii constau, ca înainte de diviziunea celulară, din două cromatide. Celula este pregătită pentru o nouă diviziune.
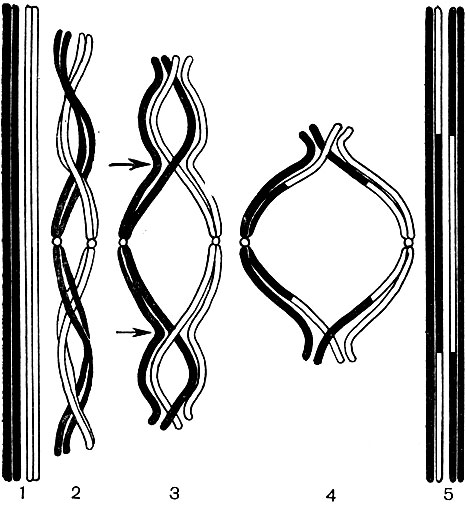
Figura: 38A. Schema de încrucișare în timpul meiozei în stadiul B din Fig. 38. 1 - doi cromozomi omologi, fiecare dintre ei constând din două cromatide surori; 2 - cromozomii omologi se împletesc între ei; 3 - fiecare cromozom este divizat în cromatidele sale constitutive; cromozomii încep să se respingă și să divergă, dar în unele puncte (în figură - în patru) rămân conectați; cromatidele surori sunt dublate în bucla centrală, iar cromatidele surori în ambele bucle extreme; punctele în care este posibilă trecerea sunt afișate cu săgeți; 4 - cromatidele au schimbat zone, iar cromozomii, care au suferit un fel de hibridizare reciprocă, diverg; 5 - doi divergenți, dar deja „hibrizi” (spre deosebire de cei arătați pe 1) cromozomi
Când generația diploidă de celule este înlocuită de una haploidă, apare așa-numita diviziune nucleară de reducere, meioza. În timpul meiozei (Fig. 38), cromozomii omologi ai fiecărei perechi se apropie unul de altul, se învecinează strâns de-a lungul lungimii lor și se răsucesc. Secțiuni separate sunt schimbate între cromozomii omologi adiacenți. Ca rezultat, unele dintre genele cromozomilor paterni sunt transferate în compoziția cromozomilor materni, iar genele corespunzătoare ale cromozomilor materni ocupă locurile libere din cromozomii paterni (trecerea peste fenomen - Fig. 38A). Aspect acești și alți cromozomi nu se modifică ca rezultat, dar compoziția lor calitativă devine diferită. Moștenirea paternă și maternă este redistribuită și mixtă. Mai mult, învelișul nuclear și nucleolul se dizolvă, se formează un aparat ax, la fel ca în timpul mitozei. Cromozomii omologi sunt separați și, cu ajutorul firelor axului, diverg spre polii celulei. Un pol are un set haploid de cromozomi (un cromozom omolog din fiecare pereche), celălalt are un al doilea set haploid.
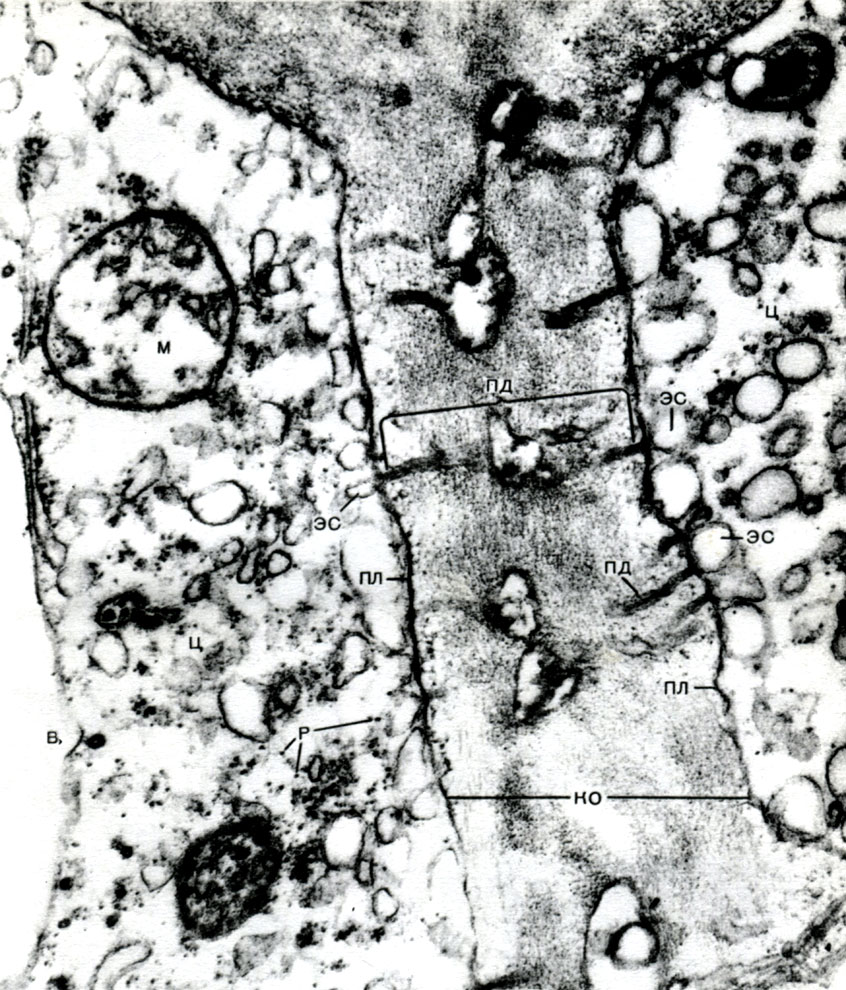
În urma meiozei, nucleii haploizi formați se divid prin mitoză. În acest caz, fiecare cromozom al setului haploid este împărțit în două cromatide, acestea divergând și se formează celule haploide fiice.
Meioza diferă de mitoză în două moduri fundamentale. În primul rând, în timpul meiozei, fiecare pereche de cromozomi omologi se adună și o parte a materialului ereditar este schimbată între ei, ceea ce nu este cazul în timpul mitozei. În al doilea rând, în timpul meiozei, cromozomii omologi diverg la polii celulei, câte unul din fiecare pereche, iar în timpul mitozei, fiecare cromozom este împărțit în cromatide, iar cromatidele diverg la poli, câte unul din fiecare cromozom. Ca urmare a meiozei, celulele haploide sunt formate dintr-o celulă diploidă.
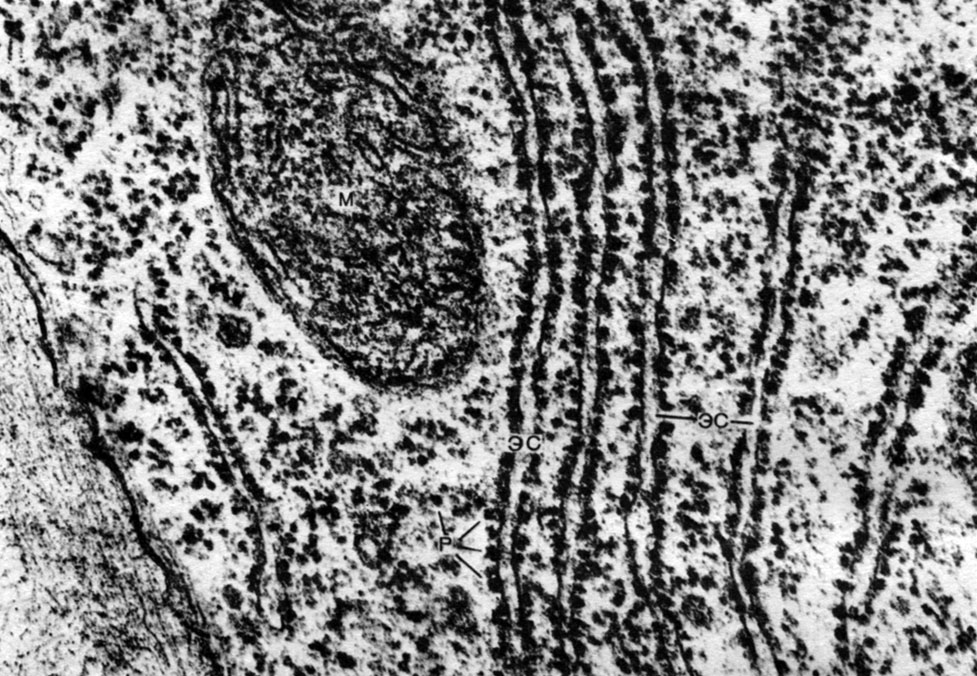
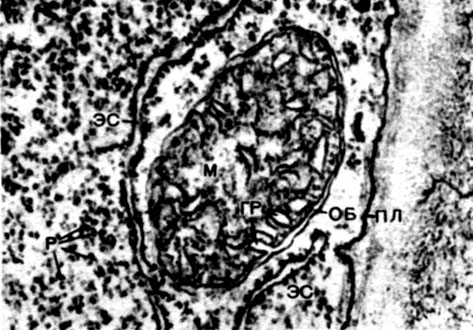
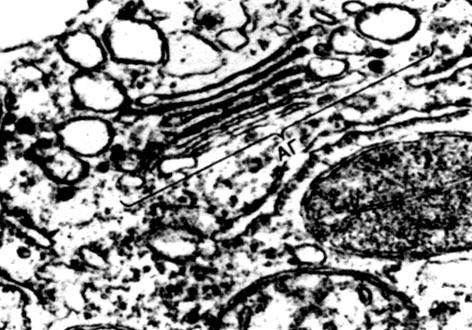
Tabelul 10. Structura celulelor vegetale. Aparatul Golgi în citoplasma părului rădăcinii în curs de dezvoltare al unei ridichi (Raphanus sativus). Micrografie electronică (mărire x 52000) de M.F. Danilova: ag - aparatul Golgi, secțiunile sacilor plati și a bulelor
În celulele unor țesuturi, în timpul dezvoltării lor, apare mitoza incompletă: materialul cromozomic din nuclee se dublează, cromozomii se împart în jumătate, dar în loc să formeze doi nuclei, rămân în nucleul original. Din acel moment, conține nu un diploid, ci un set tetraploid (cvadruplu) de cromozomi. Procesul care duce la această duplicare a cromozomilor într-un nucleu se numește endomitoză - mitoză internă. Dacă se întâmplă de două ori cu un nucleu, atunci devine un set octoploid (de opt ori) etc. Celulele ale căror nuclee poartă mai mult de două seturi de cromozomi sunt numite poliploide, adică multiploide. Poliploidia celulelor în unele cazuri crește viabilitatea lor, deoarece fiecare genă este duplicată de câteva alte gene ale aceleiași. Genele de o singură calitate acționează la unison, iar deteriorarea uneia dintre ele nu duce la pierderea trăsăturii pe care o determină, deoarece este compensată de munca genelor omogene rămase. În multe cazuri, celulele poliploide sunt mai mari și mai bogate în conținut decât celulele diploide. S-au dezvoltat soiurile de plante cultivate poliploide, care au calități economice crescute.
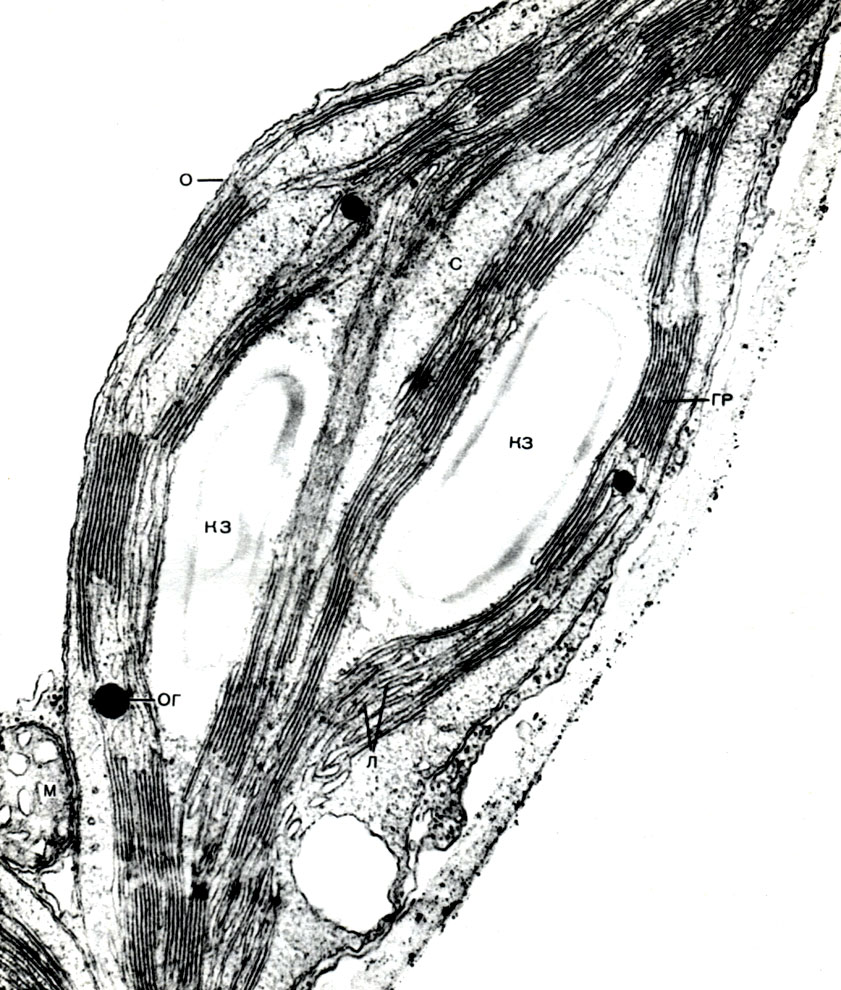
Știința care studiază celula se numește citologie. Celula este baza structurii tuturor ființelor vii și a tuturor proceselor vieții; prin urmare, majoritatea celor mai importante probleme biologice generale pot fi rezolvate cu implicarea citologiei și a metodelor de cercetare a acesteia.
Cu ajutorul citologiei, se rezolvă multe probleme practic importante: lupta împotriva bolilor plantelor (fungice, bacteriene), creșterea de noi soiuri de plante cultivate, depășirea sterilității (sterilității) soiurilor hibride.
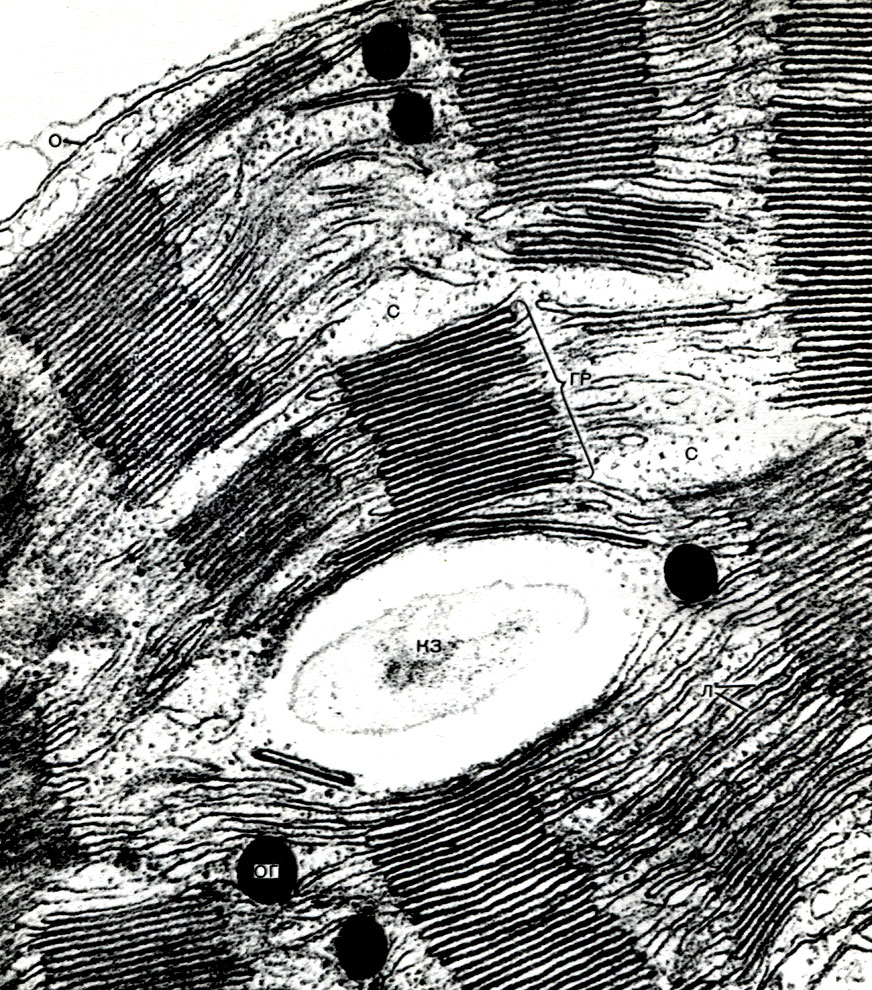
Citologia modernă investighează aceste probleme folosind o varietate de metode - microscopice, biochimice, biofizice, genetice - și interacționând îndeaproape cu alte științe biologice conexe.