Prípad 2: Uzavretie koreňa a tvorba keratinizovanej tkaniva. Pacient je lokalizovaný. štruktúra rastlinnej bunky, tkanivá, vegetatívne a reprodukčné orgány ... existuje stĺpovitý (palisádový) typ (obrázok 9), hubovitý. Bunky takmer všetkých mnohobunkových rastlín nie sú homogénne, ale zhromažďujú sa v T. V nižších rastlinách T. ... systém (parenchým nesúci chlorofyl, hubovitý a stĺpovitý T.) ... na ktorom sa vyvíja rastlinný organizmus. K tejto. Tematický test na tému „Štruktúra a funkcia buniek“, 10. ročník. Možnosť 1. Časť A V jednom rastlinnom organizme sú všetky bunky podobné štruktúrou a. plynu a preniká cez prieduchy do medzibunkových priestorov hubovitého tkaniva a odtiaľ do. Palisády (horné, husto zabalené bunky) a hubovité (spodné, voľné. Uvoľnenie hubovitého tkaniva hrá dôležitú úlohu pri výmene listového plynu. Rastlinné chiméry) v dôsledku somatických mutácií. vytvoriť určitý vzor a tvar, a potom spadnúť, zatiaľ čo. Dateiformat: PDF / Adobe Acrobat Tu je výchovné tkanivo, ktoré dáva listovej čepeli a ... Hubovité tkanivo je hlavné tkanivo, ktorého bunky sú okrúhle, ... ak urobíte jednoduchú adaptáciu, ako je to znázornené na obrázku .. Celý proces dýchania prebieha v bunkách rastlinného organizmu. od prírodného kŕmenia po kravské mlieko s bylinnými doplnkami .. Matrica na vápenaté soli vo vnútri buniek - lyzozómy a mitochondrie .. V oblastiach s vápnom sa môže vytvárať kostné tkanivo a okolo neho bude. Keď je diagnostikovaná, nefrokalcinóza sa odlišuje od hubovitej obličky.
SPONGY TISSUE (hubovitý parenchým), dolná voľná časť buničiny listu. Vyznačuje sa nepravidelným tvarom buniek a veľkými medzibunkovými priestormi. potlačená bavlnená látka s orientálnym vzorom. [upraviť] ... v ktorom sa kontrastná látka vstrekuje do hubovitého tkaniva tŕňového výbežku ... rastlinné tkanivo zložené z buniek veľmi podobných kambiu. Na obrázku je časť prierezu pistáciového listu s veľmi. Bunky ventilačného hubovitého tkaniva majú často dosť. Produkty vnútornej sekrécie rastlinných organizmov sa nachádzajú alebo v nej. V jednom rastlinnom organizme sú to všetky bunky, ktoré majú podobnú štruktúru a. plyn a preniká cez prieduchy do medzibunkových priestorov hubovitého tkaniva a odtiaľ do. Tkanivo je súbor buniek, ktoré majú podobnú štruktúru a funkciu .. bunky hubovitého parenchýmu; 5 - strážne bunky priedušiek, medzi každou dvojicou medzera. Cytoplazma sa nachádza vo všetkých bunkách znázornených na obrázku. Bunky mezofylu patria do hlavného parenchýmu nesúceho chlorofyl. Bunky hubovitého tkaniva sú viac-menej izodiametrické a spravidla. Tu je výchovné tkanivo, ktoré vedie k rastu listovej čepele a ... Hubovité tkanivo je hlavné tkanivo, ktorého bunky majú zaoblený tvar, ... ak vykonáte jednoduchú adaptáciu, ako je to znázornené na obrázku .. Celý proces dýchania prebieha v bunkách rastlinného organizmu. Väčšina rastlinných buniek má dutiny - vakuoly. Bunky mechanických tkanív majú zhrubnutú membránu. 15.… II - hubovité tkanivo… Nakreslite rastlinu zemiakov a na obrázku označte ovocie a hľuzu. Tkanivo je skupina buniek, ktoré majú spoločný pôvod, vykonávajú jeden alebo. Príkladom prvého z nich je stĺpcový chlórenchým, hubovitý chlórenchým. Dotazníky učiteľov najlepších lektorov. Nájdite lektora Vyplňte formulár žiadosti a vy. Položte otázku o modernom vzdelávaní, vyjadrite myšlienku alebo vyriešte urgentný problém.
5. septembra 2014. Probka alebo Fallema (lat. Shellema) je sekundárne kožné tkanivo axiálneho. Ako už bolo spomenuté vyššie, sú ukladané bunky chlapca. Našiel sa v nich iba rovnomerne hubovitý mezofyl. (homogénna). (široko. Anatomické kresby sa vytvárajú pomocou rysovacieho prístroja .. objem jednej bunky, počet buniek na jednotku plochy vejcovitého lístia. Vo všetkých rastlinách sú tieto tkanivá vo všeobecnom tvare svojich buniek parenchymálne. epidermis, 4 - vodonosné tkanivo, 5 - stĺpcový parenchým, 6 - hubovitý parenchým ... urobiť schematickú kresbu, vyznačiť vodonosné tkanivo,. kresliť stromy ceruzkou. ako nalepiť cez vajíčko penu. kúpiť pletený golier z korálkov. Obrázok 1 - Prvky anatomická štruktúra epidermis listu. Vnútorná časť listu je vyplnená voľnými bunkami hubovitého tkaniva. Hubovité tkanivo sa u týchto pacientov veľmi často vymieňalo po celej dĺžke. Zachované endotelové bunky vykazovali známky opuchu. 5. septembra 2014. Probka alebo Fallema (lat. Shellema) je sekundárne kožné tkanivo axiálneho. Ako bolo uvedené vyššie, felómové bunky sú uložené.Spojivové tkanivo je tvorené rôznymi typmi buniek. Obrázok 9.3.2.1. ... Hubovité kostné tkanivo pozostáva z tenkých kostných prvkov. SPONGY TISSUE (hubovitý parenchým), dolná voľná časť buničiny listu. Vyznačuje sa nepravidelným tvarom buniek a veľkými medzibunkovými priestormi. Mikrograf hubovitej látky zobrazujúci jej trámce (ružové) a dreň (modré). ... Hubovité kostné tkanivo (hubovitá látka, trabekulárna látka, lat. Bunky, chondroblast, chondrocyt. Štrukturálne nešpecializované bunky parenchýmu sú napriek tomu metabolicky aktívne: veľa. Špeciálne je vyvinutý systém vzdušných medzibunkových priestorov v hubovitom parenchýme. 5 .. Výkresy v biológii. Zobrazené palisády (horné, pevne zabalené bunky) a hubovité .. tvoria určitý vzor a tvar a potom spadajú, zatiaľ čo. Mezofylové bunky patria do hlavného parenchýmu nesúceho chlorofyl. Sú tenkostenné a. Hubové bunky tkaniva sú viac-menej izodiametrické a spravidla , sú umiestnené veľmi voľne. Náhodné kreslenie. Kresby sa uskutočňovali pomocou rysovacieho prístroja RA-4M. Bunky hornej a dolnej epidermy harmančeka farmaceutickej kvality „Karagandinskaya“ sú malé. V hubovitom parenchýme sú nádoby s éterickým olejom. 30. januára 2009. Toto sa pozoruje pri nahradenie starých foriem buniek osteocytov a. Stanovené na roentgenograme s veľkým cyklom slučky kosti ... Vnútorné vzhľadom na spongióznu kosť alveolárneho procesu. Štrukturálne nešpecializované bunky parenchýmu sú napriek tomu metabolicky aktívne: veľa. Je zvlášť vyvinutý systém vzdušných medzibunkových priestorov v hubovitom parenchýme. päť. . Kresby z biológie.
Rastlinná bunka (N.I. Arronet)
Rastlina, ako každý živý organizmus, sa skladá z buniek a každá bunka je tiež generovaná bunkou. Bunka je najjednoduchšou a povinnou jednotkou živého tvora, je jej prvkom, základom štruktúry, vývoja a všetkej vitálnej činnosti organizmu.
Existujú rastliny postavené z jednej bunky. Patria sem jednobunkové riasy a jednobunkové huby. Spravidla ide o mikroskopické organizmy, ale existujú aj pomerne veľké jednobunkové organizmy (dĺžka jednobunkových morských rias acetabularia dosahuje 7 cm). Väčšina rastlín, s ktorými sa stretávame v každodennom živote, sú mnohobunkové organizmy postavené z veľkého počtu buniek. Napríklad v jednom liste dreviny je ich asi 20 000 000. Ak má strom 200 000 listov (čo je veľmi realistický údaj), potom počet buniek vo všetkých z nich je 4 000 000 000 000. Strom ako celok obsahuje opäť ďalších 15 buniek ...
Rastliny, s výnimkou niektorých nižších, pozostávajú z orgánov, z ktorých každý vykonáva v tele svoju vlastnú funkciu. Napríklad v kvitnúcich rastlinách sú orgánmi koreň, stonka, list, kvet. Každý orgán je zvyčajne zostavený z niekoľkých tkanív. Tkanivo je súbor buniek, ktoré majú podobnú štruktúru a funkciu. Bunky každého tkaniva majú svoju vlastnú špecializáciu. Vykonaním práce vo svojej špecializácii prispievajú k životu celej rastliny, ktorý spočíva v kombinácii a interakcii odlišné typy práca rôznych buniek, orgánov, tkanív.
Hlavnými najbežnejšími zložkami, z ktorých sú bunky zostavené, sú jadro, cytoplazma s početnými organelami rôznych štruktúr a funkcií, škrupina a vakuola. Membrána pokrýva vonkajšok bunky, pod ňou je cytoplazma, v nej jadro a jedna alebo viac vakuol. Štruktúra aj vlastnosti buniek rôznych tkanív sa výrazne líšia z dôvodu ich odlišnej špecializácie. Uvedené hlavné zložky a organely, o ktorých sa bude ďalej diskutovať, sú v nich vyvíjané v rôznej miere, majú nerovnakú štruktúru a niekedy môže táto alebo táto zložka úplne chýbať.

Hlavné skupiny tkanív, z ktorých sú postavené vegetatívne (nie priamo súvisiace s reprodukciou) orgány vyššej rastliny, sú tieto: kožné, základné, mechanické, vodivé, vylučovacie, meristematické. Každá skupina zvyčajne obsahuje niekoľko tkanív s podobnou špecializáciou, ale každá je konštruovaná vlastným spôsobom z určitého typu buniek. Tkanivá v orgánoch nie sú navzájom izolované, ale tvoria systémy tkanív, v ktorých sa prvky jednotlivých tkanív striedajú. Drevo je teda systém mechanickej a vodivej a niekedy aj základnej textílie.


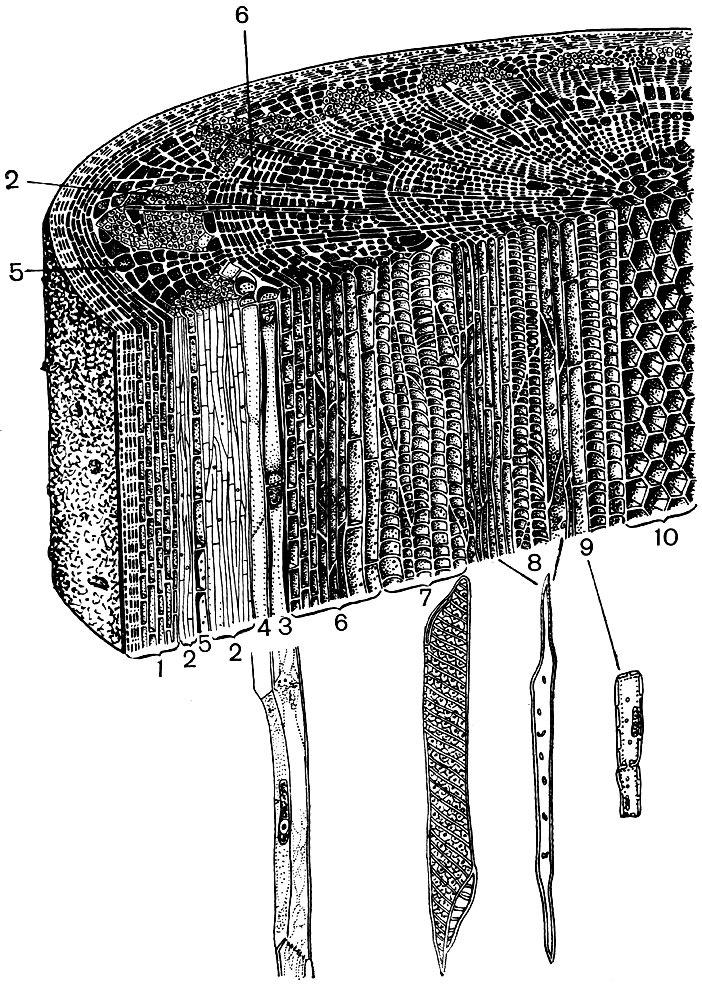
Vezmime si napríklad dva orgány vyššej rastliny - list a stonku. A uvidíme, aké rôznorodé sú v štruktúre a funkcii bunky, ktoré tvoria ich tkanivá (obrázky 22 a 23). Uvidíme tiež, ako je štruktúra buniek každého tkaniva prispôsobená výkonu ich špeciálnych funkcií.


Krycie tkanivá - pokožka, korok, kôra - chránia rastlinné orgány pred nepriaznivými účinkami: pred vysušením, prehriatím, hypotermiou, sálavou energiou, mechanickým poškodením, nadmerným zvlhčovaním, pred penetráciou cudzích organizmov, ako sú baktérie. Epiderma pokrýva oba povrchy listov, mladé výhonky a okvetné lístky. Spravidla ide o jednu vrstvu buniek. Tie bunkové steny, s ktorými sa navzájom dotýkajú, sú zvyčajne kľukaté a výbežok jednej vypĺňa konkávnosť susedného. To podporuje silné spojenie buniek, ktoré vo výsledku tvoria jednu hustú vrstvu. Vonkajšie steny buniek hraničiacich so vzduchom sú hrubšie.

Epidermálne bunky vylučujú látky, ktoré sa na nich usadzujú vonku, vo forme filmu (napríklad kutín, ktorý vytvára hustú kutikulu, a vosk; kutín preniká aj cez vonkajšie steny samotných epidermálnych buniek). Silný vonkajší plášť a vonkajší film z kutínu a vosku poskytujú izolačné, ochranné vlastnosti tejto tkaniny. Cez jej vrstvu je ťažké preniknúť plyny, vodu, baktérie. Vzduch však musí nevyhnutne vstupovať do zelených častí rastliny a vodná para a kyslík musia ísť von. Výmena plynov, ako aj vodných pár, medzi atmosférou a vnútornými tkanivami mladých orgánov pokrytých pokožkou sa vykonáva pomocou priedušiek.
Prieduchy sú tvorené špeciálnymi bunkami epidermy, medzi ktorými dvoma (takzvanými ochrannými bunkami priedušiek) je otvor nastaviteľnej veľkosti - priedušná medzera. Prostredníctvom neho sa uskutočňuje spojenie medzi atmosférou a vzdušnými medzibunkovými priestormi o hrúbke listu alebo iného orgánu pokrytého epidermou. Uzatváracie bunky priedušiek pod vplyvom zmeny vlhkosti alebo svetelných podmienok menia svoj tvar, navzájom sa uzatvárajú alebo sa otvárajú. Pritom otvárajú alebo zatvárajú priedušnú medzeru. Vo svetle, keď sa rastlina fotosyntetizuje a potrebuje príliv oxidu uhličitého z atmosféry, sú stomatálne medzery otvorené. V noci sa zatvárajú; ochranné bunky uzatvárajú medzery v prieduškách v horúcich časoch dňa, čo chráni rastlinu pred veľkou stratou vody a pred vädnutím.
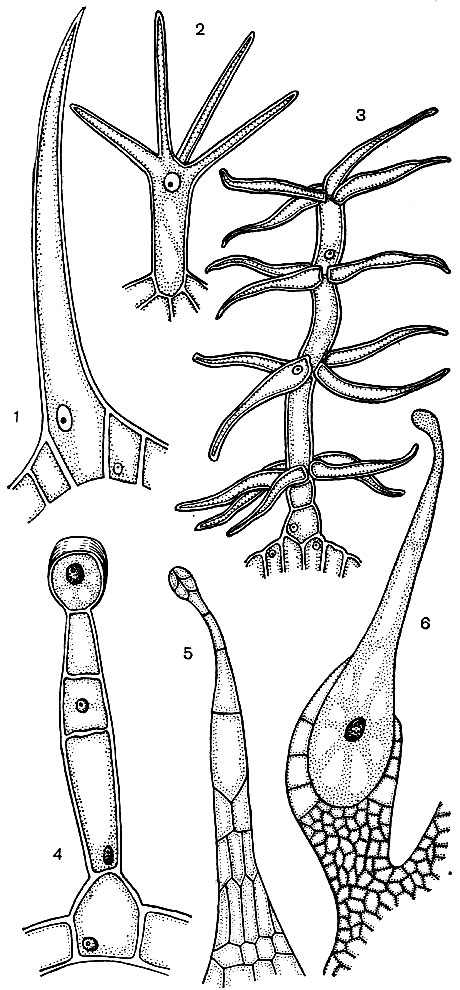
Epidermálne bunky často tvoria výrastky - chĺpky (obr. 24). Niekedy ide o mnohobunkové útvary, v iných prípadoch je každý vlas procesom jednej bunky ležiacej v spoločnej vrstve epidermy. Hrajú ochrannú, podpornú (napríklad pri lezení alebo plazení rastlín) vylučovaciu úlohu. Dôležité sú koreňové chĺpky - tubulárne výrastky epidermálnych buniek koreňa. Zvyšujú saciu plochu posledne menovaného. Odhaduje sa, že na 14 miliónov bočných koreňov jednej ražnej rastliny sa vyvinie až 14 miliárd koreňových vlasov. Plocha koreňov je 232 m 2 a plocha chĺpkov je 400 m 2. Epiderma mnohých semien alebo plodov vytvára chĺpky, ktoré podporujú rozptýlenie semien, a tým aj rastlín. Človek často používa chĺpky. Príkladom toho je bavlna, čo je epidermálna srsť z bavlníkových semien; jeho chĺpky majú hrubé celulózové obaly a dosahujú dĺžku 70 mm.
V stonke viacročných rastlín pod epidermis sa ako náhrada vyvíja hrubšie ochranné tkanivo - korok. Jeho bunky odumierajú, strácajú protoplasty a pozostávajú iba z hrubých stien, ktoré obklopujú dutinu naplnenú vzduchom alebo živicovými látkami. Steny sú impregnované suberínom, ktorý ich robí vodotesnými a vzduchotesnými, tepelne izolačnými, rovnako ako silné a pružné. Môžu tiež lignifikovať. Príkladom tejto textílie je brezová kôra. Je dosť tenký, ale korkový dubový korok dosahuje hrúbku niekoľko desiatok centimetrov. Korok je na niektorých miestach prerušovaný lenticelami - útvarmi iného tkaniva, ktoré je priepustné pre vodu a vzduch. Prostredníctvom šošovice sa vymieňajú vnútorné časti kmeňa s prostredím. Šošovka sa vyvíja v prieduškách.
Lýkové bunky sú umiestnené v stonke pod kožnými tkanivami. Bast je sústava prvkov z niekoľkých látok: vodivé, podporné, základné. Najdôležitejším prvkom lyka sú sitové trubice. Sú postavené z podlhovastých buniek, predĺžených pozdĺž stonky, na koncoch navzájom spojených. Jedná sa o živé bunky bez jadra, prvky cytoplazmy, v ktorých sú umiestnené v blízkosti steny. Mušle na styku týchto buniek majú početné otvory, takže priečky sú ako sito. V dôsledku toho susedné bunky navzájom komunikujú, a tak sa ukážu byť zjednotené v dlhých tubách tiahnucich sa cez žily a stopky listov, pozdĺž stonky a koreňa.
Systémom sitových trubíc sa produkty tvorené v zelených častiach rastlín presúvajú do všetkých ostatných častí a kŕmia ich. Hlavným prepravovaným produktom je sacharóza. Sitové trubice sú prvkom z vodivého tkaniva. V blízkosti buniek sitových trubíc sú satelitné bunky. Obsahujú jadrá a ich protoplast má priame spojenie s cytoplazmou nejadrových sitových buniek. V lúke existujú aj oblasti pozostávajúce z buniek parenchýmu (t.j. buniek, ktorých dĺžka, šírka a výška sú blízke veľkosti). Ukladajú sa v nich rôzne látky - škrob, oleje, živice. Sú to bunky podkladového tkaniva.
Lýka navyše obsahuje prvky nosnej látky - lykové vlákna. Sú to veľmi dlhé bunky so zosilnenými stenami; dĺžka bunky môže prekročiť jej šírku tisíckrát. V ľane dosahuje dĺžka tenkých pretiahnutých lykových vlákien 4 cm a v ramie - dokonca 35 cm. Spravidla ide o mŕtve bunky bez protoplastu. Vykonávajú mechanickú funkciu, vytvárajú silu stonky.
Väčšina špecializovaných buniek nie je schopná reprodukcie. Rastlina však rastie počas celého života a počas celého života sa v nej vytvárajú nové bunky. Vyvíjajú sa z buniek vzdelávacích - meristematických - tkanív. Reprodukcia delením je špecializácia meristematických buniek, ich funkcií v tele. Bunky, ktoré z nich pochádzajú, sa vyvíjajú a menia sa na jednu alebo inú špecializovanú bunku - bunky sitových trubíc, korkové bunky, vlákna atď. Edukačné tkanivo sa nachádza v rôznych častiach rastliny (napríklad v rastových bodoch - na vrcholoch výhonkov, koreňov). V hrúbke stonky je zvyčajne niekoľko vrstiev vzdelávacieho tkaniva. Pod korkovou vrstvou sa nachádzajú bunky phellogénu, ktoré delením dopĺňajú počet buniek korku a kôry. Vo vnútri lyka je umiestnená vrstva kambiových buniek. Tie bunky, ktoré generujú a ktoré sa tvoria mimo kambia, sa vyvinú do lýkových buniek (obr. 25). Keď sa dostanete dovnútra, to znamená, bližšie k osi kmeňa, spôsobia vznik drevených buniek.
Drevo sa skladá z prvkov z vodivých, podporných a základných tkanín. Medzi prvé patria drevené vlákna - dlho mŕtve bunky so zdvojenými stenami; do druhej - cievy, ktoré sú výsledkom fúzie mnohých buniek; do tretích - buniek drevnatého parenchýmu. Bunky, ktoré sa diferencujú z kambiálnych prvkov na vaskulárne bunky (cievy), rastú do dĺžky a šírky. Ich bočné steny sú zhrubnuté a lignifikované. V stenách však póry zostávajú uzavreté iba tenkou škrupinou. Priečky na spojoch susedných buniek zmiznú, protoplast odumrie. Vďaka tomu sa vytvárajú dlhé nádoby pozostávajúce z niektorých stien. Vodivý systém prechádza cez koreň a stonku do listu. Prostredníctvom takýchto nádob sa vykonáva stúpajúci tok vody a solí v nej rozpustených z koreňov do všetkých orgánov rastliny.
V strede stonky sú jadrové bunky - okrúhle alebo polyedrické parenchýmové bunky. Toto sú prvky hlavnej textílie. Niekedy sú duté a obsahujú vzduch; niekedy sú naplnené rezervnými živinami, rôznymi kryštálmi, trieslovinami. Ich steny môžu byť lignifikované. Radiálne lúče prenikajú do dreva a lákajú sa. Ich bunky sú deriváty kambia a majú akumulačnú funkciu. Na úrovni lyka sa tieto lúče nápadne rozširujú.
V dužine listov medzi hornou a dolnou epidermou je hlavné tkanivo - bunky s tenkými membránami a veľkým počtom zelených plastidov - chloroplasty. V týchto bunkách prebieha fotosyntéza. Horné vrstvy pozostávajú väčšinou z podlhovastých buniek, ktoré tesne susedia - tento stĺpcový parenchým. Spodné vrstvy sa uvoľňujú medzi bunkami, existujú medzibunkové priestory - priestory naplnené vzduchom; je to hubovitý parenchým. Do parenchýmu prenikajú rozvetvené žily, ktoré sú postavené zo zväzkov mechanických (rôzne vlákna) a vodivých (sitové trubice a vodonosné vrstvy) tkanív.
![]()
Táto krátka a schematická prezentácia plánu štruktúry stonky, listu ukazuje, aké rozmanité sú veľkosť, tvar, štruktúra a funkcie buniek tej istej rastliny. Ak vezmete rôzne rastliny, môžete sa stretnúť s ešte väčšou rozmanitosťou buniek. Spravidla sú však bunky tkanív s rovnakým názvom, dokonca aj v rastlinách, ktoré sú od seba veľmi vzdialené, podobné, pretože tieto bunky plnia podobné funkcie. Epidermálne bunky listu brezy a napríklad púpava lekárska sú si navzájom viac podobné ako epidermálna bunka púpavy lekárskej a jej sito. Charakteristické vlastnosti bunky sú spojené predovšetkým s jej špeciálnou funkciou. Bunky špecializované na mechanické funkcie majú zhrubnuté a často lignifikované membrány. Špecializácia na fotosyntézu vedie k výskytu chloroplastov v bunkách. Špecializácia na vodivú funkciu je spojená s predĺžením buniek, so stratou protoplastu, so zmenami v bunkových membránach na spojoch, kvôli ktorým komunikujú dutiny susedných buniek. Bunky špecializované na ochrannú funkciu sa vyznačujú rôznymi zmenami na vonkajších stenách, prítomnosťou chĺpkov a schopnosťou produkovať ochranné látky.
Špecializácia na funkciu skladovania živín môže viesť k zväčšeniu veľkosti buniek, k vzhľadu veľmi veľkých vakuol v nich. V bunkách meristematického, intenzívne sa deliaceho, sú vyvinuté najmä tie intracelulárne štruktúry, ktoré zabezpečujú syntézu látok - základných častí bunky.
V závislosti od povahy špecializácie sú niektoré znaky štruktúry a práce bunky vyvinuté veľmi silno, zatiaľ čo iné sú naopak rozvinuté málo alebo úplne chýbajú. Ako sme videli, niektoré bunky, ktoré sa stali špecializovanými, zomrú a je v mŕtvom stave, keď začnú vykonávať svoju špeciálnu funkciu v mnohobunkovom organizme (korkové bunky, drevené vlákna, cievy).
Bunky mnohobunkových vláknitých rias a jednobunkových rastlín sú veľmi rozmanité a zvláštne (obr. 26 a 27). Bunka ktorejkoľvek z nich sa navyše veľmi líši od buniek mnohobunkových rastlín. Ona sama musí vykonávať niekoľko funkcií, ktoré sú u mnohobunkových rastlín rozdelené medzi bunky rôznych tkanív.
Súčasne aj veľmi odlišné bunky majú hlboké podobnosti v štruktúre a funkcii. U mnohobunkových organizmov je to v prvom rade spôsobené tým, že všetky bunky tela (ak vylúčime vegetatívne rozmnožovanie, pri ktorom sa okamžite oddeľuje celý mnohobunkový komplex od materskej rastliny ako nový jedinec alebo jeho základ), sú potomkami tej istej pôvodnej bunky. Preto bez ohľadu na to, aké špecializované sú bunky, majú spoločný pôvod, a teda sú príbuznými.
Po druhé, spoločné znaky v štruktúre buniek rastlín rôznych druhov sú spojené so skutočnosťou, že všetky rastliny súvisia s tým či oným stupňom. Všetky rastliny sa vyvinuli evolúciou od bežných jednobunkových predkov. Spoločné znaky dedia bunky moderných rastlín od starodávnych progenitorových buniek. S tým súvisí aj prítomnosť spoločných znakov štruktúry a práce rastlinných a živočíšnych buniek.
Po tretie, podobnosť je spojená so skutočnosťou, že všetky živé bunky, bez ohľadu na to, akú zvláštnu funkciu môžu v tele vykonávať, si musia v prvom rade zabezpečiť svoj vlastný život. Bunky absorbujú živiny, spracúvajú ich, získavajú energiu a budujú svoje vlastné telo, dýchajú, zbavujú sa nepotrebných látok, bojujú s rôznymi škodami, reagujú na zmeny vonkajších podmienok, obnovujú svoju životne dôležitú činnosť a rastú. Všetky tieto procesy v rôznych bunkách sa uskutočňujú podobným spôsobom a pomocou štruktúr rovnakého typu, ktoré sú spoločné z hľadiska štruktúry nielen pre rôzne rastlinné bunky, ale aj pre rastlinné a živočíšne bunky. Je potrebné povedať, že výkon akejkoľvek špeciálnej funkcie bunky je založený na jej všeobecných vlastnostiach. Jedna alebo iná vlastnosť, schopnosť obsiahnutá vo všetkých bunkách, sa vyvíja obzvlášť silno v špecializovanej bunke a zaisťuje, aby bunka vykonávala svoju hlavnú, špeciálnu funkciu. Tie spoločné znaky, bez ktorých nie je možné vykonať túto špeciálnu funkciu, sa v bunke zachovajú a zvyšok sa môže stratiť. Extrémnym prípadom sú mŕtve špecializované bunky. Špeciálna funkcia takýchto buniek súvisí s ich membránou; protoplast je potrebný iba dovtedy, kým vytvorí škrupinu; potom odumrie a celá bunka sa skladá iba z neživej škrupiny, ktorá pracuje pre potreby rastliny.
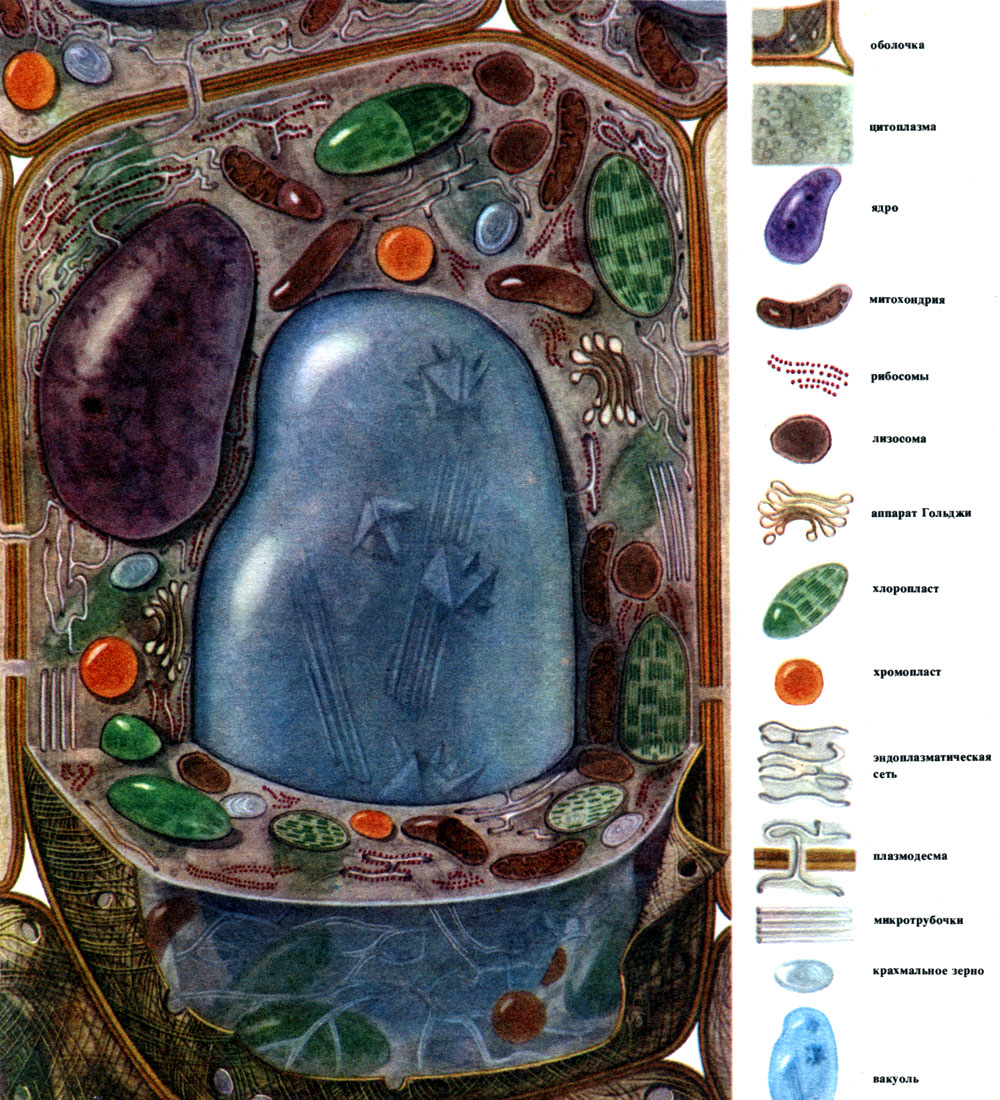
Vzhľadom na všeobecné vlastnosti štruktúry a vitálnej aktivity rastlinných buniek je vhodné hovoriť o určitej typickej bunke, ktorá absorbovala spoločné znaky rôznych špecializovaných buniek. Takáto bunka, hoci neexistuje v „čistej forme“, sa dá dokonca zobraziť (tabuľka 7, obr. 28). Zo skutočných buniek sa viac podobajú takým „typickým“ bunkám listového parenchýmu (tabuľka 8, obr. 29).
Prejdime k popisu štruktúry a práce rastlinnej bunky.
Tabuľka 5. Krytosemenné rastliny: vľavo hore - kvet magnólie; vpravo hore - kvety jabĺk; dole je lotos.
Tabuľka 6. Krytosemenné rastliny: hore (zľava doprava) - zvončeky, pelargonium, šafran (krokus); vľavo dole - astry; vpravo dole - lopúch.
Vonku je rastlinná bunka pokrytá membránou, ktorá nemá rovnakú hrúbku a štruktúru v rôznych bunkách. Látky, ktoré ju tvoria, sa vytvárajú v cytoplazme a ukladajú sa mimo nej, čím sa postupne vytvorí škrupina. Týmito látkami sú predovšetkým veľkomolekulárne polysacharidy - pektín, hemicelulóza a celulóza v malom množstve. Tvoria takzvanú primárnu škrupinu. Je dosť elastický, pretože bunka rastie, naťahuje sa a tiež rastie, a preto nezasahuje do bunkového rastu. Vytvára však určitú pevnosť bunky a je schopná ju chrániť pred mechanickým poškodením. Existujú bunky, ktoré nemajú takúto primárnu membránu - sú to bunky slúžiace na nepohlavné a sexuálne rozmnožovanie (zoospory a gaméty rias a nižších húb, mužské gamety vyšších rastlín). Mnoho buniek má nielen primárnu, ale aj sekundárnu membránu. Je tvorený pod primárnym zdrojom a je vyrobený hlavne z celulózy. Celulóza je polysacharid, ktorého molekuly tvoria najjemnejšie vlákna - mikrofibrily. V škrupine sú celulózové vlákna ponorené do amorfnej látky pozostávajúcej z pektínových zlúčenín. V niektorých bunkách sú tieto celulózové mikrofibrily umiestnené po celej dĺžke bunky v prstencoch; kvôli tomu sa také bunky môžu natiahnuť na dĺžku, ale nie na šírku (napríklad bunky ciev stonky). Ostatné majú vlákna pozdĺžne; bunky s takouto membránou sú pri pretiahnutí elastické, ale v pozdĺžnom pretiahnutí veľmi tuhé. V iných sú umiestnené šikmo a tvoria špirálu (epidermálne chĺpky z bavlníkových semien, lykové vlákna). To všetko pripomína železobetónové konštrukcie a celulózové nite tu zohrávajú úlohu železných tyčí a pektínové látky zase cement. Bunky so sekundárnou membránou sú veľmi odolné. Tvoria mechanické podporné tkanivá rastliny. Sekundárna membrána niekedy zohráva úlohu skladu výživných produktov: látky, ktoré ju tvoria, sa môžu premeniť na iné, jednoduchšie, ktoré sa konzumujú ako jedlo.
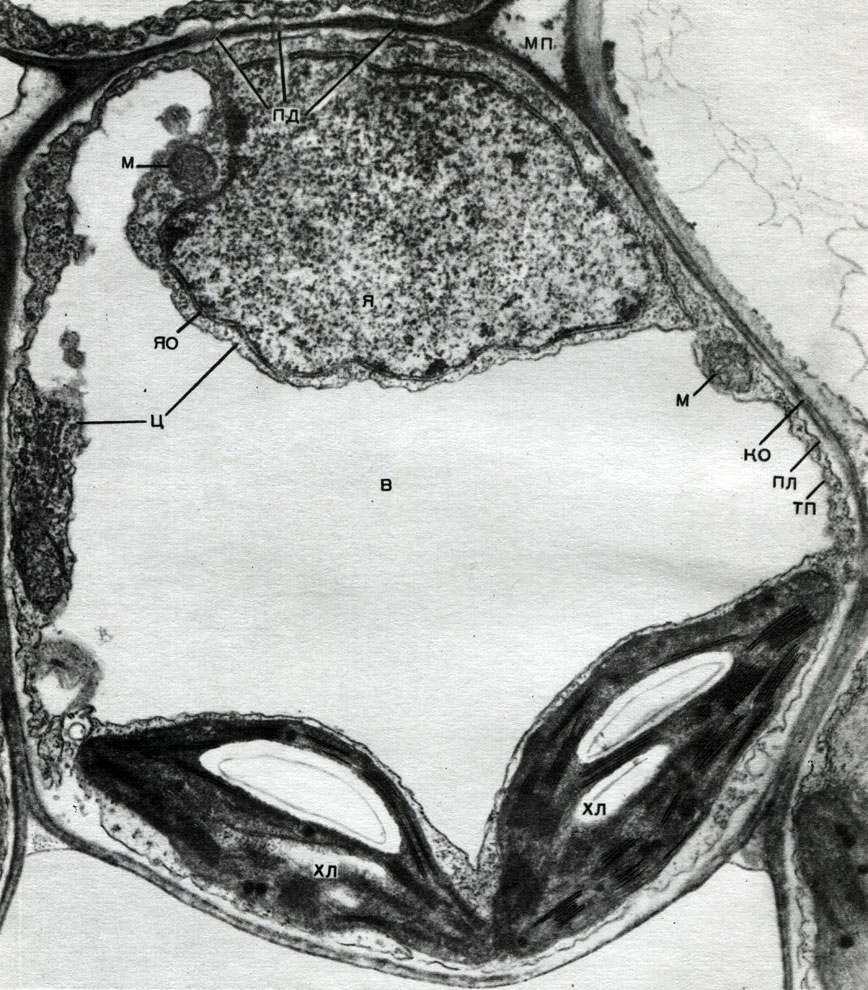
V škrupine sú nezhustnuté miesta - póry (v primárnej škrupine sa im hovorí nórové polia), cez ktoré sa uskutočňuje komunikácia medzi susednými bunkami. Cez pórovité polia a póry prechádzajú tenké povrazy cytoplazmy. Jedná sa o plazmodesmata, ktoré sa viažu na cytoplazmu susedných buniek (tabuľka 9). Vymieňajú si látky medzi susednými bunkami. Plazmodesmata spolu s prvkami vedenia tkaniva spájajú bunky a tkanivá tela do jedného celku.
Výmena látok a šírenie vzrušenia umožňujú bunkám vzájomne ovplyvňovať vývoj a prácu a každé tkanivo ovplyvňuje vitálne funkcie všetkých ostatných tkanív. Takto sa vytvára koordinácia práce všetkých častí jedného organizmu, celej rastliny.
V mnohých bunkách je bunková membrána starnutím impregnovaná látkami, ktoré ju ďalej posilňujú. Hromadenie lignínu v ňom vedie k lignifikácii škrupiny. Steny drevených buniek sú lignifikované a často sú to lykové vlákna, bunkové škrupiny semennej kože, niekedy oplodie (obr. 30) a dokonca aj plátky. Bunky s lignifikovanými stenami sú nielen silnejšie, ale aj menej priepustné pre mikróby a vodu. Membrány niektorých ďalších buniek - kožných tkanív, ako aj na povrchoch rán - sa stávajú korkovými a nasiaknutými suberínom. Obsah buniek potom odumiera, pretože suberín je nepriepustný pre vodu a plyny, ale tkanivo z týchto odumretých buniek chráni vnútorné živé časti rastliny pred škodlivými vonkajšími vplyvmi. Ako už bolo spomenuté, vonkajšie membrány epidermálnych buniek sú zvyčajne impregnované alebo pokryté kutínom a voskom, čo bunkám zabráni v nadmernom odparovaní vody a pred infekciou. Plášť môže byť tiež impregnovaný minerálmi, hlavne soľami vápnika.
V živote buniek však existujú obdobia, keď by ich membrána mala byť menej odolná ako predtým. Stáva sa to napríklad počas klíčenia semien.
Polysacharidová bunková membrána je charakteristickým znakom štruktúry rastlinnej bunky, ktorá ju odlišuje od živočíšnej bunky.
Membrána alebo bunková stena je ochranný útvar. Pod membránou je cytoplazma. Jeho najvzdialenejšou vrstvou, susediacou s membránou, je membrána povrchových buniek - plazmalemma. Je to kombinácia vrstiev tukových a bielkovinových molekúl. Takéto membrány sa nazývajú lipoproteínové membrány („lipos“ - tuk, „proteín“ - proteín). Membrána tohto dizajnu vymedzuje cytoplazmu z vakuol; táto membrána sa nazýva tonoplast. Mnoho bunkových organel je tvorených z lipoproteínových membrán. Avšak v každom prípade je membrána postavená z tukovitých látok (lipidov) a bielkovín obsiahnutých v tejto konkrétnej membráne. Kvalitatívna rozmanitosť lipidov a najmä proteínov je kolosálna, a teda obrovská rozmanitosť membrán líšiacich sa vlastnosťami, a to tak v rámci jednej bunky, ako aj v rôznych bunkách.
Plazmalemma reguluje vstup látok do bunky a ich výstup z nej, zaisťuje selektívny prienik látok do a z bunky. Rýchlosť penetrácie cez membránu rôznych látok je rôzna. Cez ňu dobre preniká voda a plynné látky. Látky rozpustné v tukoch tiež ľahko prenikajú, pravdepodobne kvôli tomu, že má lipidovú vrstvu. Predpokladá sa, že lipidová vrstva membrány je preniknutá pórmi. To umožňuje látkam nerozpustným v tukoch prejsť cez membránu. Póry nesú elektrický náboj, takže prienik iónov cez ne nie je úplne voľný. Za určitých podmienok sa náboj pórov mení a tým sa reguluje permeabilita membrán pre ióny. Membrána však nie je rovnako priepustná pre rôzne ióny s rovnakým nábojom a pre rôzne nenabité molekuly blízkej veľkosti. Toto je prejav najdôležitejšej vlastnosti membrány - selektivity jej priepustnosti: pre niektoré molekuly a ióny je lepšie priepustná, pre iné horšia.
Látka sa pohybuje do bunky a difunduje do nej, ak je jej koncentrácia vonku vyššia ako koncentrácia vo vnútri. V opačnom prípade by malo z bunky difundovať. Ale po preniknutí do bunky môžu byť molekuly tejto látky okamžite zapojené do metabolizmu a podrobené transformácii na iné látky. Potom koncentrácia tejto látky vo vnútri bunky opäť poklesne a nová časť jej molekúl difunduje do bunky. Výsledkom je, že bunky, ktoré aktívne konzumujú túto alebo tú látku, tieto intenzívne vstupujú z okolitých buniek.
Ďalším mechanizmom na zvýšenie absorpcie látky bunkou je jej väzba na bielkoviny alebo iné látky. Väzbou látka opúšťa intracelulárny roztok a neruší ďalšiu difúziu tej istej látky. Ďalej vstupuje do bunky a hromadí sa v nej vo viazanej forme. Takže v bunkách niektorých morských rias sa jód hromadí v koncentráciách miliónkrát vyšších ako je ich koncentrácia v morskej vode.
Vo všetkých týchto prípadoch hovoríme o pohybe látok difúziou. Bunkové membrány však majú aj mechanizmy na presun látok cez seba z nižšej koncentrácie na vyššiu. Medzi bielkovinami membrány sú bielkovinové látky, ktorých úlohou je presunúť rôzne látky z jednej strany membrány na druhú. Nazývajú sa transportné enzýmy. Pravdepodobne taký transportný enzým vytvára medziprodukt s transportovanou látkou; ten prechádza membránou, potom sa rozpadne na svoje pôvodné časti, potom sa enzým vráti späť a prenesená látka zostáva na druhej strane membrány. Podrobnosti o tomto procese nie sú známe, ale samotný fakt takého aktívneho transportu látok je nepochybný. Na rozdiel od difúzneho, pasívneho, enzymatického transportu je potrebný energetický výdaj. Pravdepodobne je to vďaka tomuto aktívnemu transportu iónov, že bunky koreňovej epidermy sú schopné absorbovať anorganické látky potrebné pre rastlinu z pôdy a potom ich prenášať cez rastlinu z bunky do bunky.
Chemickými alebo fyzikálnymi zmenami vo vonkajšom a vnútornom prostredí menia bunkové membrány svoju priepustnosť, ako aj stupeň a samotnú povahu jej selektivity. Na tomto sú založené mechanizmy regulácie pohybu látok do a z bunky. Zmena v permeabilite membrán pre živiny sa odráža v intenzite metabolických procesov v bunke, na povahe syntéz, ktoré sa v nej vyskytujú, a počas celého jej života. Zmena permeability membrán pre ióny sodíka a draslíka je mechanizmom šírenia bunkovej excitácie - vzniku a pohybu bioprúdu. Na povrch membrány je pripojené veľké množstvo katiónov, hlavne iónov draslíka. Preto nesie zvonku kladný náboj. Pod vplyvom podnetov sa z postihnutej oblasti odštepujú ióny draslíka, náboj oblasti klesá a stáva sa elektronegatívnym vzhľadom na susedné oblasti membrány. Táto elektronegativita je zase dráždivá pre susedné oblasti, rovnakým spôsobom odstraňuje ich náboj a depolarizuje ich. Vlna elektronegativity sa šíri cez membránu - to je biologický prúd. Potom ióny draslíka opäť sedia na membráne a dávajú jej počiatočný náboj - po vlne depolarizácie nasleduje vlna obnovy. Takéto membrány tiež tvoria povrchy plazmodesmata prechádzajúce bunkovými stenami a spájajúce cytoplazmu susedných buniek. Pozdĺž nich sa pohybuje aj bioprúd, ktorý sa šíri z bunky do bunky. Bioprúdy rastlinných buniek sú stále zle pochopené. Je však zrejmé, že ide o signalizačnú metódu používanú v rastlinnej bunke na spustenie niektorých chemických reakcií a inhibíciu iných. Faktom je, že všetky chemické procesy prebiehajúce v bunke a tvoriace podstatu jej životne dôležitej činnosti sa uskutočňujú za účasti biologických katalyzátorov - enzýmov. Každá reakcia je možná, iba ak je aktívny enzým, ktorý ju spúšťa. Väčšina enzýmov sa aktivuje pôsobením určitých anorganických katiónov: K., Na. , Ca .., Mg .., Mn ... Bioprúd, ktorý mení permeabilitu membrány pre nabité látky, vytvára podmienky pre ich penetráciu cez membrány a kontakt s enzýmami. Bioprúd teda aktivuje určité enzýmy, a tým reguluje a usmerňuje metabolizmus v bunke. Regulácia metabolizmu bioprúdmi je len jedným z mnohých spôsobov regulácie intracelulárneho metabolizmu.
To, čo sa už hovorilo o priepustnosti povrchovej membrány bunky - plazmalemme - platí aj pre iné intracelulárne membrány, vrátane membrán, z ktorých je vybudovaných veľa bunkových organel.
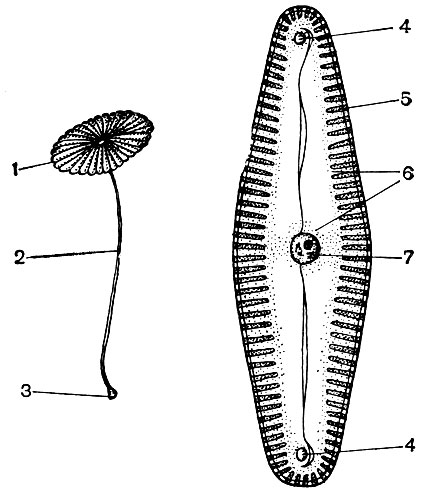
Cytoplazma, ktorá sa kedysi považovala za homogénny koloidný roztok bielkovinových látok, je v skutočnosti zložito štruktúrovaná. S rozvojom mikroskopických výskumných techník sa objasňovali čoraz jemnejšie detaily štruktúry cytoplazmy. V cytoplazme boli objavené rôzne organely (organely) - štruktúry, z ktorých každá plní určité fyziologické a biochemické funkcie. Najdôležitejšie organely cytoplazmy sú mitochondrie, endoplazmatické retikulum (endoplazmatické retikulum), Golgiho aparát, ribozómy, plastidy, lyzozómy. Pohyblivé bunky (zoospory a gamety rias, spermie prasličky, papradie, cykasy, niektoré jednobunkové a koloniálne riasy) majú pohybové organely - bičíky.
Obzvlášť veľa nových poznatkov o jemnej štruktúre cytoplazmy prinieslo a prináša elektrónový mikroskop, ktorý umožňuje študovať podrobnosti o štruktúre samotných organel. Moderné biofyzikálne a biochemické metódy umožňujú izolovať určité organely cytoplazmy v čistej forme a potom ich študovať chemické zloženie a ich funkcie. Mimo bunky, v prostredí so zložitým zložením, je mnoho organel schopných vykonávať prácu, keď sú v bunke.
Časť cytoplazmy, v ktorej sú organely ponorené, a ktorá sa zatiaľ javí ako štrukturálna, sa nazýva základná látka cytoplazmy alebo hyaloplazmy. Hyaloplazma nie je v žiadnom prípade pasívnym plnivom, ale aktívne pracujúcou časťou cytoplazmy. Prebieha v ňom množstvo životne dôležitých chemických procesov, obsahuje veľa enzýmových proteínov, pomocou ktorých sa tieto procesy uskutočňujú.
Život buniek pozostáva z nepretržitej chemickej práce, ktorá sa ako celok nazýva metabolizmus. Bunka je v podstate chemická továreň, ktorá vyrába širokú škálu produktov a nezávisle na sebe extrahuje energiu potrebnú na ich výrobu. Jej produkty sú látky, ktoré sama potrebuje na udržanie vlastného života (na budovanie tela počas rastu a vývoja, na výmenu opotrebovaných častí) a na vytváranie dcérskych buniek počas reprodukcie a pre potreby ďalších buniek v tele.
Všetky chemické reakcie prebiehajúce v bunke možno rozdeliť do dvoch skupín. Výsledkom je, že niektoré z týchto alebo týchto látok sa rozpadajú na menšie molekuly. V dôsledku ďalších malomolekulárnych látok sa syntetizujú látky s väčšími molekulami. Molekula akejkoľvek látky sa skladá z atómov, ktoré sú držané pohromade chemickými väzbami, to znamená určitým množstvom koncentrovanej potenciálnej chemickej energie. Keď sa molekula rozpadne, väzby sa rozbijú a ich chemická energia sa uvoľní. Pre syntézu, to znamená pre formovanie väčšej molekuly z malých, je potrebné vytvárať nové chemické väzby, Je potrebné vložiť do nich určitú časť energie. Naopak, rozkladné reakcie nakoniec prebiehajú s uvoľňovaním energie, pretože sa počas nich porušujú väzby medzi atómami. Biologický význam rozkladných reakcií prebiehajúcich v bunke je ten, že uvoľňujú chemickú energiu, ktorú potom bunka používa na syntézne reakcie a na výrobu ďalších typov práce (elektrická, mechanická, práca na prepravu látok). Pretože bunka počas svojho života syntetizuje rôzne látky, musí neustále ďalšie látky rozkladať. Uvoľnená energia sa okamžite nepoužíva na syntézy. Najskôr sa ukladá tvorbou špeciálnych látok - akumulátorov chemickej energie - kyseliny adenozíntrifosforečnej (ATP) a príbuzných zlúčenín. V správnom čase a vo vhodnom bode bunky sa ATP štiepi a vzdáva sa energie na syntézu látky potrebnej pre bunku.
Všetky chemické reakcie v bunke - syntéza aj rozpad - sa uskutočňujú pomocou enzýmov. Enzýmy sú bielkovinové látky, ktoré urýchľujú priebeh reakcií. Toto zrýchlenie je také veľké, že bez enzýmov by také reakcie v bunke neboli vôbec možné. Existujú prípady, kedy sa vďaka enzýmu reakcia zrýchli 1011-krát. To znamená, že reakcia, ktorá sa skončí účasťou enzýmu do 0,01 sekundy, bez nej, by pokračovala 31 rokov. Je zrejmé, že také reakcie bez enzýmu by boli jednoducho nereálne.
Vďaka enzýmom je navyše riadený a regulovaný priebeh chemických reakcií v bunke. Aktivita enzýmov v bunke sa mení v súlade s jej potrebou určitej látky alebo energie, to znamená v dôsledku práce práve týchto enzýmov. V prípade potreby látky sa tieto enzýmy, vďaka ktorým vzniká, znova aktivujú alebo syntetizujú. Tvorba látok, uvoľňovanie a ukladanie energie je výsledkom postupných reakcií, výsledkom práce celého reťazca enzýmov. Všetky štruktúry živých častí bunky sú postavené presne z enzýmov a z látok, ktoré tieto enzýmy držia pohromade. V tomto prípade sú enzýmy zúčastňujúce sa susedných, postupne prebiehajúcich reakcií, umiestnené vedľa seba. Prenášajú medzi sebou molekuly látok, ktoré premieňajú, akoby pozdĺž dopravníka, a každý z nich vykonáva na molekule svoju vlastnú pracovnú operáciu.
Hyaloplazma obsahuje enzýmy, ktoré štiepia molekuly glukózy na jednoduchšie molekuly kyseliny pyrohroznovej. Energia uvoľnená v tomto prípade je uložená tvorbou molekúl ATP. Rovnaký proces prebieha aj v bunkovom jadre. Väčšina energie sa však získava v špeciálnych organelách cytoplazmy - mitochondriách, pretože dochádza k hlbšiemu rozkladu látok.
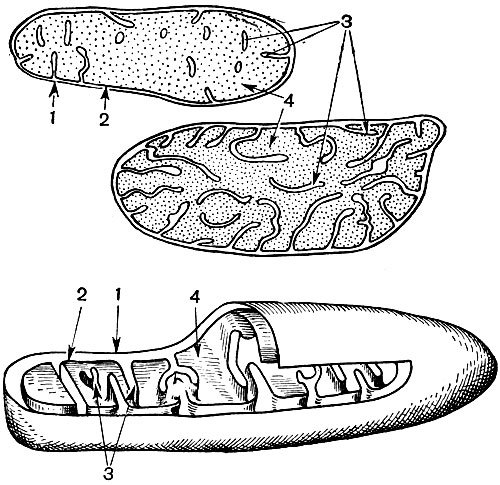
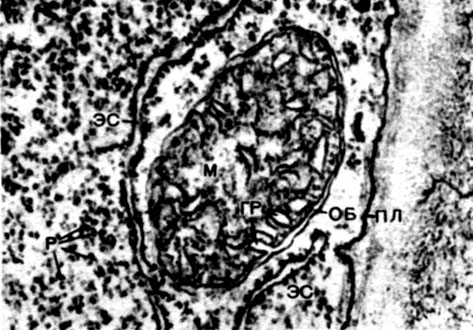
Mitochondrie sú malé telieska zaobleného alebo podlhovastého tvaru, veľké 0,5 - 1,5 mikrónu, to znamená veľkosť baktérie. Ich počet v bunke je zvyčajne veľký, asi 100 - 3 000. Existujú však bunky s malým počtom mitochondrií. Spermie morských rias fucus teda obsahuje iba 4 mitochondrie a v jednobunkových riasach mikromonas - jednu. Mitochondrie sú viditeľné pod svetelným mikroskopom, ale ich jemnú štruktúru je možné študovať iba pomocou elektrónového mikroskopu (tabuľka 10, diagram štruktúry - obr. 31). Mitochondrie sú formácie postavené z lipoproteínových membrán ponorených do základnej látky - matrice. Mitochondriálna membrána je tvorená dvoma membránami, medzi ktorými je medzera.
Vnútorná membrána škrupiny poskytuje vo vnútri početné invaginácie, to sú kristá. Medzi nimi je matica. Ako vnútorná membrána mitochondriálnej membrány, tak aj kristy z nej tvorené sú zostavené z usporiadania umiestnených enzýmov. Vďaka záhybom - cristae je pracovná plocha membrán vo vnútri mitochondrií veľmi veľká. V mitochondriálnej matrici, teda medzi kristami, sa nachádza množstvo enzýmov.
Kombinácia týchto enzýmov uskutočňuje intracelulárne dýchanie a ukladanie energie uvoľnenej počas dýchania vo forme ATP. Práca mitochondrií úzko súvisí s procesmi prebiehajúcimi v hyaloplazme, kde prebiehajú prvé stupne rozkladu glukózy a iných látok na kyselinu pyrohroznovú. V mitochondriách dochádza k jeho ďalšiemu štiepeniu. Kyselina pyrohroznová preniká do mitochondrií a tu sa krok za krokom oxiduje na oxid uhličitý a vodu a súčasne sa spotrebúva kyslík. Jedná sa o intracelulárne dýchanie, pri ktorom bunka, štiepiace a oxidujúce látky, produkuje veľa energie, ktorú potom môže použiť na rôzne svoje potreby.
Prvý stupeň rozkladu molekuly glukózy, počas ktorého je rozdelená na polovicu a prebieha v hyaloplazme, dáva bunke iba dve molekuly ATP.
Výsledkom druhého stupňa vedúceho k úplnému „spáleniu“ glukózy je ďalších 36 molekúl ATP. Preto sú mitochondrie svojou funkciou elektrárne bunky, stroje na získavanie väčšiny energie. Samotný rozklad produktov rozkladu glukózy nastáva v mitochondriálnej matrici, zatiaľ čo ATP sa tvorí v dôsledku reakcií prebiehajúcich na jeho vnútorných membránach, ktoré zahŕňajú dýchacie enzýmy a enzýmy, ktoré zabezpečujú tvorbu ATP. Počet kristí v mitochondriách sa môže líšiť. Čím viac z nich, tým vyššia je biochemická aktivita mitochondrií.
Hovoríme tu o glukóze ako látke, ktorej štiepenie bunka produkuje energiu. Glukóza je centrálna, ale nie jediná takáto látka. Jeho molekula má hlavný reťazec šiestich atómov uhlíka, ktoré sú navzájom spojené. V dôsledku dlhého a zložitého reťazca reakcií sa jeho molekula rozdrví, oxiduje a nakoniec sa rozdelí na šesť molekúl anorganickej látky - oxidu uhličitého (CO 2), z ktorých každá obsahuje iba jeden atóm uhlíka, a je extrémne oxidovaná. Sčítaním všetkých po sebe nasledujúcich reakcií oxidácie glukózy a vylúčením všetkých medziproduktov môžete získať celkovú reakciu tohto procesu:
Škrob sa ľahko premieňa na glukózu a potom prechádza vyššie opísanou degradáciou. Bielkoviny a tuky poskytujú rôzne organické kyseliny, ktoré sa pomocou rovnakých enzýmov premieňajú na medziprodukty štiepenia glukózy a potom sa oxidujú rovnakým spôsobom ako glukóza.
Úplná biologická oxidácia organických látok je podobná ich spaľovaniu. Výsledkom je v obidvoch prípadoch uvoľnený oxid uhličitý, voda a energia. Počas spaľovania sa však táto energia uvoľňuje vo forme tepla a to úplne naraz; počas biologickej oxidácie sa energia chemických väzieb uvoľňuje po častiach a jej hlavná časť je viazaná a prechádza do energie fosfátovej chemickej väzby ATP. Výsledkom je, že bunka dostane koncentrát energie v takejto forme, ktorý sa potom v správnom čase a v príslušnom okamihu môže použiť na vytvorenie nových chemických väzieb, na syntézu nových látok, ako aj na výrobu iných druhov práce - elektrickej, mechanickej a tiež na prácu pri preprave látok z prostredia do bunky, z bunky do prostredia a z bunky do bunky.
Čím aktívnejšia je vitálna aktivita bunky, tým väčšia je jej potreba energie a tým viac mitochondrií obsahuje. Môžu byť nerovnomerne rozložené v tej istej bunke: v časti bunky, ktorá v súčasnosti pracuje aktívnejšie, je ich viac.
Mitochondrie sú schopné syntetizovať niektoré látky, z ktorých sa samy skladajú. Vďaka tomu sa môžu mitochondrie množiť.
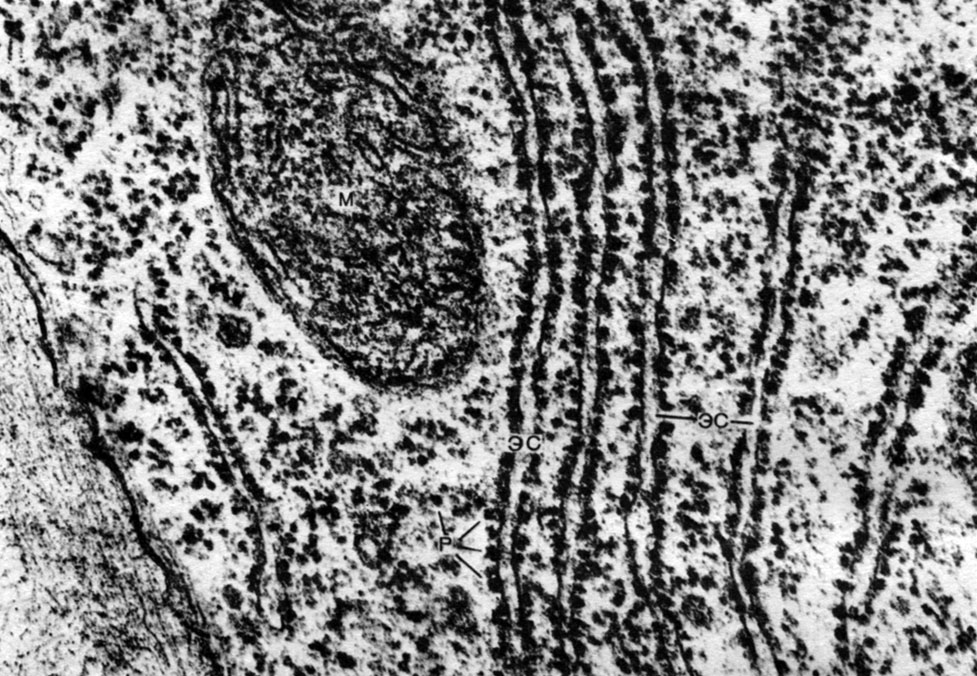
Endoplazmatické retikulum je organoid cytoplazmy, v ktorom sa syntetizuje veľké množstvo látok (tabuľka 10). Endoplazmatické retikulum je systém kanálov, ktoré prestupujú cytoplazmu a ktoré sa v niektorých oblastiach zužujú, v iných sa rozširujú a vytvárajú buď cisterny, ploché vaky alebo rozvetvovacie trubice. Steny všetkých týchto formácií sú postavené z membrán, ktoré obsahujú enzýmy.
Rovnako ako v iných membránových formáciách bunky sú enzýmy v retikulu usporiadané usporiadane. V takom prípade susedné enzýmy uskutočňujú postupne reakcie (pracovné operácie) a ich skupina vykonáva celý reťazec reakcií vedúcich k vytvoreniu konkrétnej látky.
Rozlišujte medzi agranulárnym (hladkým) a granulovaným endoplazmatickým retikulom. Na vonkajšom povrchu kanálov granulovaného retikula sa nachádzajú početné malé organely - ribozómy, ktorých funkciou je syntéza proteínových molekúl. Agranulárne endoplazmatické retikulum, ktoré kvantitatívne prevláda nad granulovaným v rastlinnej bunke, nenesie ribozómy.
Endoplazmatické retikulum je nielen dopravníkom mnohých druhov enzymatickej premeny látok, hlavne na ich syntézu, ale aj sústavou diaľnic, po ktorých sa látky pohybujú bunkou. Retikulum začína od vonkajšej membrány membrány jadra a vetvením sa približuje k rôznym organelám cytoplazmy, ako aj k plazmalemme. Spája teda všetky časti bunky navzájom. Okrem toho kanály endoplazmatického retikula prechádzajú cez plazmodesmata a spájajú retikulum susedných buniek.
Ďalej membrány endoplazmatického retikula rozdeľujú cytoplazmu na početné oddelenia, vďaka čomu nemôže byť bunka predstavovaná ako homogénna hmota, v ktorej je zmiešané množstvo látok. Podmienky v jednom oddelení sa môžu úplne líšiť od ostatných; procesy prebiehajúce v ňom môžu prebiehať iba v ňom, zatiaľ čo navzájom prebiehajú ďalšie procesy.
Nakoniec sú membrány endoplazmatického retikula tie povrchy, pozdĺž ktorých sa šíria bioprúdy, čo sú signály, ktoré menia selektívnu permeabilitu membrán, a tým aktivitu enzýmov. Vďaka tomu sa niektoré chemické reakcie uvedú do pohybu, iné sú inhibované - metabolizmus podlieha regulácii a prebieha koordinovane.
Mnoho látok syntetizovaných v bunke musí byť koncentrovaných a uvoľnených z bunky buď do vonkajšieho prostredia, alebo do intracelulárnej vakuoly. Bunka navyše koncentruje aj látky, ktoré sa do nej dostávajú z iných buniek, napríklad ak si ich ukladá do rezervy. Túto prácu vykonávajú diktyozómy. Zvyčajne v rastlinnej bunke existuje niekoľko diktyozómov a celá ich sada sa nazýva Golgeov prístroj (alebo komplex) a daná bunka. Každý diktyozóm je systém skladaných membrán (tabuľka 10 - rez, obr. 32 - volumetrický diagram). Dutiny medzi membránami, ktoré tvoria tento organoid, vyzerajú ako úzke štrbiny, potom ako ploché vaky - cisterny, potom bubliny. Ich tvar sa mení počas činnosti organoidu a zjavne závisí od stupňa naplnenia medzimembránových priestorov vylučovanými a nahromadenými látkami. Vytvorené a prerastené vezikuly sú oddelené od organoidu. Zdá sa, že veľa bunkových vakuol obklopených membránami - tonoplastom je produktom činnosti Golgiho aparátu; to sú bubliny od nej oddelené a potom zväčšené.
Golgiho aparát je vyvinutý najmä vo vylučovacích (sekrečných) bunkách, v ktorých sa ukladajú rôzne látky alebo z ktorých sa vylučujú rôzne látky. Syntetizuje a uvoľňuje látky, ktoré tvoria bunkovú stenu.
Lyzozómy sú pomerne malé (priemer asi 0,5 mikrónu) zaoblené telieska - ďalší organoid cytoplazmy. Sú pokryté membránou - lipoproteínovou membránou. Obsahom lyzozómov sú enzýmy, ktoré trávia bielkoviny, sacharidy, nukleové kyseliny a lipidy. Lyzozómová membrána zabraňuje uvoľňovaniu enzýmov z organoidu do hyaloplazmy, inak by sa tieto enzýmy trávili.
Dá sa predpokladať, že lyzozómy sú produktmi činnosti Golgiho aparátu, od ktorých sa oddeľujú bubliny, v ktorých tento organoid nahromadil tráviace enzýmy.
Tie časti bunky, ktoré počas vývoja odumierajú, sú zničené lyzozomálnymi enzýmami. V zosnulej bunke sú lyzozómy zničené, enzýmy sa nachádzajú v cytoplazme a celá bunka, s výnimkou membrány, podlieha tráveniu.
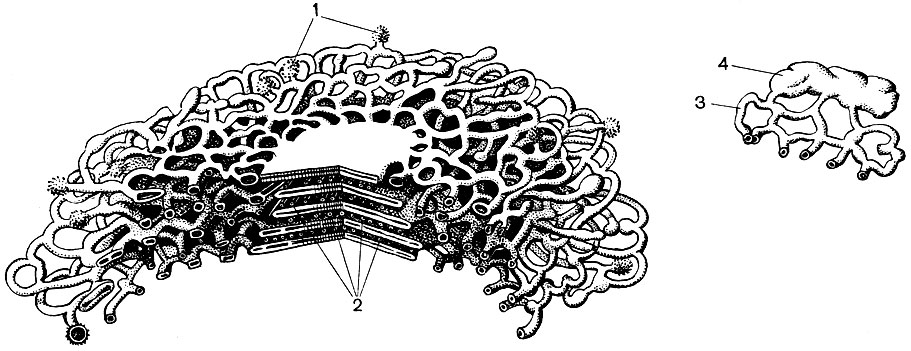
Ribozómy sú veľmi malé organely s priemerom asi 250 A. Majú takmer sférický tvar. Niektoré z nich sú pripevnené k vonkajšiemu (hyaloplazmatickému) povrchu membrán, ktoré tvoria kanály granulovaného endoplazmatického retikula; niektoré sú v hyaloplazme vo voľnom stave. Bunka môže obsahovať až 5 miliónov ribozómov. Sú „prístrojom na syntézu bielkovín. Preto je ich obzvlášť veľa v bunkách, ktoré aktívne tvoria bielkoviny - v rastúcich bunkách, v bunkách vylučujúcich bielkovinové látky. Ribozómy sa nachádzajú aj v mitochondriách a chloroplastoch, kde syntetizujú časť proteínov, z ktorých tieto organely.
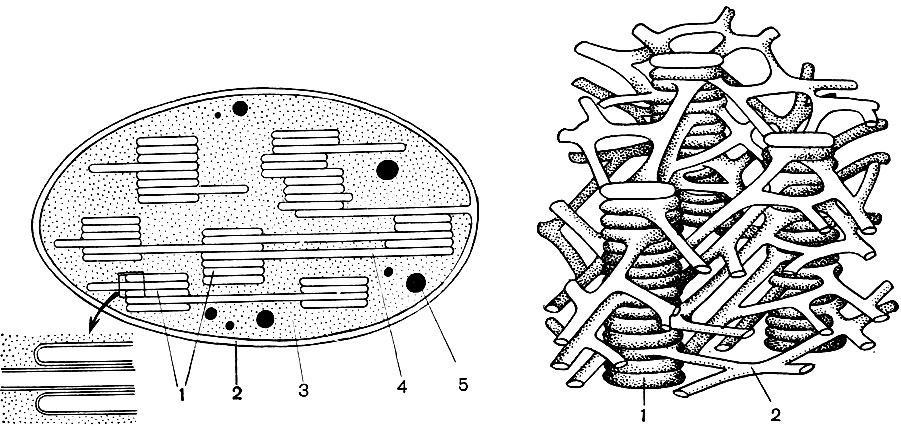
Organely nazývané mikrotubuly sa nachádzajú v mnohých bunkách. Ich samotný názov hovorí o ich tvare - jedná sa o trubice s kanálom vo vnútri. Ich vonkajší priemer je asi 250A. Niekedy sú to dvojité trubice - dve jednoduché, ktoré ležia vedľa seba a majú spoločnú stenu, ktorá oddeľuje ich dutiny. Steny mikrotubulov sú postavené z bielkovinových molekúl. Predpokladá sa, že mikrotubuly sú spojené s kontraktilnou (motorickou) aktivitou cytoplazmy a jej formácií. Z nich, ako aj zo stavebných častí, sú zjavne postavené kontraktilné štruktúry bičíka, organoidu, pomocou ktorého sa pohybujú niektoré jednobunkové a koloniálne riasy, ako aj bunky slúžiace na reprodukciu mnohých nižších rastlín. Počas delenia buniek sa z mikrotubulov vytvárajú vretenové vlákna, o ktorých bude pojednané nižšie. Počas obdobia delenia sa mikrotubuly zhromažďujú v skupinách a vytvárajú tieto vlákna. Na konci delenia sa vlákna opäť rozpadajú na samostatné mikrotubuly. V bunkách alebo ich častiach, ktoré nemajú hustú membránu, majú mikrotubuly pravdepodobne podpornú funkciu, ktorá tvorí vnútornú kostru bunky.
Plastidy sú organely vlastné iba rastlinným bunkám. Spravidla sú to veľké telá, zreteľne viditeľné pod svetelným mikroskopom.
Existujú 3 typy plastidov: bezfarebné - leukoplasty, zelené - chloroplasty, sfarbené inými farbami - chromoplasty. Každý typ plastidov má svoju vlastnú štruktúru a nesie svoje vlastné inherentné funkcie. Prechody plastidov z jedného typu na druhý sú však možné. Ekologizácia zemiakových hľúz je teda spôsobená preskupením ich leukoplastov do chloroplastov. V koreňovej plodine mrkvy prechádzajú leukoplasty do chromoplastov. Všetky tri typy plastidov sú tvorené z proplastidov.
Proplastidy sú bezfarebné telieska, podobné mitochondriám, ale o niečo väčšie ako oni. Nachádzajú sa vo veľkom množstve v meristematických bunkách. Leukoplasty sa nachádzajú v bunkách nefarbených častí rastlín (ovocie, semená, korene, listová pokožka). Ich forma je neurčitá. Najčastejšie existujú leukoplasty, v ktorých je uložený škrob (je tvorený z cukrov). Existujú leukoplasty, ktoré ukladajú bielkoviny. Najmenej bežné sú tukové leukoplasty; vznikajú počas starnutia chloroplastov. Medzi leukoplastmi a proplastidmi nie sú významné rozdiely.
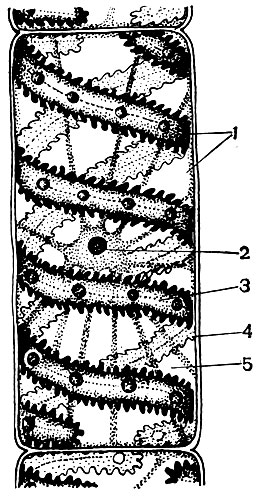
Chloroplasty sú plastidy vyšších rastlín, v ktorých prebieha proces fotosyntézy, to znamená využitie energie svetelných lúčov na tvorbu organických látok z anorganických látok (oxid uhličitý a voda) so súčasným uvoľňovaním kyslíka do atmosféry. Chloroplasty majú formu bikonvexnej šošovky, ich veľkosť je asi 4 - 6 mk. Nachádzajú sa v narenchymálnych bunkách listov a iných zelených častiach vyšších rastlín. Ich počet v bunke sa pohybuje od 25 do 30.
Štruktúra chloroplastov je uvedená v tabuľkách 11 a 12 (pohľad v reze) a na diagrame na obrázku 33. Chloroplast je zvonka pokrytý membránou pozostávajúcou z dvoch lipoproteínových membrán. Pod ním sú v hlavnej látke (stróma) usporiadané početné útvary - lamely - usporiadane. Tvoria ploché vrecká, ktoré ležia na sebe v pravidelných stoličkách. Títo stoici, ktorí sa podobajú na mince zložené do stĺpca, sa nazývajú zrná. Prechádzajú nimi dlhšie lamely, takže všetky zrná chloroplastov sú spojené do jedného systému. Zloženie membrán tvoriacich granule zahŕňa zelený pigment - chlorofyl. Práve tu prebiehajú svetelné reakcie fotosyntézy - absorpcia svetelných lúčov chlorofylom a premena svetelnej energie na energiu excitovaných elektrónov. Elektróny excitované svetlom, to znamená tie, ktoré majú prebytočnú energiu, sa vzdávajú energie na rozklad vody a syntézu ATP. Pri rozklade vody vzniká kyslík a vodík. Do atmosféry sa uvoľňuje kyslík a vodík je viazaný proteínom ferredoxínom. Ferredoxín sa potom znovu oxiduje, pričom sa tento vodík uvoľňuje do redukčného činidla, v skratke NADP. NADP ide do redukovanej formy - NADP-H 2. Výsledkom svetelných reakcií fotosyntézy je teda tvorba ATP, NADP-H 2 a kyslíka a voda a svetelná energia sa spotrebúvajú.
ATP akumuluje veľa energie - potom sa používa na syntézu, ako aj na ďalšie potreby bunky. NADP-H 2 je akumulátor vodíka a ľahko ho potom rozdáva. Preto je NADP-H2 chemickým redukčným činidlom. S redukciou súvisí veľké množstvo biosyntéz a NADPH2 pri týchto reakciách pôsobí ako dodávateľ vodíka.
Ďalej pomocou stromálnych enzýmov chloroplastov, to znamená mimo granúl, dochádza k temným reakciám: vodík a energia obsiahnutá v ATP sa používajú na zníženie atmosférického oxidu uhličitého (CO2) a na jeho zahrnutie do zloženia organických látok. Prvá organická hmota, ktorá vznikla v dôsledku fotosyntézy, prechádza veľkým počtom prešmykov a vedie k množstvu rôznych organických látok syntetizovaných v rastline a z jej tela. Mnohé z týchto transformácií prebiehajú práve tam, v stróme chloroplastov, kde sú enzýmy na tvorbu cukrov, tukov a všetko potrebné na syntézu bielkovín. Cukry potom môžu buď prechádzať z chloroplastov do iných bunkových štruktúr a odtiaľ do ďalších rastlinných buniek, alebo vytvárať škrob, ktorého zrná možno často vidieť v chloroplastoch. Tuky sa tiež ukladajú v chloroplastoch alebo vo forme kvapiek, alebo vo forme jednoduchších látok, prekurzorov tukov, sa uvoľňujú z chloroplastov.
Komplikácia látok je spojená s vytváraním nových chemických väzieb a zvyčajne vyžaduje energiu. Jeho zdrojom je rovnaká fotosyntéza. Faktom je, že významná časť látok vytvorených v dôsledku fotosyntézy sa opäť rozkladá v hyaloplazme a mitochondriách (v prípade úplného spaľovania na látky, ktoré slúžia ako východiskový materiál pre fotosyntézu - CO2 a H2O). Výsledkom tohto procesu, ktorý je v podstate zvrátením fotosyntézy, je energia, ktorá sa predtým nahromadila v chemických väzbách rozložených látok, a tá sa - opäť pomocou ATP - vynakladá na tvorbu nových chemických väzieb syntetizovaných molekúl. Významná časť výroby fotosyntézy je teda potrebná iba na to, aby sa na ňu naviazala energia svetla a potom, čo sa z nej stala chemická látka, sa použije na syntézu úplne iných látok.
A ako stavebný materiál pre tieto syntézy sa používa iba časť organickej hmoty tvorenej počas fotosyntézy.
Výroba fotosyntézy (biomasy) je kolosálna. Za rok na planéte to je asi 1010 ton. Organické látky vytvorené z rastlín sú jediným zdrojom života nielen pre rastliny, ale aj pre zvieratá, pretože tieto druhy spracúvajú hotové organické látky, pričom sa živia priamo rastlinami alebo inými zvieratami, ktoré: zase sa živia rastlinami. Fotosyntéza je teda jadrom všetkého moderného života na Zemi. Všetky transformácie látok a energie v rastlinách a živočíchoch sú preskupenia, rekombinácie a prenosy hmoty a energie primárnych produktov fotosyntézy. Fotosyntéza je dôležitá pre všetko živé a skutočnosť, že jedným z jej produktov je voľný kyslík pochádzajúci z molekuly vody a uvoľňovaný do atmosféry. Predpokladá sa, že všetok kyslík v atmosfére pochádza z fotosyntézy. Je nevyhnutný pre dýchanie rastlín i zvierat.
Chloroplasty sú schopné sa pohybovať po bunke. Za slabého osvetlenia sú umiestnené pod bunkovou stenou, ktorá smeruje k svetlu. Pri tom sa zväčšujú na väčšiu plochu. Ak je svetlo príliš intenzívne, stočia sa k nemu a zoradí sa pozdĺž tieňov rovnobežných so svetelnými lúčmi. Pri strednom osvetlení zaujímajú chloroplasty polohu medzi dvoma krajnými. V každom prípade sa dosiahne jeden výsledok: chloroplasty sú v najpriaznivejších svetelných podmienkach pre fotosyntézu. Takýto pohyb chloroplastov (fototaxia) je prejavom jedného z druhov dráždivosti rastlín.
Chloroplasty majú v bunkovom systéme určitú autonómiu. Majú svoje vlastné ribozómy a súbor látok, ktoré určujú syntézu množstva vlastných proteínov chloroplastov. Existujú aj enzýmy, ktorých práca vedie k tvorbe lipidov tvoriacich lamely, a chlorofylu. Ako sme videli, chloroplast má tiež autonómny systém výroby energie. Vďaka tomu všetkému sú chloroplasty schopné samostatne budovať svoje vlastné štruktúry. Existuje dokonca názor, že chloroplasty (ako mitochondrie) pochádzajú z niektorých nižších organizmov, ktoré sa usadili v rastlinnej bunke a najskôr s ňou vstúpili do symbiózy a potom sa stali jej integrálnou súčasťou, organoidom.
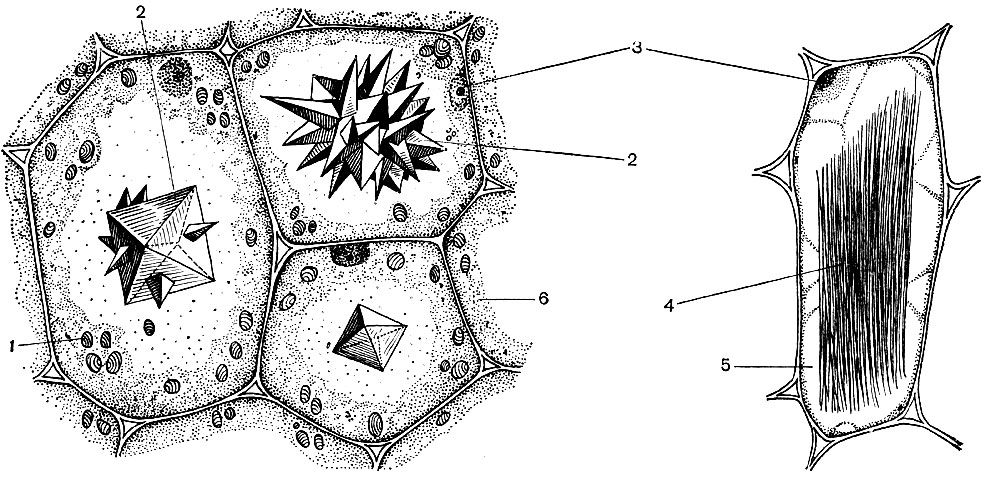
V nižších rastlinách sa fotosyntéza tiež vykonáva špecializovanými, aj keď nie tak vysoko vyvinutými ako membránové štruktúry v chloroplastoch. Vo fotosyntetických baktériách tvoria membrány obsahujúce chlorofyl sieť, ktorá prestupuje bakteriálnym telom. V modrozelených riasach sú fotosyntetické membrány zrastené do plochých bublín. V zelených a iných riasach je systém týchto membrán oddelený od zvyšku bunky krycou membránou a vytvára špeciálny organoid - chromatofor. Počet chromatoforov v bunke je malý; bunka často obsahuje iba jeden chromatofor. Ich tvar je u rias rôznych druhov veľmi odlišný.
V Spirogyre má chromatofor formu stuhy špirálovitej pozdĺž bunkových stien; v Clostridium sú to rebrované valce; zygnema má hviezdne telá.
Chromoplasty vznikajú buď z proplastidov, alebo z chloroplastov alebo z leukoplastov. Ich štruktúra vnútornej membrány je oveľa jednoduchšia ako štruktúra chloroplastov. Gran nie je, stróma obsahuje veľa žltého alebo oranžového pigmentu. Chromoplasty sa nachádzajú v bunkách okvetných lístkov, ovocia, okopanín.
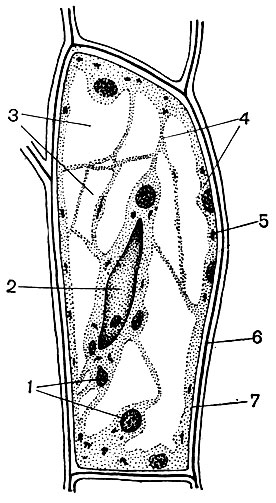
Typická rastlinná bunka má veľkú vakuolu naplnenú tekutým obsahom. Vakuola často zaberá takmer celý objem bunky, takže cytoplazma je len tenkou vrstvou susediacou s bunkovou membránou. Mladé bunky majú niekoľko malých vakuol, ktoré sa pri vývoji buniek zväčšujú a spájajú do jednej. Obsah vakuoly - bunkovej šťavy - je vodný roztok veľmi veľkého množstva látok: cukrov, aminokyselín, iných organických kyselín, pigmentov (farbív), vitamínov, trieslovín, alkaloidov, glykozidov, anorganických solí (dusičnany, fosfáty, chloridy) a niekedy aj bielkovín.
Všetky tieto látky sú odpadovými látkami bunky. Niektoré z nich sa skladujú vo vakuolárnej (bunkovej) šťave ako rezervné látky a časom sa dostanú späť do cytoplazmy na ďalšie použitie. Iné sú metabolické odpadové látky odstránené z cytoplazmy. Kyselina šťaveľová sa teda odstráni do vákua; kryštály šťavelanu vápenatého sa často ukladajú vo vákuovej šťave - niekedy vo forme monokryštálov, v iných prípadoch vo forme konglomerátu kryštálov tejto soli - mnohostranné (drúzy) alebo ihlicovité (raffidy) - znázornené na obrázku 34.
Prevažná väčšina rastlín (s výnimkou prokaryotických organizmov) má každá živá bunka jadro alebo niekoľko jadier. Bunka bez jadra môže žiť iba krátko. Anuklované bunky sitových skúmaviek sú živé bunky. Nežijú však dlho. Vo všetkých ostatných prípadoch sú nejadrové bunky mŕtve.

Jadro vždy leží v cytoplazme. Tvar jadra môže byť rôzny - guľatý, oválny, silne pretiahnutý, nepravidelne viaclaločný. V niektorých bunkách sa počas jeho fungovania menia obrysy jadra a na jeho povrchu sa vytvárajú laloky rôznych veľkostí. Veľkosti jadier nie sú rovnaké v bunkách rôznych rastlín a v rôznych bunkách tej istej rastliny. Pomerne veľké jadrá sa nachádzajú v mladých, meristematických bunkách, v ktorých môžu zaberať až 3/4 objemu celej bunky. Relatívna a niekedy absolútna veľkosť jadier vo vyvinutých bunkách je oveľa menšia ako u mladých.
Vonku je jadro pokryté škrupinou pozostávajúcou z dvoch membrán, medzi ktorými je medzera - perinukleárny priestor. Škrupina je prerušená pórmi. Vonkajšia z dvoch membrán membrány vydáva výrastky, ktoré priamo prechádzajú do stien endoplazmatického retikula cytoplazmy. Póry aj priame spojenie endoplazmatického retikula s perinukleárnym priestorom poskytujú úzky kontakt medzi jadrom a cytoplazmou.
Obsahom jadra je granulovaná základná látka (jadrová šťava alebo nukleoplazma), v ktorej sú umiestnené hustejšie štruktúry - chromozómy a jadierko. Nukleolus je prístroj na syntetizáciu materiálu ribozómov a miesta ich zhromažďovania z tohto materiálu.
Chromozómy sú tvorené z veľkého množstva molekúl deoxyribonukleovej kyseliny (DNA) kombinovaných s molekulami histónového proteínu. Molekuly DNA sú dlhé, zložito zabalené dvojreťazce. Každá molekula sa skladá z dvoch vlákien, ktoré sú špirálovito stočené jeden okolo druhého. Niť je zase reťazec obrovského množstva takzvaných nukleotidov. Nukleotid je zlúčenina dusíkatej bázy, uhľohydrátov (deoxyribóza) a kyseliny fosforečnej.
Každý z nukleotidov obsahuje jednu z nasledujúcich štyroch dusíkatých báz: adenín, guanín, cytozín alebo tymín. Podľa toho sa v DNA rozlišujú 4 rôzne nukleotidy: adenín (A), guanín (G), cytozín (C) a tymín (T). Nukleotidy sú navzájom spojené prostredníctvom svojich fosfátových skupín, čím vytvárajú dlhý reťazec. Dva reťazce, skrútené okolo seba a tvoriace jednu molekulu DNA, sú navzájom spojené chemickou interakciou (takzvané vodíkové väzby) dusíkatých báz ich nukleotidov. Bázy tvoria páry - jedna báza z jedného reťazca, druhá z druhého. Diagram na obrázku 35 poskytuje predstavu o poradí, v akom sú nukleotidy spojené do reťazca, a o dvoch reťazcoch navzájom.
Aj keď sú všetky molekuly deoxyribonukleovej kyseliny zostavené podľa opísaného jediného plánu, ich špecifické kvalitatívne zloženie je odlišné; líšia sa tiež veľkosťou molekúl. Molekula DNA obsahuje až 50 - 100 tisíc párov báz, ale počet ich párov v každej molekule sa líši od počtu párov nukleotidov v akejkoľvek inej molekule. Okrem toho existujú 4 rôzne typy nukleotidov a každá molekula DNA má vlastné inherentné percento nukleotidov každého daného typu. Inými slovami, kvantitatívny pomer A: G: C: T pre každú molekulu DNA je iný. A nakoniec, v každej molekule DNA je poradie striedania nukleotidov A, G, C, T charakteristické iba pre túto molekulu. Permutácia iba dvoch párov s 50 000 pármi báz, aj keď susedia, dramaticky mení vlastnosti celej molekuly; to isté, nahradenie najmenej jedného páru, povedzme od - G do A - T alebo G - Iz; to isté - absencia jedného páru z rovnakých 50 000. V skutočnosti sa molekuly DNA môžu navzájom líšiť nie v jednom spojení, ani v jednom alebo dvoch pároch nukleotidov, ale v obrovskom počte z nich naraz. Počet možných permutácií nukleotidových párov v molekule DNA je nekonečný a podľa toho je počet rôznych molekúl nekonečný, z ktorých každá má svoje vlastné vlastnosti. Sekvencia nukleotidov v molekule DNA je šifrovaný záznam o zložení konkrétneho proteínu, ktorý je charakteristický pre danú bunku. V každej molekule DNA sa postupne nachádzajú záznamy o zložení niekoľkých proteínov a vo všetkých molekulách DNA všetkých chromozómov jadra - záznamy o zložení všetkých proteínov bunky, ktorú je možné v nej syntetizovať počas jej života. Podstata týchto kódovaných záznamov je nasledovná.
Molekuly každého proteínu sú reťazce aminokyselín zapojených do série. Existuje asi 20 rôznych aminokyselín a podstata proteínu závisí od toho, z ktorých aminokyselín sa jeho molekula skladá, aké je ich celkové množstvo v molekule a v akom poradí sú navzájom spojené. Napríklad časť DNA zodpovedná za zloženie určitého proteínu (každá takáto časť sa nazýva gén) je záznam špecifickej sekvencie aminokyselín, ktoré tvoria molekulu daného proteínu, ich celkového počtu v ňom. Každé 3 po sebe idúce nukleotidy reťazca DNA označujú (kódujú) jednu aminokyselinu zodpovedajúcej molekuly proteínu. Nasledujúce 3 nukleotidy kódujú ďalšiu aminokyselinu atď. Počet možných odlišných kombinácií 3 nukleotidov spomedzi štyroch rôznych typov je viac ako dosť na kódovanie 20 aminokyselín. Je zaujímavé, že rovnaké aminokyseliny v celom živom svete sú kódované rovnakými kombináciami nukleotidov. Výsledkom je, že v jednom géne jeho nukleotidové zloženie kóduje aminokyselinové zloženie všetkých proteínov, ktoré je možné syntetizovať v danej bunke.
Bunkové proteíny sú enzýmy. Enzýmy určujú priebeh všetkých reakcií, ktoré tvoria podstatu vitálnej činnosti buniek. Tvorba a transformácia všetkých ostatných látok v bunke závisí od prítomnosti určitých enzýmov, či už ide o tuky, uhľohydráty, alkaloidy, živice atď. To zase vytvára všetky vlastnosti bunky, ktoré ju odlišujú od buniek iných rastlín v bunkových jadrách ktorý kódoval zloženie ďalšej sady enzýmov. Preto sú informácie o zložení všetkých proteínov, ktoré sa dajú v bunke vytvoriť, informáciou o všetkých vlastnostiach bunky a organizmu. Ďalej uvidíme, že táto informácia je dedičná, to znamená, že je úplne prenášaná z bunky do bunky počas ich reprodukcie a z materskej rastliny na dcérske rastliny.
Táto informácia je uložená v chromozómoch. Uskutočnenie týchto informácií - syntéza bielkovín - však v nich neprebieha. Gény chromozómov pôsobia iba ako iniciátory tejto syntézy. Keď bunka potrebuje tvorbu konkrétneho proteínu, aktivuje sa gén, v ktorom je zloženie tohto proteínu šifrované. To znamená, že na časti reťazca DNA, ktorý tvorí tento gén, sa tvoria molekuly takzvanej informačnej ribonukleovej kyseliny (i-RNA).
Štruktúra a zloženie týchto molekúl, ktoré sú jednotlivými reťazcami nukleotidov, odráža nukleotidovú štruktúru génu, na ktorom boli vytvorené. Takto sa kopírujú informácie o zložení budúceho proteínu.
Tvorba molekúl m-RNA (veľa sa ich tvorí na každom géne) tiež znamená množenie informácií, akoby to bolo, potlač mnohých identických matíc, ktoré odrážajú štruktúru rovnakého génu, a tým nesú informácie o aminokyselinovom zložení daného proteínu v nukleotidovej sekvencii ich molekúl. Tieto génové výtlačky prechádzajú z jadra do cytoplazmy. Tu sa dešifrujú informácie obsiahnuté v matriciach - molekulách i-RNA, informácie sa realizujú, preložia sa z jazyka nukleotidovej sekvencie do jazyka aminokyselinovej sekvencie, teda syntézy proteínových molekúl daného zloženia.
Na dekódovaní a syntéze je okrem molekuly i-RNA zapojené aj veľké množstvo molekúl rôznych transportných ribonukleových kyselín (t-RNA), ribozómov a množstvo enzýmov. Aminokyseliny sa viažu na t-RNA - molekulu po molekule. Každá z 20 druhov aminokyselín má svoju vlastnú t-RNA. Molekula t-RNA má teda chemické skupiny, ktoré dokážu rozpoznať svoju aminokyselinu, a to tak, že si ju vyberie zo všetkých dostupných aminokyselín. To sa deje pomocou špeciálnych enzýmov. Po rozpoznaní svojej aminokyseliny s ňou t-RNA nadväzuje spojenie. Ribozóm je pripojený k začiatku molekuly i-RNA, ktorá sa pohybom pozdĺž i-RNA spája navzájom v polypeptidovom reťazci presne s tými aminokyselinami, ktorých poradie je šifrované nukleotidovou sekvenciou danej i-RNA. K tomuto čítaniu informácií, k ich dekódovaniu dochádza vďaka špeciálnemu mechanizmu dostupnému v transportnej RNA, ktorého molekuly tvoria presne tie aminokyseliny, ktoré sú „označené“, „napísané“ nukleotidovou sekvenciou molekuly i-RNA. Ribozóm je prístroj na chemickú väzbu aminokyselín na molekulu proteínu.
Všeobecná schéma je teda nasledovná. Na aktivovanom géne - časti jedného z reťazcov molekuly DNA - sa syntetizujú nukleotidové reťazce, molekuly i-RNA, ktorých zloženie presne odráža zloženie génu, a preto nesie záznam o zložení proteínu kódovaného týmto génom. V cytoplazme ribozómu sa na základe informácií prenášaných z génu molekulami m-RNA pomocou transportných RNA kombinujú rôzne aminokyseliny v danej sekvencii, čo vedie k vytvoreniu molekuly proteínu, ktorého zloženie je kódované v tomto géne.
Tvorba molekúl i-RNA na aktivovanom géne slúži ako príkaz aj ako špecifický program pre syntézu prísne definovaného proteínu. Rovnaká molekula mRNA sa používa ako plán na vytvorenie mnohých rovnakých molekúl proteínu. Je to však dosť krátkodobé, preto je na dlhodobú syntézu nových molekúl rovnakého proteínu nevyhnutná tvorba identických, ale nových kópií molekúl m-RNA na rovnakom géne. S prechodom génu do neaktívneho stavu - a to sa stane, keď bunková potreba daného proteínu zmizne - je blokovaný, prestáva tvoriť i-RNA a čoskoro sa zastaví syntéza tohto proteínu. Počas života bunky potrebuje rôzne bielkoviny. Zakaždým sa aktivujú gény, ktoré ich definujú, a vytvorí sa zodpovedajúce zloženie i-RNA molekúl.
Každý ribozóm počas života produkuje veľa molekúl rôznych proteínov. Môže fungovať na základe akejkoľvek i-RNA a výsledok, povaha proteínu, ktorý vytvorila, závisí iba od zloženia i-RNA v kontakte, s ktorým ribozóm tentoraz pracoval.
Bunkové jadro teda vykonáva nasledujúce vzájomne súvisiace funkcie. Uchováva informácie o zložení všetkých proteínov, ktoré je možné syntetizovať v danej bunke počas jej života. (Výnimkou sú niektoré proteíny mitochondrií a chloroplastov. Ich zloženie je zakódované vo vlastnej DNA týchto organel, kde sa tieto DNA nachádzajú. Tu na mieste prebiehajú všetky stupne dekódovania informácií obsiahnutých v týchto DNA, vrátane syntézy týchto proteínov pomocou vlastné ribozómy.)
Jadro uchováva informácie o všetkých vlastnostiach bunky a organizmu. Jadro organizuje syntézu každého z týchto proteínov v správny čas. Počas bunkového delenia, sprevádzaného nukleárnym delením, sa všetky tieto informácie prenášajú v plnom rozsahu do každého z novovzniknutých jadier, do každej novej bunky. To je možné vďaka skutočnosti, že pred rozdelením sa všetok génový materiál chromozómov zdvojnásobuje, vytvárajú sa jeho dve identické kópie a jedna z nich končí v každom z nových jadier. Jadro každej bunky obsahuje kompletnú sadu génov charakteristických pre daný organizmus. Nie všetky gény však fungujú počas života rôznych špecializovaných buniek. V niektorých bunkách funguje jedna časť génov, v iných - iná, v tretej - tretia. Preto sa bunky rôznych tkanív jedného organizmu navzájom líšia. Významná časť génov zostáva v pasívnom stave, ktorý „ukladá informácie“ iba od narodenia po smrť bunky alebo organizmu. Gény v danej bunke navyše nepracujú súčasne: niektoré sú aktívne v jednom období života bunky, v určitej fáze jej vývoja; iné - na druhej atď.
Prečo niektoré gény zostávajú neaktívne, ako sa iné zapínajú a vypínajú, je veľmi zložitý a dôležitý problém, ktorý sa teraz intenzívne skúma. Zdá sa, že proteíny-históny hrajú dôležitú úlohu pri blokovaní a odblokovaní génov. Sú súčasťou chromozómov a sú spojené s DNA. K aktivácii, „oslobodeniu“ génu pravdepodobne dôjde, keď sa molekula histónu oddelí od zodpovedajúcej časti DNA, čím sa odhalia jeho reťazce, ktoré im umožnia rozmotať sa a začať chemicky fungovať. Pripojenie histónu vedie k zablokovaniu génu. Ako je však kontrolované viazanie a oddeľovanie histónov, nie je dosť jasné.
Génový materiál, ako už bolo povedané, sa nachádza v chromozómoch. Preto je práca jadra na ukladanie dedičných informácií, ich duplikovanie a prenos z bunky do bunky, organizovanie syntézy rôznych proteínov počas života bunky, predovšetkým prácou chromozómov. Aj keď existujú po celý život bunky, v jadre sa rozlišujú vo forme jasných štruktúr iba počas bunkového delenia. V tomto okamihu možno kondenzáciu materiálu chromozómov a chromozómu zistiť pomocou metód mikroskopie ako jednotlivých, jasne definovaných útvarov.
Chromozómy majú rôzne tvary. Jedná sa buď o rovné alebo zakrivené palice, alebo o rôzne háčiky, oválne telá, gule (obr. 36). Veľmi sa líšia veľkosťou. Každá rastlinná bunka daného druhu obsahuje vo svojom jadre rovnakú sadu (alebo sady) chromozómov z prísne definovaného počtu rôznych, ale prísne definovaných chromozómov. U všetkých vyšších a niektorých nižších rastlín sa počas ich životného cyklu striedajú dve generácie: s bunkami obsahujúcimi v ich jadrách jednu (haploidnú, n) sadu chromozómov a s bunkami, ktorých jadrá majú dvojitú (diploidnú, 2 n) sadu chromozómov. Haploidná sada pozostáva z n rôznych chromozómov, jedného z každého typu. Číselná hodnota je pre všetky haploidné bunky všetkých rastlín daného druhu prísne konštantná. Takže pre lesné jahody je toto číslo 7, pre hrášok - tiež 7, ale pre fazuľu - 11, pre jabloň - 17, pre hlinenú hrušku - 51, pre cukrovú trstinu - 60. Diploidná súprava sú dve haploidné dohromady. Obsahuje dva chromozómy každého typu. Ak má hrach v haploidnej sade 7 rôznych chromozómov, potom má diploidná sada 7 rôznych párov chromozómov a dva chromozómy v každom páre sú rovnaké. Je ľahké vypočítať, že pre uvedené rastliny je diploidná sada (2n) pre jahody 14, pre hrášok - 14, pre fazuľa - 22, pre jabloň - 34, pre hlinenú hrušku - 102 a pre cukrovú trstinu - 120 chromozómov.
V životnom cykle machov a papradí sa organizmy striedajú s diploidnými a haploidnými bunkami. V papradiach vedú tieto organizmy samostatný, izolovaný život. V machoch žije diploidná rastlina na haploidnej rastline. V diploidnom organizme sa vytvárajú haploidné bunky - spóry. Každý 1 z nich, klíčiaci, dáva haploidný organizmus, to znamená organizmus zostavený z haploidných buniek. V ňom sa tvoria haploidné pohlavné bunky - gaméty.
V okamihu oplodnenia sa mužské a ženské gaméty zlúčia a vytvorí sa jedna bunka - zygota, ktorá má jedno jadro - výsledok fúzie jadier oboch gamét. Z každej z gamét dostane jadro zygoty haploidnú sadu chromozómov a vo výsledku ich má dvojitú, diploidnú. Organizmus sa vyvíja zo zygoty, ktorej každá bunka má diploidnú sadu chromozómov.

V diploidnej sade sú dva chromozómy každého páru identické v tvare, vnútornej štruktúre a obsahujú gény, ktoré riadia vzhľad homogénnych znakov (obr. 36). Nazývajú sa homológne chromozómy. Jeden z nich pochádza z haploidnej sady otcovskej gamety, druhý z materskej. Preto v dvojdomých organizmoch jeden z nich nesie gény, ktoré určujú vývoj charakteristík v jeho jurisdikcii podľa otcovského typu, druhý - podľa materského typu.
Homologické chromozómy iného páru rovnakým spôsobom určujú vývoj ďalšej série znakov, tretieho páru - tretieho radu atď. Aj keď otcovské a materské organizmy patria k rovnakému druhu, ich dedičné vlastnosti nie sú totožné - majú tiež individuálne rozdiely. Niektoré gény jedného homológneho chromozómu preto nie sú identické so zodpovedajúcimi génmi druhého.
Haploidná súprava, ktorá je súčasťou diploidu a pochádza z otcovskej pohlavnej bunky, nesie so svojimi individuálnymi vlastnosťami otcovskú dedičnosť a haploidná súprava z materskej pohlavnej bunky nesie materskú. Komplexná interakcia homogénnych, ale nie vždy identických génov dvoch haploidných súborov, ktoré spolu tvoria jeden diploid, určuje, aké vlastnosti sa objavia u diploidného potomka, ktorý je v podstate hybridom otca a matky.
U gymnospermov a krytosemenných rastlín sa v zásade vyskytuje striedanie generácií, ako u machov a papradí, ale ich haploidná fáza je výrazne znížená a často ju predstavuje iba skupina buniek. Nežije samostatne, ale v tele haploidnej rastliny. V krytosemenných rastlinách je ženská haploidná generácia uzavretá v embryonálnom vaku umiestnenom vo vajíčku a mužská generácia je vo vnútri peľového zrna.
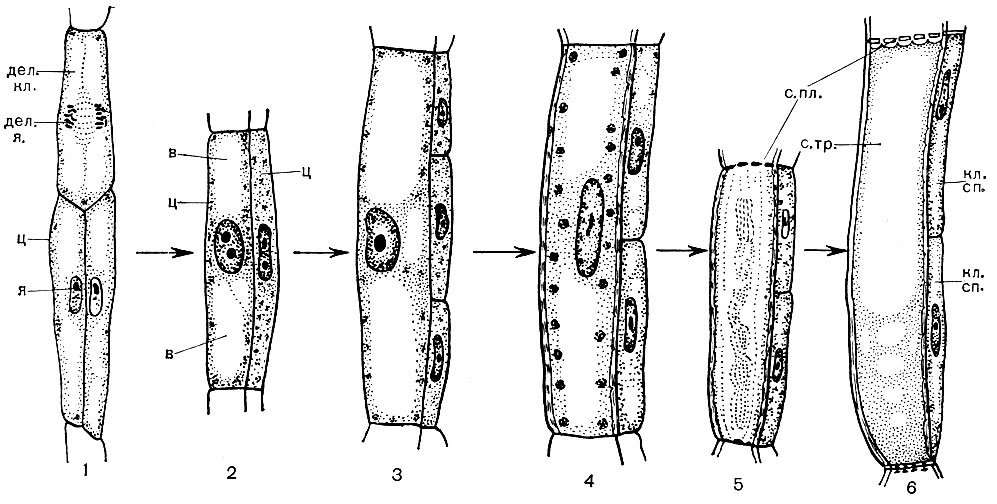
Už pred bunkovým delením si každá molekula DNA v každom chromozóme pripája svoju kópiu - druhú z tej istej molekuly. Vďaka tomu sa zdvojnásobí celý dedičný materiál bunky a každý chromozóm sa teraz skladá z dvoch rovnakých častí - chromatidov. Ďalej je „úlohou“ bunky rozdeliť každý chromozóm na chromatidy a striktne ich rovnomerne rozdeliť medzi budúce dcérske bunky: do každého z nich musí byť zaslaný jeden chromatid z každého chromozómu. Toto sa deje nasledovne. Bezprostredne pred bunkovým delením sú chromozómy silne kondenzované a stiahnuté. Potom sú umiestnené v jednej rovine - pozdĺž rovníka jadra, pričom jeden chromatid každého z nich smeruje k jednému pólu bunky, druhý - k opačnému. Jadrová obálka zmizne a jadierko sa tiež rozpustí. Medzi pólmi bunky sa objavujú vlákna, ktoré spolu tvoria vretenovitý tvar. Vlákna sa zhromažďujú z navzájom spojených mikrotubulov. Vreteno sa skladá z dvoch druhov závitov. Niektoré sú spojité bunky prechádzajúce od jedného pólu k druhému. Ďalšie ťahajú, pričom každá z nich spája pól s jednou z chromatidov. V chromozóme je miesto - kinetochór, ku ktorému sú pripevnené sťahovacie nite - jedno z jedného pólu, druhé z druhého. Ďalej sa chromatidy rozchádzajú. Chromatidy, ktoré predtým tvorili každý chromozóm, sú navzájom oddelené a vytiahnuté vláknami sa rozchádzajú k opačným pólom bunky. Výsledkom je, že na každom póle sa zhromaždí jedna úplná sada chromatidov, ktoré sú teraz chromozómami. Potom sa vlákna vretena rozpadnú, okolo každej sady chromozómov sa vytvorí jadrový obal, chromozómy sa veľmi nafúknu (despirálujú). V každom jadre sa objaví jadro. Jadro získava štruktúru charakteristickú pre jadrá nedeliacich sa buniek. V strednej rovine bunky sa vytvorí septum, ktoré bunku rozdelí na dve dcérske bunky.
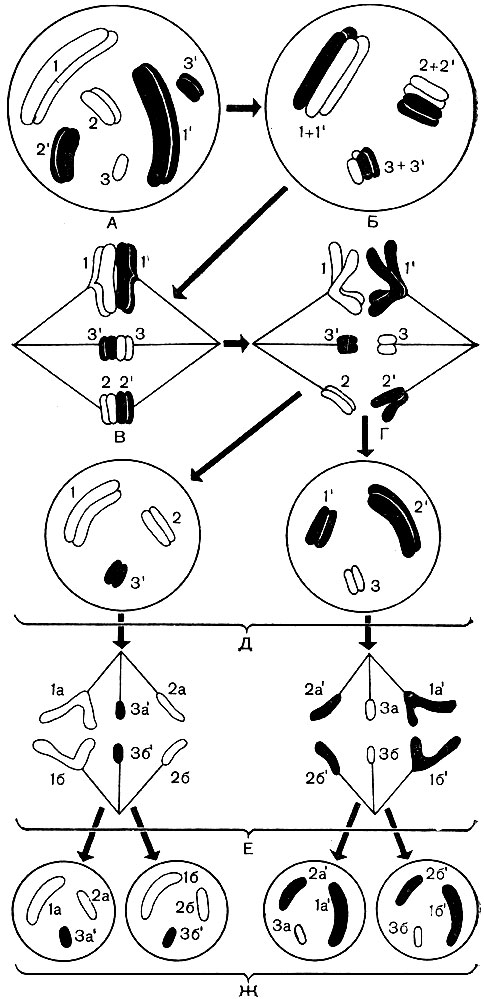
Obrázok: 38. Schéma správania chromozómov „podmienenej“ bunky počas meiózy. A - zobrazuje diploidné bunkové jadro obsahujúce 3 páry chromozómov (1 a 1 "- 1. pár homológnych chromozómov, 2 a 2" - druhý pár, 3 a 3 "- 3. pár). 2n \u003d 6,3 chromozómy pochádzajúce z materského organizmu sú ľahké (počet bez mozgovej príhody); ich spárované homológne chromozómy pochádzajúce z otcovského organizmu sú začiernené (počty mozgových príhod). Homologické chromozómy sú označené spoločným počtom. Každý chromozóm pozostáva z dvoch chromatidov. B - fúzia homológnych chromozómov V tomto štádiu si homológne chromozómy vymieňajú navzájom samostatné sekcie. Výsledkom je určité prerozdelenie dedičného materiálu medzi matkami a otcami medzi chromozómami (kríženie). C - vytvárajú sa vlákna vretena, ktoré sa pripájajú k chromozómom, jadrová membrána zmizne. D - homológne chromozómy sa rozchádzajú k opačným pólom. bunky, na póloch je z každej dvojice jeden homológny chromozóm, celkový počet chromozómov na každom póle je polovičný ako v pôvodnom jadre A. D - obrázok existujú dve jadrá s haploidnou sadou chromozómov v každom z nich; jeden z nich dostal viac babičkiných (svetlých, materských vo vzťahu k jadru A) a menej dedových (začiernených, otcovských vo vzťahu k jadru A) chromozómov, v druhom - naopak; nové jadrá teda nie sú navzájom úplne identické v zložení svojej dedičnej látky; ich rozdiel je spôsobený aj krížením, ktoré sa vyskytuje u chromozómov v štádiu B. E - G - mitotické rozdelenie každého z haploidných jadier D: pozdĺžne štiepenie každého chromozómu, divergencia chromozómov k pólom, tvorba dvoch haploidných jadier z každého jadra D. Výsledkom boli 4 bunky s haploidnými jadrami namiesto jednej bunky s diploidnou sadou chromozómov
Organely nie sú distribuované striktne rovnomerne medzi dcérske bunky, ale potom sa ich základné časti syntetizujú v bunkách, zhromaždia sa nové kópie organel každého druhu a ich počet v každej bunke sa obnoví na normálny stav. Samotné bunky rastú. V chromozómoch sa dedičný materiál duplikuje, potom chromozómy pozostávajú, rovnako ako pred bunkovým delením, z dvoch chromatidov. Bunka je pripravená na nové rozdelenie.
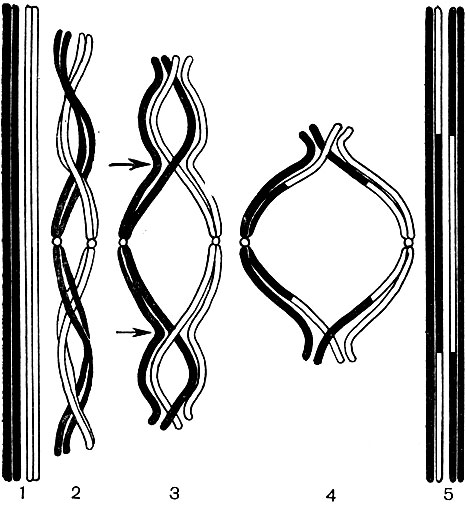
Obrázok: 38A. Crossoverová schéma počas meiózy v štádiu B na obr. 38. 1 - dva homológne chromozómy, z ktorých každý pozostáva z dvoch sesterských chromatidov; 2 - homologické chromozómy sú navzájom prepletené; 3 - každý chromozóm je rozdelený na svoje základné chromatidy; chromozómy sa začnú navzájom odpudzovať a rozchádzajú sa, ale v niektorých bodoch (na obrázku - na štyroch) zostávajú spojené; sesterské chromatidy sú zdvojnásobené v centrálnej slučke a nesesterské chromatidy v obidvoch krajných slučkách; body, kde je možný prechod, sú označené šípkami; 4 - chromatidy si vymenili oblasti a chromozómy, ktoré prešli akousi vzájomnou hybridizáciou, sa rozchádzajú; 5 - dva odlišné, ale už „hybridné“ (na rozdiel od tých, ktoré sú uvedené na 1) chromozóme
Keď je diploidná generácia buniek nahradená haploidnou, nastáva takzvané redukčné jadrové delenie, meióza. Počas meiózy (obr. 38) sa homologické chromozómy každého páru priblížia k sebe, navzájom po svojej dĺžke tesne susedia a krútia sa. Medzi susednými homológnymi chromozómami sa vymieňajú samostatné časti. Vďaka tomu sa niektoré z génov otcovských chromozómov prevedú do zloženia materských chromozómov a zodpovedajúce gény materských chromozómov obsadzujú uvoľnené miesta v otcovských chromozómoch (fenomén kríženia - obr. 38A). Vzhľad tieto a ďalšie chromozómy sa vo výsledku nemenia, ale ich kvalitatívne zloženie sa líši. Dedičstvo po otcovi a matke sa prerozdeľuje a zmieša. Ďalej sa jadrový obal a jadro rozpustia, vytvorí sa vretenový aparát, rovnaký ako počas mitózy. Homologické chromozómy sú oddelené a pomocou vretenových nití sa rozchádzajú k pólom bunky. Jeden pól má jednu haploidnú sadu chromozómov (jeden homológny chromozóm z každého páru), druhý má druhú haploidnú sadu.
Po meióze sa vytvorené haploidné jadrá delia mitózou. V tomto prípade je každý chromozóm haploidnej sady rozdelený na dve chromatidy, rozchádzajú sa a vznikajú dcérske haploidné bunky.
Meióza sa líši od mitózy dvoma zásadnými spôsobmi. Najskôr sa počas meiózy zhlukuje každý pár homológnych chromozómov a vymieňa sa medzi nimi časť dedičného materiálu, čo počas mitózy neplatí. Po druhé, počas meiózy sa homológne chromozómy rozchádzajú k pólom bunky, jeden z každého páru, a počas mitózy sa každý chromozóm štiepi na chromatidy a chromatidy sa rozchádzajú k pólom, jeden z každého chromozómu. V dôsledku meiózy sa z diploidnej bunky vytvárajú haploidné bunky.
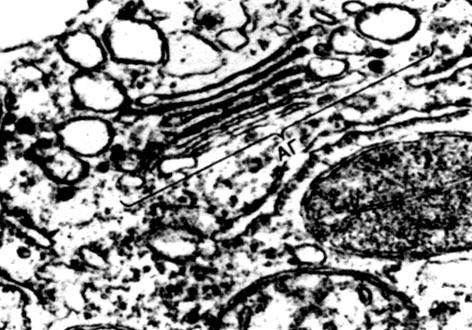
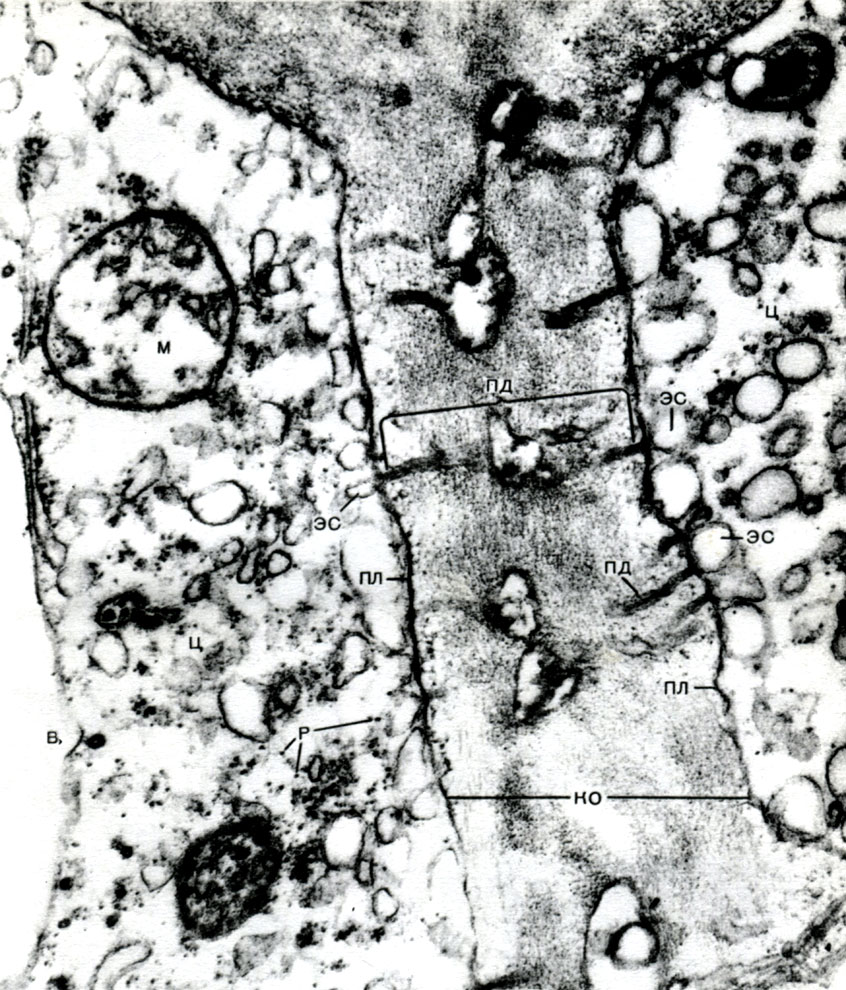
Tabuľka 10. Štruktúra rastlinných buniek. Golgiho aparát v cytoplazme vyvíjajúcich sa koreňových vlasov reďkovky (Raphanus sativus). Elektrónový mikrofotografia (zväčšenie x 52000) od M.F. Danilova: ag - Golgiho prístroj, časti plochých vakov a bubliny sú viditeľné
V bunkách niektorých tkanív počas ich vývoja dochádza k neúplnej mitóze: chromozómový materiál v jadrách sa zdvojnásobí, chromozómy sa rozdelia na polovicu, ale namiesto toho, aby vytvorili dve jadrá, zostanú v pôvodnom jadre. Od tejto chvíle neobsahuje diploidnú, ale tetraploidnú (štvornásobnú) sadu chromozómov. Proces vedúci k tejto duplikácii chromozómov v jednom jadre sa nazýva endomitóza - vnútorná mitóza. Ak sa to stane jednému jadru dvakrát, stane sa z neho osemnásobná (osemnásobná) sada atď. Bunky, ktorých jadrá nesú viac ako dve sady chromozómov, sa nazývajú polyploidné, to znamená multiploidné. Bunková polyploidia v niektorých prípadoch zvyšuje ich životaschopnosť, pretože každý gén je duplikovaný niekoľkými ďalšími rovnakými génmi. Gény jednej kvality pôsobia jednotne a poškodenie žiadneho z nich nevedie k strate znaku, ktorý určuje, pretože je kompenzované prácou zvyšných homogénnych génov. V mnohých prípadoch sú polyploidné bunky väčšie a obsahovo bohatšie ako diploidné bunky. Boli vyšľachtené odrody polyploidných kultúrnych rastlín, ktoré zvýšili ekonomické vlastnosti.
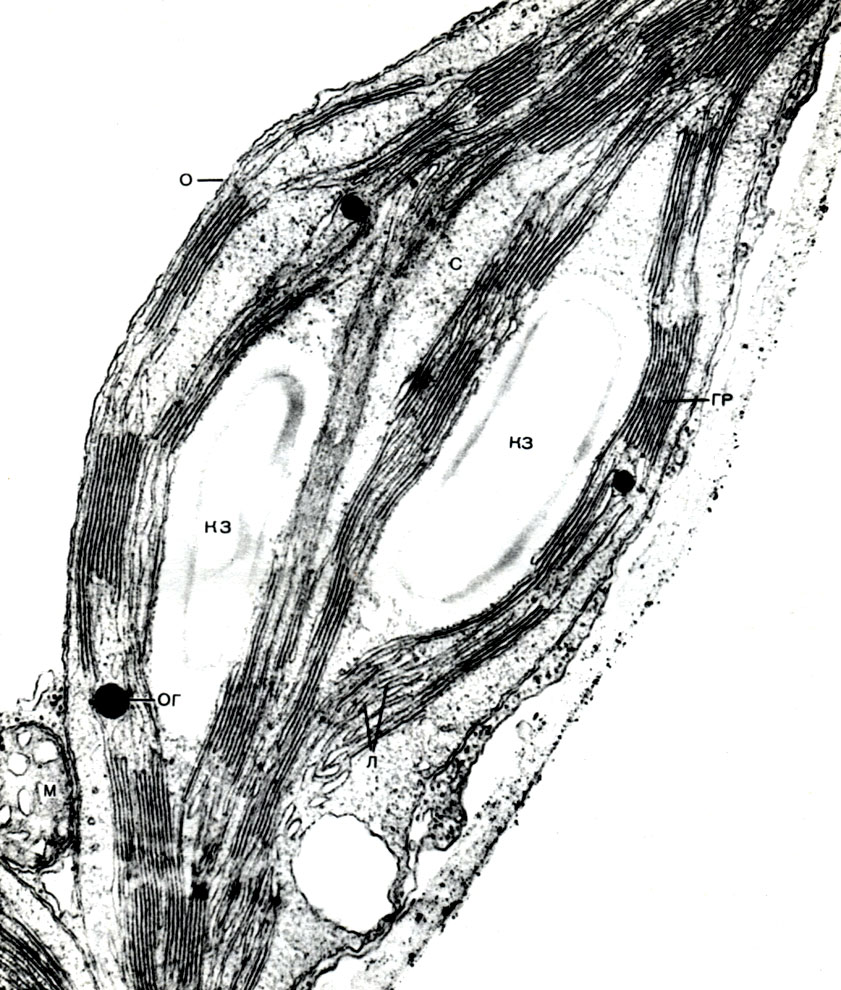
Veda, ktorá skúma bunku, sa nazýva cytológia. Bunka je základom štruktúry všetkého živého a všetkých životných procesov; preto je možné najdôležitejšie všeobecné biologické problémy vyriešiť cytológiou a metódami jej výskumu.
Pomocou cytológie sa rieši veľa prakticky dôležitých otázok: boj proti chorobám rastlín (hubovým, bakteriálnym), šľachtenie nových odrôd kultúrnych rastlín, prekonávanie sterility (neplodnosti) hybridných odrôd.
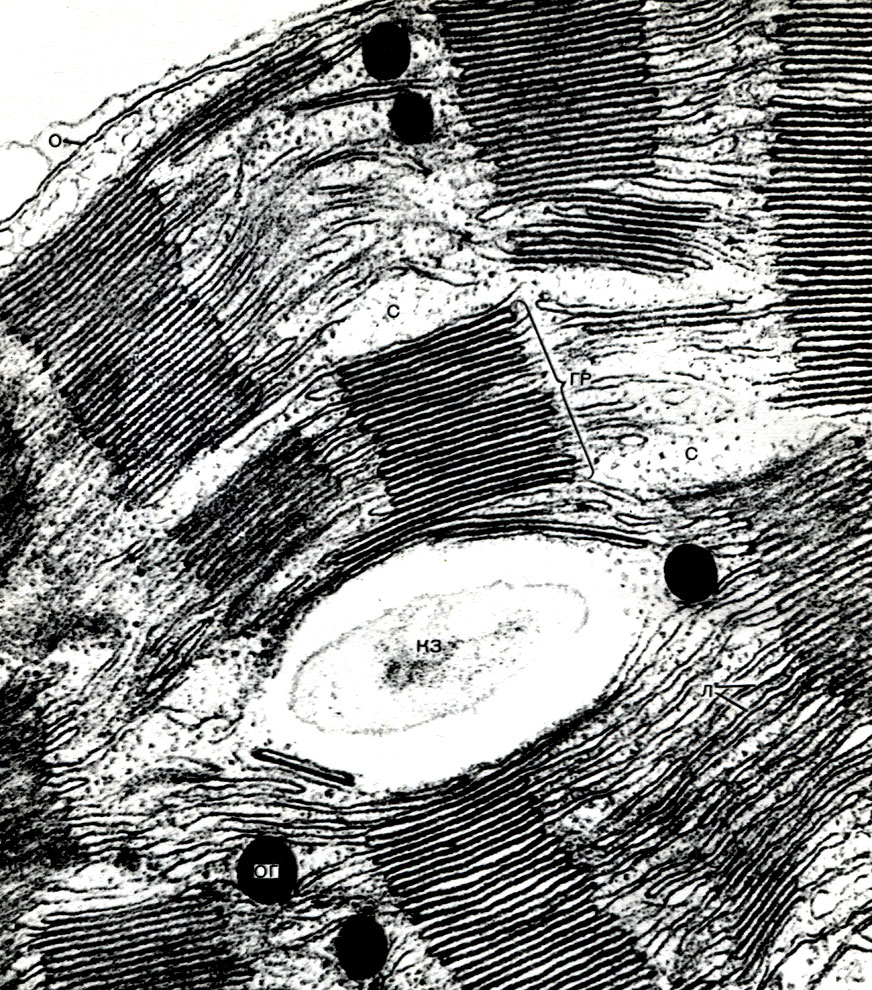
Moderná cytológia skúma tieto problémy pomocou rôznych metód - mikroskopických, biochemických, biofyzikálnych, genetických - a úzko interagujúcich s inými príbuznými biologickými vedami.