Viruskrankheiten traten in der Antike auf, aber die Virologie als Wissenschaft begann sich Ende des 19. Jahrhunderts zu entwickeln.
1892 stellte der russische Wissenschaftler-Botaniker D. I. Ivanovsky, der die Mosaikkrankheit von Tabakblättern untersuchte, fest, dass diese Krankheit durch die kleinsten Mikroorganismen verursacht wird, die durch feinporige Bakterienfilter gelangen. Diese Mikroorganismen werden als filtrierbare Viren (vom lateinischen Virus - Gift) bezeichnet. Später wurde gezeigt, dass andere Mikroorganismen durch Bakterienfilter gelangen. Daher wurden die gefilterten Viren einfach als Viren bezeichnet.
Im Allgemeinen sagen die Form des Virions und das Vorhandensein oder Fehlen einer Hülle wenig darüber aus, welche Krankheiten Viren verursachen können oder welche Arten sie infizieren können, sind aber dennoch ein nützlicher Weg, um mit der Klassifizierung von Viren zu beginnen. Abbildung 5 Viren können eine komplexe Form haben oder relativ einfach sein.
Welche der folgenden Aussagen zur Struktur des Virus ist richtig? Der Kern des Virus enthält das Genom oder den allgemeinen genetischen Inhalt des Virus. Virale Genome sind im Vergleich zu Bakterien oder Eukaryoten, die nur Gene enthalten, die für Proteine \u200b\u200bkodieren, die das Virus nicht aus der Wirtszelle erhalten kann, typischerweise klein. Dieses genetische Material kann einzelsträngig oder doppelsträngig sein. Es kann auch linear oder kreisförmig sein. Während die meisten Viren ein Segment Nukleinsäure enthalten, haben andere Genome, die aus mehreren Segmenten bestehen.
Einen großen Beitrag zur Erforschung von Viren leisteten sowjetische Virologen: M. A. Morozov, N. F. Gamaleya, L. A. Zilber, M. P. Chumakov, A. A. Smorodintsev, V. M. Zhdanov und andere.
Viren sind nichtzelluläre Form Existenz lebender Materie. Sie sind sehr klein. Nach dem bildlichen Ausdruck von VM Zhdanov "kann ihre Größe im Verhältnis zur Größe der durchschnittlichen Bakterien mit der Größe einer Maus im Verhältnis zum Elefanten verglichen werden." Viren konnten erst nach der Erfindung des Elektronenmikroskops erkannt werden.
Viren können als obligatorisch angesehen werden intrazelluläre Parasiten... Das Virus muss sich an eine lebende Zelle anlagern, sie aufnehmen, ihre Proteine \u200b\u200bproduzieren und ihr Genom kopieren und einen Weg finden, der Zelle zu entkommen, damit das Virus andere Zellen und letztendlich andere Menschen infizieren kann. Viren können nur bestimmte Arten von Wirten und nur bestimmte Zellen in diesem Wirt infizieren. Die molekulare Basis für diese Spezifität ist, dass ein spezifisches Oberflächenmolekül, das als viraler Rezeptor bekannt ist, auf der Oberfläche der Wirtszelle gefunden werden muss, damit sich das Virus anlagern kann.
Gegenwärtig werden viele Methoden zur Untersuchung von Viren verwendet: chemische, physikalische, molekularbiologische, immunbiologische und genetische.
Alle Viren sind in Infektionen von Menschen, Tieren, Insekten, Bakterien und Pflanzen unterteilt.
Viren haben eine Vielzahl von Formen und biologischen Eigenschaften, aber alle haben gemeinsame strukturelle Merkmale. Reife Viruspartikel werden Virionen genannt.
Darüber hinaus sind metabolische Unterschiede, die bei verschiedenen Zelltypen aufgrund der unterschiedlichen Genexpression beobachtet werden, ein wahrscheinlicher Faktor, bei dem Zellen das Virus zur Replikation verwenden können. Die Zelle muss die vom Virus benötigten Substanzen enthalten, wie z. B. Enzyme, in denen das virale Genom selbst keine Gene enthält oder das Virus sich mit dieser Zelle nicht replizieren kann.
Der Virus muss die Zelle zur Replikation "übernehmen". Der virale Replikationszyklus kann zu dramatischen biochemischen und strukturellen Veränderungen in der Wirtszelle führen, die zu Zellschäden führen können. Diese Veränderungen, die als zytopathische Wirkungen bezeichnet werden, können die Zellfunktion verändern oder sogar die Zelle zerstören. Einige infizierte Zellen, wie die mit dem Erkältungsvirus infizierten, sterben durch Lyse oder Apoptose ab und setzen gleichzeitig alle Virionen der Nachkommen frei. Symptome viruserkrankungen sind das Ergebnis einer Immunantwort auf ein Virus, das versucht, das Virus aus dem Körper zu kontrollieren und zu eliminieren, sowie auf durch das Virus verursachte Zellschäden.
Im Gegensatz zu anderen Mikroorganismen, die sowohl DNA als auch RNA enthalten, enthält das Virion nur eine der Nukleinsäuren - entweder DNA oder RNA.
Die Nukleinsäure von Viren kann einzelsträngig und doppelsträngig sein. Fast alle Viren, die RNA enthalten, haben einzelsträngige RNA in ihrem Genom, und diejenigen, die DNA enthalten, haben doppelsträngige DNA. In Übereinstimmung mit zwei Arten von genetischen Substanzen werden Viren in RNA- und DNA-haltige Viren unterteilt. DNA-haltige Familien umfassen 5 Familien, RNA-haltige - 10 Familien.
Während des Knospungsprozesses wird die Zelle nicht sofort lysiert oder getötet. Die meisten produktiven Virusinfektionen folgen ähnlichen Schritten im Virusreplikationszyklus: Verschachteln, Infiltrieren, Entfernen der Beschichtung, Replikation, Zusammenbau und Freisetzung. Das Virus bindet an eine spezifische Rezeptorstelle auf der Membran der Wirtszelle durch Bindungsproteine \u200b\u200bim Kapsid oder in seine Hülle eingebettete Proteine. Die Bindung ist spezifisch und normalerweise bindet das Virus nur an Zellen einer oder mehrerer Arten und nur an bestimmte Zelltypen innerhalb dieser Arten mit den entsprechenden Rezeptoren.
* (Hier finden Sie nur Daten zu einigen für den Menschen pathogenen Viren.)
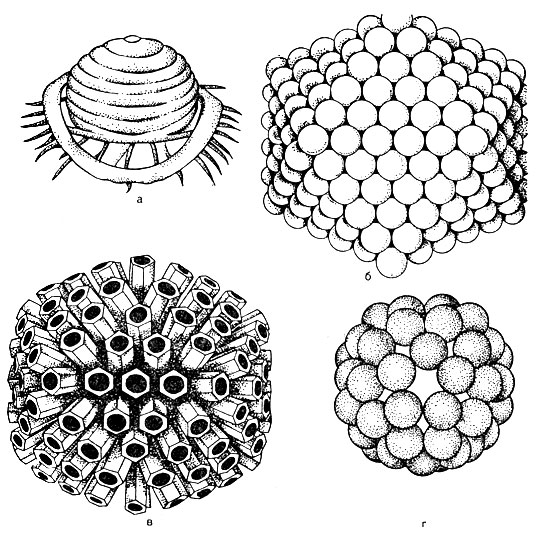
Virion-Struktur... Im Zentrum des Virions befindet sich eine Nukleinsäure, die von einem Kapsid (aus der griechischen Kanca-Box) umgeben ist. Das Kapsid besteht aus Proteinuntereinheiten, die als Kapsomere bezeichnet werden. Ein reifes Virus ist ein Nukleokapsid in chemischer Struktur. Die Anzahl der Kapsomere und die Art und Weise, wie sie verpackt sind (Abb. 52), sind für jeden Virustyp streng konstant. Beispielsweise enthält das Poliomyelitis-Virus 32 Kapsomere, während das Adenovirus 252 Kapsomere enthält. Kapsomere können in Form eines Polyeders mit gleichmäßigen symmetrischen Kanten gestapelt werden - quaderförmige Form (z. B. Adenovirus). Das spiralförmige (sphärische) Styling ist charakteristisch für Influenzaviren. Es kann eine Art von Symmetrie geben, bei der die Nukleinsäure die Form einer Feder hat, um die Kapsomere gelegt werden. In diesem Fall hat das Virus eine stabförmige Form - ein Virus, das eine Tabakblattkrankheit verursacht.
Im Gegensatz zu tierischen Viren wird die Bakteriophagen-Nukleinsäure nackt in die Wirtszelle eingeführt, wobei das Kapsid außerhalb der Zelle verbleibt. Pflanzen- und Tierviren können durch Endozytose in Zellen eindringen, wobei die Zellmembran das gesamte Virus umgibt und verschlingt. Einige umhüllte Viren gelangen in die Zelle, wenn die Virushülle direkt mit der Zellmembran verschmilzt. Das virale Kapsid zersetzt sich dann in der Zelle und die virale Nukleinsäure wird freigesetzt, die dann für die Replikation und Transkription verfügbar wird.
Der Replikationsmechanismus hängt vom viralen Genom ab. Es gibt natürlich Ausnahmen von diesem Muster. Wenn die Wirtszelle nicht die für die Virusreplikation erforderlichen Enzyme bereitstellt, liefern virale Gene Informationen für die direkte Synthese der fehlenden Proteine.
Der Phage hat eine komplexe Art von Symmetrie: Der Kopf ist quaderförmig und der Prozess ist stabförmig (spermienförmig) (siehe Abb. 21, 22).
So werden Viren je nach Verpackungsmethode in quaderförmige, kugelförmige, stäbchenförmige und spermatozoide Formen unterteilt.
Das letzte Stadium der Virusreplikation ist die Freisetzung neuer Virionen in den Wirtsorganismus, wo sie benachbarte Zellen infizieren und den Replikationszyklus wiederholen können. Einige Viren werden freigesetzt, wenn die Wirtszelle stirbt, während andere Viren infizierte Zellen verlassen können, indem sie durch die Membran austreten, ohne die Zelle direkt abzutöten.
6 Bei Infektion mit dem Influenzavirus binden sich Glykoproteine \u200b\u200ban die Wirtsepithelzelle. Dadurch wird das Virus absorbiert. Abbildung 7 Viren verursachen beim Menschen eine Vielzahl von Krankheiten, von leichten bis zu schweren Krankheiten.
Einige Viren, die komplexer sind, haben eine Hülle namens Peplos. Es entsteht, wenn das Virus die Wirtszelle verlässt. In diesem Fall wird das virale Kapsid von der inneren Oberfläche der cytoplasmatischen Membran der Wirtszelle umhüllt und eine oder mehrere Schichten der Supercapsid-Hülle werden gebildet. Nur einige Viren haben eine solche Hülle, zum Beispiel die Viren Tollwut, Herpes, Enzephalitis. Diese Hülle enthält Phospholipide, die durch Äther abgebaut werden. Somit ist es durch Einwirken auf Äther möglich, ein Virus mit Peplos von einem Virus mit einem "nackten Kapsid" zu unterscheiden.
Der Impfstoff kann unter Verwendung abgeschwächter lebender Viren, abgetöteter Viren oder molekularer Untereinheiten des Virus hergestellt werden. Im Allgemeinen führen lebende Viren zu einer erhöhten Immunität, können jedoch in geringer Häufigkeit Krankheiten verursachen. Getötete Virusimpfstoffe und Viren von Untereinheiten können keine Krankheiten verursachen, führen jedoch im Allgemeinen zu einer weniger wirksamen oder langfristigen Immunität.
Abgeschwächte Lebendvirus-Impfstoffe wurden im Labor entwickelt, um bei Empfängern nur wenige Symptome zu verursachen und ihnen gleichzeitig Immunität gegen zukünftige Infektionen zu verleihen. Poliomyelitis war eine der Krankheiten, die einen Meilenstein bei der Verwendung von Impfstoffen markierten. Der Erfolg des Polio-Impfstoffs ebnete den Weg für die routinemäßige Verteilung von Impfstoffen für Kinder gegen Masern, Mumps, Röteln, Windpocken und andere Krankheiten. Lebendimpfstoffe werden normalerweise hergestellt, indem das Wildtyp-Virus geschwächt wird, indem es im Labor in Geweben oder bei Temperaturen gezüchtet wird, die sich von denen des Virus im Wirt unterscheiden.
Bei einigen Viren ragen Kapsomere in Form von Dornen (diese Dornen sind stumpf) aus der äußeren Lipidschicht der Hülle heraus. Solche Viren werden Peplomere genannt (z. B. Influenzavirus, siehe Abbildung 52).
Die Nukleinsäure des Virus ist ein Träger erblicher Eigenschaften, und das Kapsid und die äußere Hülle tragen Schutzfunktionen, als ob sie die Nukleinsäure schützen würden. Darüber hinaus erleichtern sie den Eintritt des Virus in die Zelle.
Beispielsweise kann das Virus in Zellen in vitro, Embryonen von Vögeln oder lebenden Tieren gezüchtet werden. Die Anpassung an diese neuen Zellen oder die Temperatur führt zu Mutationen im Genom des Virus, wodurch sie im Labor besser wachsen können und gleichzeitig ihre Fähigkeit beeinträchtigen, Krankheiten zu verursachen, wenn sie wieder in einen im Wirt gefundenen Zustand eingeführt werden. Diese abgeschwächten Viren verursachen daher immer noch Infektionen, wachsen jedoch nicht sehr gut, sodass sich die Immunantworten im Laufe der Zeit entwickeln können, um schwere Krankheiten zu verhindern.
Die Gefahr der Verwendung von Lebendimpfstoffen, die im Allgemeinen wirksamer sind als getötete Impfstoffe, ist ein geringes, aber signifikantes Risiko, dass diese Viren durch umgekehrte Mutationen in ihre krankheitsverursachende Form zurückkehren. Reverse Mutationen treten auf, wenn ein Impfstoff in einem Wirt mutiert, so dass er dem Wirt vorliest und erneut Krankheiten verursachen kann, die dann bei einer Epidemie auf andere Menschen übertragen werden können.
Virusgrößen... Viren werden in Nanometern gemessen. Ihre Größe schwankt in einem weiten Bereich von 15-20 bis 350-400 nm.
Methoden zur Virusmessung: 1) Filtration durch Bakterienfilter mit bekannter Porengröße; 2) Ultrazentrifugation - große Viren fallen schneller aus; 3) Fotografieren von Viren in einem Elektronenmikroskop.
In der Grippe hilft eine Mutation in Genen für Oberflächenmoleküle dem Virus, der schützenden Immunität zu entkommen, die es in der vergangenen Grippesaison möglicherweise erhalten hat, und macht es erforderlich, dass Menschen jedes Jahr geimpft werden. Andere Viren, wie Kinderkrankheiten, Masern, Mumps und Röteln, mutieren so wenig, dass Jahr für Jahr derselbe Impfstoff verwendet wird.
Impfstoffe und antivirale Medikamente zur Behandlung
In einigen Fällen können Impfstoffe zur Behandlung einer aktiven Virusinfektion eingesetzt werden. Bei Tollwut, einer tödlichen neurologischen Erkrankung, die im Speichel von mit Tollwut kontaminierten Viren übertragen wird, kann das Fortschreiten der Erkrankung vom Zeitpunkt des Tierbisses bis zum Eintritt in das Zentralnervensystem zwei Wochen oder länger dauern. Dies ist genug Zeit, um eine Person zu impfen, bei der der Verdacht besteht, von einem tollwütigen Tier gebissen zu werden, und die erhöhte Immunantwort durch die Impfung reicht aus, um zu verhindern, dass das Virus in das Nervengewebe eindringt.
Die chemische Zusammensetzung von Viren... Die Menge und der Gehalt an DNA- und RNA-Viren sind nicht gleich. In der DNA reicht das Molekulargewicht von 1 · 10 6 bis 1,6 · 10 8 und in der RNA von 2 · 10 6 bis 9,0 · 10 6.
Proteine \u200b\u200bin Virionen kommen in unbedeutender Anzahl vor, sie bestehen aus 16-20 Aminosäuren. Neben Kapsidproteinen gibt es auch interne Proteine, die mit Nukleinsäure assoziiert sind. Proteine \u200b\u200bbestimmen die antigenen Eigenschaften von Viren und schützen das Virus aufgrund der dichten Packung von Polypeptidketten auch vor der Wirkung von Wirtszellenenzymen.
Auf diese Weise werden die tödlichen neurologischen Folgen der Krankheit verhindert und das Individuum muss sich nur von dem infizierten Biss erholen. Dieser Ansatz wird auch zur Behandlung von Ebola verwendet, einem der schnellsten und tödlichsten Viren, die Menschen infizieren, obwohl es normalerweise begrenzte Populationen infiziert. Ebola ist auch die häufigste Todesursache bei Gorillas. Durch die Verwendung neu entwickelter Impfstoffe, die die Immunantwort verstärken, soll das Immunsystem der Betroffenen das Virus besser kontrollieren und möglicherweise die Mortalität senken.
Lipide und Kohlenhydrate befinden sich in der Außenhülle komplexer Virionen. Die Wirtszellmembran ist die Quelle für Lipide und Kohlenhydrate. Polysaccharide, die Teil einiger Viren sind, bestimmen ihre Fähigkeit, eine Agglutination von Erythrozyten zu verursachen.
Virusenzyme... Viren haben keinen eigenen Stoffwechsel und benötigen daher keine Stoffwechselenzyme. Es wurde jedoch festgestellt, dass einige Viren Enzyme aufweisen, die ihr Eindringen in die Wirtszelle erleichtern. Beispielsweise wurde im Influenza-A-Virus Neuraminidase gefunden, die in den Membranen tierischer Zellen (Erythrozyten usw.) enthaltene Neuraminsäure abspaltet. Phagen haben Lysozym, das die Zellmembran, Phosphatase usw. zerstört.
Eine andere Möglichkeit zur Behandlung von Virusinfektionen ist die Verwendung antiviraler Medikamente. Diese Medikamente haben oft eine begrenzte Fähigkeit, Viruserkrankungen zu heilen, wurden jedoch verwendet, um Symptome für eine Vielzahl von Viruserkrankungen zu kontrollieren und zu reduzieren. Bei den meisten Viren hemmen diese Medikamente das Virus, indem sie die Wirkung eines oder mehrerer seiner Proteine \u200b\u200bblockieren. Es ist wichtig, dass die Zielproteine \u200b\u200bvon viralen Genen codiert werden und dass diese Moleküle in einer gesunden Wirtszelle nicht vorhanden sind.
Somit wird das Viruswachstum gehemmt, ohne den Wirt zu schädigen. Da sind viele antivirale Medikamenteverfügbar zur Behandlung von Infektionen, von denen einige spezifisch für ein bestimmtes Virus sind und andere, die mehrere Viren betreffen können. Antivirale Medikamente wurden zur Behandlung von Herpes genitalis und Grippe entwickelt. Bei Herpes genitalis können Medikamente wie Aciclovir die Anzahl und Dauer von Episoden aktiver Viruserkrankungen verringern, bei denen Patienten virale Läsionen in ihren Hautzellen entwickeln.
Nachweis von viralen Antigenen... Virale Antigene in infizierten Wirtszellen können unter Verwendung von Immunfluoreszenztechniken nachgewiesen werden. Zubereitungen, die Zellen enthalten virus infiziertwerden mit spezifischen Immunlumineszenzseren behandelt. Bei Betrachtung unter einem Fluoreszenzmikroskop wird an Stellen, an denen sich Viruspartikel ansammeln, ein charakteristisches Leuchten beobachtet. Die Art des Virus wird durch die Entsprechung des spezifischen Lumineszenzserums bestimmt, das die Lumineszenz verursacht hat.
Da das Virus lebenslang im Nervengewebe des Körpers verborgen bleibt, ist dieses Medikament kein Heilmittel, kann jedoch die Symptome der Krankheit besser beherrschen. Bei der Grippe können Medikamente wie Tamiflu die Dauer der Grippesymptome um ein oder zwei Tage verkürzen, aber das Medikament verhindert die Symptome nicht vollständig. Andere antivirale Medikamentewie Ribavirin wurden verwendet, um eine Vielzahl von Virusinfektionen zu behandeln.
Wenn eines dieser Arzneimittel allein verwendet wird, kann das Virus aufgrund der hohen Mutationsrate des Virus schnell eine Resistenz gegen das Arzneimittel entwickeln. Viren sind zellfreie Einheiten, die normalerweise nur mit einem Elektronenmikroskop gesehen werden können. Viren sind vielfältig und infizieren Archaeen, Bakterien, Pilze, Pflanzen und Tiere. Viren bestehen aus einem Nukleinsäurekern, der von einem Proteinkapsid mit oder ohne äußere Lipidhülle umgeben ist.
Die Einschleppung des Virus in die Zelle, seine Wechselwirkung mit der Wirtszelle und die Reproduktion (Reproduktion) bestehen aus einer Reihe aufeinanderfolgender Stufen.
Stufe 1. Beginnt mit dem Adsorptionsprozess auf Kosten der Virion- und Zellrezeptoren. Bei komplexen Virionen befinden sich Rezeptoren auf der Oberfläche der Hülle in Form von Styloid-Auswüchsen (Influenzavirus), in einfachen Virionen, auf der Oberfläche des Kapsids.
Stufe 2. Das Eindringen des Virus in die Wirtszelle verläuft für verschiedene Viren unterschiedlich. Zum Beispiel durchdringen einige Phagen die Membran mit ihrem Prozess und injizieren Nukleinsäure in die Wirtszelle (siehe Kapitel 8). Andere Viren treten in die Zelle ein, indem sie mit Hilfe einer Vakuole ein Viruspartikel ansaugen, dh an der Stelle der Einführung in die Zellmembran wird eine Vertiefung gebildet, dann werden ihre Ränder geschlossen und das Virus erscheint in der Zelle. Dieser Rückzug wird als Viropexis bezeichnet.
Stufe 3. "Entfernen des Virus" (Zerfall). Für seine Reproduktion wird die virale Nukleinsäure von ihren schützenden Proteinabdeckungen (Hülle und Kapsid) befreit. Der Entkleidungsprozess kann während der Adsorption beginnen oder auftreten, wenn sich das Virus bereits in der Zelle befindet.
Stadium 4. In diesem Stadium erfolgt die Replikation (Reproduktion) von Nukleinsäuren und die Synthese von viralen Proteinen. Dieses Stadium tritt unter Beteiligung der DNA oder RNA der Wirtszelle auf.
Stufe 5. Zusammenbau des Virions. Dieser Prozess wird durch die Selbstorganisation von Proteinpartikeln um die virale Nukleinsäure erleichtert. Die Proteinsynthese kann unmittelbar nach der viralen Nukleinsäuresynthese oder nach einem Intervall von mehreren Minuten oder mehreren Stunden beginnen. Bei einigen Viren tritt eine Selbstorganisation im Zytoplasma auf. Andere haben Wirtszellen im Kern. Die Bildung der Außenhülle (Peplos) erfolgt immer im Zytoplasma.
Stadium 6. Die Freisetzung des Virions aus der Wirtszelle erfolgt durch Perkolation des Virus durch die Zellmembran oder durch ein in der Wirtszelle gebildetes Loch (in diesem Fall stirbt die Wirtszelle).
Arten der Virus-Zell-Interaktion... Der erste Typ, die produktive Infektion, ist durch die Bildung neuer Virionen in der Wirtszelle gekennzeichnet.
Der zweite Typ - abortive Infektion - besteht darin, dass die Nukleinsäurereplikation unterbrochen wird.
Der dritte Typ ist durch den Einbau von viraler Nukleinsäure in die DNA der Wirtszelle gekennzeichnet; Es gibt eine Form der Koexistenz des Virus und der Wirtszelle (Virogenität). In diesem Fall ist die Synchronität der viralen und zellulären DNA-Replikation gewährleistet. In Phagen wird dies Lysogenese genannt.
Mikroskopische Untersuchung... Bei einzelnen Virusinfektionen werden spezifische intrazelluläre Körper im Zytoplasma oder in den Kernen der Wirtszellen beobachtet - Einschlüsse, die diagnostischen Wert haben (Babesh-Negri-Körper bei Tollwut, Guarnieri-Körper bei Pocken usw.). Die Größe von Viruspartikeln und Körpereinschlüssen kann durch spezielle Verfahren zur Verarbeitung von Zubereitungen mit Beizmittel und Imprägnierung (z. B. das Verfahren zur Versilberung nach Morozov) künstlich erhöht und mit Immersionsmikroskopie beobachtet werden. Kleinere Virionen, die außerhalb der Sichtweite eines optischen Mikroskops liegen, werden nur elektronenmikroskopisch nachgewiesen. In Bezug auf intrazelluläre Einschlüsse gibt es unterschiedliche Sichtweisen. Einige Autoren glauben, dass sie eine Sammlung von Viren darstellen. Andere glauben, dass sie als Ergebnis der Reaktion der Zelle auf die Einschleppung von Viren entstehen.
Virusgenetik... Die Modifikation (nicht vererbte Veränderungen) von Viren ist auf die Eigenschaften der Wirtszelle zurückzuführen, in der sich das Virus vermehrt. Die modifizierten Viren erwerben die Fähigkeit, Zellen zu infizieren, die denen ähnlich sind, in denen sie modifiziert wurden. Verschiedene Viren manifestieren sich auf unterschiedliche Weise. Beispielsweise ändert sich die Form von "negativen Flecken" (Phagenkolonien) in Phagen.
Mutation - bei Viren tritt sie unter dem Einfluss derselben Mutagene auf, die eine Mutation bei Bakterien verursachen (physikalische und chemische Faktoren). Während der Nukleinsäurereplikation tritt eine Mutation auf. Mutationen beeinflussen verschiedene Eigenschaften von Viren, z. B. die Temperaturempfindlichkeit usw.
Die genetische Rekombination in Viren kann aus der gleichzeitigen Infektion einer Wirtszelle mit zwei Viren resultieren, während einzelne Gene zwischen zwei Viren ausgetauscht werden können und Rekombinanten gebildet werden, die die Gene zweier Elternteile enthalten.
Eine genetische Reaktivierung von Genen tritt manchmal auf, wenn ein inaktiviertes Virus mit einem vollständigen gekreuzt wird, was zur Rettung des inaktivierten Virus führt.
Die spontane und gerichtete Genetik von Viren ist für die Entwicklung des Infektionsprozesses von großer Bedeutung.
Beständig gegen Umweltfaktoren... Die meisten Viren werden durch Aktion inaktiviert hohe Temperaturen... Es gibt jedoch Ausnahmen, zum Beispiel ist das Hepatitis-Virus hitzebeständig.
ZU niedrige Temperaturen Viren sind nicht empfindlich, ultraviolette Sonnenstrahlen wirken inaktivierend auf Viren. Gestreutes Sonnenlicht wirkt weniger aktiv auf sie. Viren sind gegen Glycerin resistent, wodurch es möglich ist, sie lange Zeit in Glycerin zu halten. Sie sind gegen Antibiotika resistent (während der Kultivierung von Viren wird das Testmaterial mit Antibiotika behandelt, um die Bakterienflora zu unterdrücken).
Säuren, Laugen und Desinfektionsmittel inaktivieren Viren. Einige mit Formalin inaktivierte Viren behalten jedoch ihre immunogenen Eigenschaften bei, was die Verwendung von Formalin für Impfstoffe (Tollwutimpfstoff) ermöglicht.
Anfälligkeit von Tieren... Das Spektrum an anfälligen Tieren für einige Viren ist sehr breit, zum Beispiel sind viele Tiere empfindlich gegen Tollwutviren. Einige Viren betreffen nur eine Tierart, beispielsweise betrifft das Hundepestvirus nur Hunde. Es gibt Viren, für die Tiere nicht anfällig sind - zum Beispiel Masernvirus usw.
Organotropie von Viren... Viren können bestimmte Organe, Gewebe und Systeme infizieren. Zum Beispiel greift das Tollwutvirus das Nervensystem an. Das Pockenvirus ist dermatrop usw.
Freisetzung von Viren in die Umwelt... Von einem kranken Körper können Viren in den Kot ausgeschieden werden, beispielsweise das Polio-Virus und andere Enteroviren. Das Tollwutvirus wird im Speichel ausgeschieden, das Grippevirus wird in die Nasen-Rachen-Schleimhaut usw. ausgeschieden.
Die Hauptübertragungswege von Viren... Tröpfchen in der Luft (Influenza, Pocken), Lebensmittel (Poliomyelitis, Hepatitis A), Haushaltskontakt (Tollwut), übertragbar (Enzephalitis).
Antivirale Immunität... Der menschliche Körper hat eine angeborene Resistenz gegen bestimmte Viren. Zum Beispiel sind Menschen nicht anfällig für das Hundepestvirus. Tiere sind nicht anfällig für Masernviren. In diesen Fällen beruht die antivirale Immunität auf dem Fehlen von Zellen, die die Reproduktion von Viren unterstützen können.
Die antivirale Immunität wird sowohl durch zelluläre als auch durch humorale Abwehrfaktoren bestimmt, die nicht spezifisch und spezifisch sind. Unspezifische Faktoren. Ein starker Inhibitor der Virusreproduktion ist eine Proteinsubstanz - Interferon. BEIM gesunder Körper es ist in einer unbedeutenden Menge enthalten, und Viren tragen zur Produktion von Interferon bei, und seine Menge nimmt signifikant zu. Es ist nicht spezifisch, da es die Reproduktion verschiedener Viren blockiert. Es hat jedoch Gewebespezifität, dh Zellen verschiedener Gewebe bilden unterschiedliche Interferone. Es wird angenommen, dass sein Wirkungsmechanismus in der Tatsache liegt, dass er die Proteinsynthese in der Wirtszelle stört und dadurch die Reproduktion des Virus stoppt.
Spezifische Faktoren der antiviralen Immunität umfassen virusneutralisierende Antikörper, hämagglutinierende und präzipitierende Antikörper.
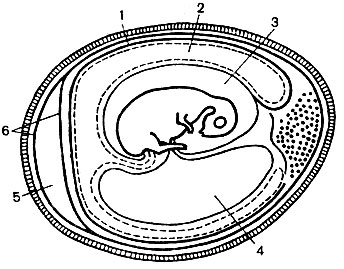
Methoden zur Viruskultivierung... Viren vermehren sich nur in lebensfähigen Zellen. Sie werden kultiviert: in Hühnerembryonen (Abb. 53), Gewebekulturen von Menschen und verschiedenen Tieren, im Körper empfindlicher Tiere anfällige Arthropoden.
In der ersten Phase der Entwicklung der Virologie war die Hauptmethode zur Untersuchung von Viren die künstliche Infektion von Tieren. Diese Methode ist jedoch kompliziert, und außerdem erwiesen sich Tiere als immun gegen viele Viren.
Von großer Bedeutung für die Entwicklung der Virologie war die Einführung von Methoden zur Kultivierung von Viren in Hühnerembryonen und in die Kultur menschlicher und tierischer Gewebezellen.
Hühnerembryo-Infektion... Zur Vermehrung von Viren werden Hühnerembryonen im Alter von 7 bis 12 Tagen verwendet, die in einem Thermostat bei 37 ° C inkubiert werden. Voraussetzung für die korrekte Entwicklung des Embryos ist die Einhaltung einer bestimmten Luftfeuchtigkeit, die durch Platzieren eines Gefäßes mit Wasser im Thermostat erzeugt werden kann.
Die Eignung eines Hühnerembryos für eine Infektion wird durch das Vorhandensein von Bewegungen des Embryos und eines entwickelten Netzwerks von Blutgefäßen auf der Chorion-Allantoic-Membran beim Scannen mit einem Ovoskop bestimmt.
Die Kultivierung von Viren in Hühnerembryonen erfolgt an verschiedenen Stellen des infizierten Embryos (siehe Abb. 53):
1) auf der Chorion-Allantoic-Membran,
2) in die allantoische Höhle;
3) in die Fruchtwasserhöhle;
4) in den Dottersack.
Hühnerembryonen werden in einer Schachtel mit sterilen Instrumenten infiziert. Vor der Infektion werden Hühnerembryonen zweimal mit einem mit Alkohol angefeuchteten Wattestäbchen abgewischt.
Infektion der Chorion-Allantoic-Membran. Nach der Desinfektion werden die Eier vorsichtig ein Stück der Schale vom stumpfen Ende abgeschnitten, die Schalenmembran wird entfernt und die Chorion-Allantoic-Membran wird gefunden. Infektiöses Material in einer Menge von 0,1 bis 0,2 ml wird mit einer Spritze oder einer Pasteurpipette auf die Chorion-Allantoic-Membran aufgetragen. Nach der Infektion wird das Loch mit einer Kappe verschlossen und der Spalt zwischen ihm und dem Hühnerembryo mit Paraffin gefüllt.
Schreiben Sie auf der anderen Seite des Eies mit Bleistift den Namen des infektiösen Materials und das Infektionsdatum.
Infektion in die Fruchtwasserhöhle. Das Ei wird ovoskopiert und an der lateralen Seite wird eine Stelle ausgewählt, an der der Chorion-Allantois keine großen Blutgefäße aufweist. Dieser Bereich ist mit einem Bleistift markiert. Die Eier werden in horizontaler Position auf einen Ständer gestellt, desinfiziert und ein Loch in der Schale mit einem speziellen sterilen Speer bis zu einer Tiefe von 213 mm durchbohrt, durch den eine Nadel mit infektiösem Material im gleichen Abstand direkt in die Fruchtwasserhöhle eingeführt wird. Um zu verhindern, dass die injizierte Flüssigkeit zurückfließt, wird über dem Airbag eine Punktion vorgenommen, wonach beide Löcher mit Paraffin gefüllt werden.
Infektion in die allantoische Höhle. Die Infektion wird in einer abgedunkelten Box durchgeführt. Der Luftraum wird notiert, die Schale über dem Luftraum wird desinfiziert und eine Spritzennadel mit dem Material wird durch das Loch in der Schale in Richtung des Embryos eingeführt. Wenn die Nadel in die allantoische Höhle gelangt, wird der Schatten des Embryos verschoben. Nach der Infektion wird das Loch mit Paraffin gefüllt.
Infektion im Dottersack. Die Schale wird desinfiziert. Das Ei wird mit dem stumpfen Ende nach rechts auf den Ständer gelegt, so dass der Dottersack nach oben zeigt. Über der Luftkammer in der Mitte ist ein Loch durchstochen. Durch das Loch in der Schale in horizontaler Richtung bis zu einer Tiefe von 2-3 mm wird eine Spritzennadel eingeführt, die in den Dottersack eintritt. Das Material wird in einem Volumen von 0,2 bis 0,3 ml injiziert. Nach dem Einbringen des Materials wird das Loch gewachst.
Das Temperaturregime und die Inkubationsdauer hängen von den biologischen Eigenschaften des eingeführten Virus ab.
Infizierte Eier werden täglich überprüft - Ovoskopie, um die Lebensfähigkeit des Embryos zu überprüfen. Wenn die Embryonen am ersten Tag sterben, ist die Ursache normalerweise ein Trauma während der Infektion. Solche Eier stammen aus Erfahrung.
Wenn es notwendig ist, jede Komponente des Embryos separat zu untersuchen, wird das Material in einer bestimmten Reihenfolge gesammelt: Das Allantois wird abgesaugt, dann wird das Fruchtwasser, die Chorion-Allanto-Membran geschnitten, das Fruchtwasser-Membran, der Embryo, der Dottersack werden getrennt und erst dann wird die Chorion-Allanto-Membran entfernt und vom Inneren getrennt Schalenoberfläche. Das Vorhandensein des Virus im infizierten Embryo wird durch die charakteristischen Veränderungen in der Chorion-Allantoic-Membran des infizierten Hühnerembryos bestimmt.
Viren ohne hämagglutinierende Aktivität werden mit RSK nachgewiesen.
Um das Virus in den allantoischen oder amniotischen Flüssigkeiten infizierter Embryonen nachzuweisen, wird RHA platziert (die Hämagglutination wird durch allantoische oder amniotische Flüssigkeiten oder eine aus der Chorion-Allantoic-Membran hergestellte Suspension verursacht).
Kultivierung von Viren in der Zellkultur... Zur Anreicherung von Viren in empfindlichen Zellkulturen werden Gewebe von Menschen und verschiedenen Tieren verwendet. Die größte praktische Anwendung fanden einschichtige Kulturen primärer trypsinierter und transplantierter Zelllinien.
Einschichtige Zellkulturen werden in flachen Glasmatratzengefäßen gezüchtet. Die Zellsuspension in einem flüssigen Nährmedium bei einer Temperatur von 37 ° C ermöglicht es, eine "in vitro" -Zellschicht mit einer bestimmten histologischen Struktur zu erhalten. Das Vorhandensein von Viren in Gewebekulturen wird durch die Veränderung (Degeneration) von Zellen nachgewiesen. Die Art der Viren wird bestimmt, indem die Wirkung der Viren durch Zugabe geeigneter typspezifischer Seren zum geimpften Material neutralisiert wird.
Diese Methoden ermöglichen eine schnellere Einbeziehung von Forschungsergebnissen und sind kostengünstiger. In Fällen, in denen Viren keine zytopathische Wirkung (Degeneration) verursachen und sich nicht in Hühnerembryonen entwickeln, werden Methoden zur Infektion von Tieren angewendet (siehe Kapitel 11).
Zur Kultivierung von Viren werden kontinuierliche Zellen verwendet, die häufiger aus Zellen bösartiger Tumoren gewonnen werden.
Einschichtkulturen werden aus menschlichen, Hühner- und Tierembryonen erhalten.
Der Vorteil von einschichtigen Zellkulturen ist die Einfachheit der Technik und die einfache Abrechnung.
Die Fähigkeit von Zellen, sich außerhalb des Körpers zu vermehren, hängt mit dem Grad der Gewebedifferenzierung zusammen. Weniger differenzierte Gewebe haben eine größere Proliferationsfähigkeit (Bindegewebe, Epithelgewebe).
Das Wesentliche der Verfahren zur Herstellung von Primärgewebekulturen ist die Zerstörung des interzellulären Gewebes und die Trennung von Zellen für die anschließende Herstellung einer Monoschicht.
Die Dissoziation von Zellen erfolgt durch Einwirken auf das Gewebe proteolytischer Enzyme, meistens Trypsin. Trypsinlösung fördert die Trennung von Zellen unter Beibehaltung ihrer Fortpflanzungsfähigkeit. Wachsende Zellkulturen erfordern ein Nährmedium. Die Zusammensetzung des Mediums ist komplex und enthält eine Reihe von Bestandteilen: Aminosäuren, Glucose, Vitamine, Mineralsalze, Coenzyme usw. Die Gewebekultur wird unter streng aseptischen Bedingungen erhalten. Dem Medium werden Antibiotika (500 U Penicillin und 250 U Streptomycin in 1 ml) zugesetzt, um das Wachstum der Bakterienflora zu unterdrücken.
Das vorbereitete Gewebe wird mit einer 0,25% igen Lösung von erwärmtem Trypsin gegossen und in einem Thermostat bei 37 ° C inkubiert. Während der Inkubation wird das Gewebe periodisch durch Drehen des Kolbens gerührt. Trypsinierte Zellen werden 5 Minuten bei 800-1000 U / min zentrifugiert.
Trypsinisierung und Zentrifugation werden sehr sorgfältig durchgeführt, um die Zellen nicht zu verletzen. Nach der Zentrifugation wird der Überstand entfernt und das Zellsediment in ein kleines Volumen Kulturmedium gegeben. Um eine homogene Masse zu erhalten, wird die Zellsuspension durch eine Mullschicht in einem Trichter (steril) filtriert. Die Zellsuspension wird durch Inokulieren von 0,1 ml in 2 Röhrchen mit Zuckerbrühe auf Sterilität überprüft.
Der Erfolg der Zellkultivierung hängt von der Inokulationsdosis ab, daher werden die Zellen nach der Trypsinisierung in der Goryaev-Kammer gezählt. Nach dem Zählen wird die Zellsuspension mit einem Nährmedium so verdünnt, dass 1 ml 500.000-1.000.000 Zellen enthält und in Reagenzgläser und Matratzen gegossen wird. Gewebekulturröhrchen werden in einer geneigten Position in einem Inkubator inkubiert.
Die beimpften Kulturen werden täglich unter einem Mikroskop mit geringer Vergrößerung betrachtet, um ihre Wachstumsmuster zu bestimmen. Normale proliferierende Zellen sind hell und wachsen in einer einzigen Schicht. Wenn die Zellen dunkel und körnig sind und sich nicht vermehren, was auf eine Kontamination (schlechte Handhabung von Geschirr oder Kontamination von Zutaten) zurückzuführen sein kann, werden solche Kulturen aus dem Experiment entfernt.
Ein Wechsel des Nährmediums 2-3 Tage nach der Aussaat verbessert die Proliferationsintensität.
Normale, gut proliferierende Zellen sind mit dem Testmaterial infiziert.
Transplantierte Kulturen werden hauptsächlich aus bösartigen Tumoren gewonnen. Hela-Stamm - Kultur von Gebärmutterhalskrebszellen einer Frau namens Helena (erhalten 1950); Stamm Hep-2, isoliert von einem Patienten mit Kehlkopfkrebs. Das Wachstum dieser Zellen wird in Laboratorien durch aufeinanderfolgende Passagen aufrechterhalten. Ihre Besonderheit liegt in der Tatsache, dass sie sich lange reproduzieren. Gegenwärtig haben diese Zellen Tausende von Generationen durchlaufen. Während der Passage verlieren sie einige morphologische und biochemische Eigenschaften - sie unterliegen Mutationen. Sie bleiben jedoch für die Kultivierung von Viren in ihnen gut geeignet. Die Kultur dieser Zellen wird von Labors auf der ganzen Welt verwendet.
Die Reproduktion des Virus in der Zellkultur erfolgt zu unterschiedlichen Zeiten, abhängig von den Eigenschaften des Virus und dem Zelltyp.
Das Vorhandensein des Virus wird anhand der zytopathischen Wirkung beurteilt. Zelldegeneration wird im Mikroskop beobachtet. Der Zeitpunkt der zytopathischen Wirkung und seine Art hängen von der Dosis und den Eigenschaften des Virus ab.
Bei einigen Viren wird nach einigen Tagen eine zytopathische Wirkung festgestellt (Pockenvirus), bei anderen nach 1-2 Wochen (Hepatitis-Virus usw.).
Derzeit sind bereits Hunderte von Viren bekannt, die Menschen infizieren. Der Kampf gegen Virusinfektionen erfolgt nach verschiedenen Methoden. Die Immunisierung ist am effektivsten. Auf diese Weise wurden Pocken beseitigt und die Inzidenz von Poliomyelitis verringert. Soziale Prävention ist wichtig im Kampf gegen Virusinfektionen - Zerstörung streunender Hunde (Kampf gegen Tollwut), persönliche Prävention usw.
Diese Maßnahmen können jedoch nicht die Beseitigung aller Viruserkrankungen gewährleisten. Wissenschaftler suchen beharrlich nach Möglichkeiten, das Virus zu infizieren, ohne die Zelle zu beschädigen, in der es sich befindet.
Daher ist es selbstverständlich, dass die Virologie im Programm der Kommunistischen Partei der Sowjetunion als einer der führenden Zweige der Naturwissenschaften bezeichnet wird, die in den kommenden Jahren vorrangig weiterentwickelt werden sollten.
Grundlegende Methoden zur Erforschung von Viren... 1. Reaktion der Hämagglutination, Reaktion der verzögerten Hämagglutination, Reaktion der indirekten Hämagglutination. Komplementbindungsreaktion.
2. Die Reaktion der Neutralisation von Viren in Gewebekulturen.
3. Methode der Immunfluoreszenz.
4. Histologische Methode - Identifizierung von Einschlüssen (Babeshs kleine Körper - Negri - mit Tollwut; Pashens kleine Körper - mit Pocken usw.).
5. Biologische Methode.
Virusinfektion Eine Zelle ist ein Komplex von Prozessen, die sich aus der Wechselwirkung einer Zelle mit einem viralen Genom ergeben. Das extrazelluläre Virion ist inert, und diese Inertheit bleibt bestehen, bis das virale Genom als unabhängige genetische Einheit intrazellulär zu funktionieren beginnt. Das Hauptmerkmal, das der unten angegebenen Klassifizierung zugrunde liegt, ist die Beziehung zwischen den Genomen des Virus und der Zelle. Wenn sich das virale Genom unabhängig von der Replikation des zellulären Genoms repliziert, handelt es sich um eine autonome Art der Virusinfektion. Wenn das virale Genom im zellulären Genom enthalten ist und sich mit diesem repliziert, ist dies ein integrativer Typ. Jede dieser beiden Haupttypen der Virusinfektion kann je nach Bildung des Virus (produktive und abortive Infektion), je nach Ergebnis des Infektionsprozesses für die Zelle (lytische und nicht-lytische Infektion) und je nach Dynamik der Wechselwirkung zwischen Virus und Zelle (akute und chronische Infektion) in verschiedene Formen unterteilt werden ).
Eine autonome Art der Virusinfektion einer Zelle ist charakteristisch für die meisten bekannten Viren von Tieren und Menschen. Bei dieser Art von Infektion repliziert sich das virale Genom unabhängig von der Replikation des zellulären Genoms.
Eine produktive autonome Virusinfektion ist durch die Bildung eines vollständigen Virus gekennzeichnet. Dies ist die am besten untersuchte Form der Interaktion zwischen Viren und Zellen. Die in Kapitel 3 diskutierten grundlegenden Mechanismen der Virusreproduktion wurden für diese spezielle Form der Infektion untersucht und beschrieben.
Eine autonome abortive Virusinfektion tritt auf, wenn die Wechselwirkung des Virus mit der Zelle nicht zur Bildung vollständig wertvoller Virionen führt. Dies tritt normalerweise auf, wenn Zellen mit einem defekten Virus infiziert sind; wenn das Virus in die gegen es resistenten Zellen eindringt; wenn anfällige Zellen unter nicht zulässigen Bedingungen mit einem vollständig wertvollen Virus infiziert sind.
Eine lytische autonome Virusinfektion führt zum Zelltod. Das Virus, das den Zelltod verursacht, wird als zytopathogen bezeichnet, und Zellschäden werden mit dem Begriff "Zytop, togene Wirkung" bezeichnet. Die Art der zytopathologischen Veränderungen bei Virusinfektionen ist sehr unterschiedlich. Es könnte degu sein
Zellversagen (Vakuolisierung des Zytoplasmas, Kernpyknose, Chromosomenaberrationen), Bildung spezifischer viraler intrazellulärer Einschlüsse, Entstehung von Riesenzellen wie Polykaryozyten oder Symplasten. Die Zelllyse ist das letzte Stadium der Wirkung von zytopathogenen Viren. Für die Manifestation der Zytopathogenität erfordert das Virus nicht die Reproduktion des Genoms und die Implementierung aller genetischen Informationen.
Eine nicht-lytische autonome Virusinfektion ist charakteristisch für Viren, die eine äußere Hülle aus Lipoprotein enthalten und Zellen durch Knospen aussenden. Retro-, Orthomyxo-, Paramyxo- und Arenaviren können eine produktive nicht-lytische Zellinfektion verursachen. In den typischsten Fällen ist jedoch eine nicht-lytische Natur für abortive Infektionen charakteristisch, wenn die genetischen Eigenschaften von Viren, die für die Implementierung zytopathogener Funktionen erforderlich sind, fehlen oder nicht auftreten.
Entsprechend der Dynamik des Infektionsprozesses werden Virusinfektionen in akute und chronische unterteilt. Wann akute Infektion In der infizierten Zelle kommt es zur Bildung von Virionen, wonach der Infektionsprozess stoppt. Bei einer chronischen Infektion produziert die infizierte Zelle bis zu ihrem natürlichen Tod Virionen oder virale Komponenten (nicht aufgrund einer bestimmten Virusinfektion), während die Tochterzellen infiziert bleiben. Eine produktive und abortive lytische Infektion ist immer akut. Eine produktive nicht-lytische Infektion ist immer chronisch. Eine abortive nicht-lytische Infektion verläuft in der Regel chronisch, kann aber auch akut auftreten.
Der integrative Typ der Virusinfektion ist durch die physikalische Vereinigung (Integration) des viralen und zellulären Genoms gekennzeichnet. In diesem Fall repliziert sich das virale Genom und fungiert als integraler Bestandteil des zellulären Genoms. Diese Art der Virusinfektion ist für Retroviren obligatorisch und bei Papa, Ado und Herpesviren möglich.
Integrative Infektionen können nach denselben Parametern wie autonome klassifiziert werden - Produktivität, Dynamik und Ergebnis; Die Integrationsbeziehung zwischen dem Virus und der Zelle kann auch produktiv und fehlgeschlagen sein. Eine produktive integrative Infektion ist typisch für Retroviren. Abtreibung, eine integrative retrovirale Infektion, tritt unter den gleichen Bedingungen auf, wie oben für autonome abortive Infektionen beschrieben. Somit fehlen defekte Retroviren
Neue Teile des Genoms (einige Stämme des Rous-Sarkom-Virus, Sarkom-Viren von Mäusen, Katzen, Affen, Leukämie-Viren von Frend- und Rauscher-Mäusen usw.) können für sich genommen nur eine abortive integrative Infektion verursachen, die sich in Gegenwart eines Virus in eine produktive Infektion verwandeln kann Assistent. Ein Beispiel für eine abortive integrative Infektion, die durch ein vollständig wertvolles Retrovirus in nicht permissiven Zellen verursacht wird, ist die Infektion vogelvirus Rous-Sarkom von Säugetierzellen. Gleichzeitig findet in den Zellen ein abortiver Integrationsprozess statt, der durch eine verringerte Produktion von viraler RNA, die Akkumulation eines unvollständigen Sortiments an viralen Proteinen, eine Beeinträchtigung ihrer Verarbeitung und das Fehlen der Bildung von Virionen gekennzeichnet ist.
In der Regel sind integrative Infektionen nicht lytisch, wie im Fall von Retroviren der Typen B, C, D, onkogenen Papov-, Adeno- und Herpesviren, aber es gibt auch lytische Infektionen, die durch das zytopathogene Retrovirus der Vogel-Retikuloendotheliose oder das "schäumende" Virus verursacht werden.
Eine virogene integrative Infektion ist eine abortive Infektion, die in eine produktive Form übersetzt werden kann. Zellen mit dieser Form der Infektion können spontan oder unter dem Einfluss spezieller (physikalischer, chemischer, biologischer) Einflüsse beginnen, ein infektiöses Virus zu produzieren. Dieses Phänomen wird als Virogenität bezeichnet, und die Zellen werden in Analogie zu Lysogenie und lysogenen Zellen im phagenbakteriellen System als virogen bezeichnet.
Mehr zum Thema Viruszellinfektion:
- Virusinfektionen und Infektionen mit Verdacht auf virale Ätiologie
- Bakterielle und virale Infektionen in der Luft: Influenza, Parainfluenza, Adenovirus-Infektion, respiratorische Synzytialinfektion. Bakterielle Bronchopneumonie, Lappenpneumonie.