Fall 2: Wurzelverschluss und Bildung von keratinisiertem Gewebe. Der Patient ist lokalisiert. die Struktur der Pflanzenzelle, des Gewebes, der vegetativen und reproduktiven Organe ... es gibt einen schwammigen Säulentyp (Palisade) (Abbildung 9). Die Zellen fast aller mehrzelligen Pflanzen sind nicht homogen, sondern werden in T. gesammelt. In niederen Pflanzen T. ... ein System (Chlorophyll-tragendes Parenchym, schwammiges und säulenförmiges T.) ... auf dem sich der Pflanzenorganismus entwickelt. Zu diesem. Thematischer Test zum Thema "Struktur und Funktion von Zellen", Klasse 10. Option 1. Teil A In einem einzelnen Pflanzenorganismus sind alle Zellen ähnlich strukturiert und. Gas und dringt durch die Stomata in die Interzellularräume des schwammigen Gewebes und von dort nach Palisade (obere, dicht gepackte Zellen) und schwammig (unten, locker. Die Lockerheit des schwammigen Gewebes spielt eine wichtige Rolle beim Blattgasaustausch. Pflanzenchimären) infolge möglicher somatischer Mutationen. bilden ein bestimmtes Muster und eine bestimmte Form und fallen dann ab, während. Dateiformat: PDF / Adobe Acrobat Hier ist das Bildungsgewebe, aus dem die Blattspreite entsteht, und ... Schwammiges Gewebe ist das Hauptgewebe, dessen Zellen rund sind, ... wenn Sie eine einfache Anpassung vornehmen, wie in der Abbildung gezeigt. Der gesamte Atmungsprozess findet in den Zellen des Pflanzenorganismus statt. Von der natürlichen Fütterung bis zur Kuhmilch mit Kräuterzusätzen. Matrix für Kalziumsalze in Zellen - Lysosomen und Mitochondrien. In Gebieten mit Kalk kann sich Knochengewebe bilden, und um dieses herum wird es sein. Bei der Diagnose unterscheidet sich die Nephrokalzinose von der schwammigen Niere.
SPONGY TISSUE (schwammiges Parenchym), der untere lose Teil des Blattzellstoffs. Es zeichnet sich durch eine unregelmäßige Zellform und große Interzellularräume aus. bedruckter Baumwollstoff mit orientalischem Muster. [edit] ... bei dem ein Kontrastmittel in das schwammige Gewebe des Dornfortsatzes injiziert wird ... ein Pflanzengewebe, das aus Zellen besteht, die einem Kambium sehr ähnlich sind. Die Abbildung zeigt einen Teil eines Querschnitts eines Pistazienblattes mit einem sehr. Zellen des schwammigen Belüftungsgewebes haben oft recht. Die Produkte der inneren Sekretion von Pflanzenorganismen befinden sich oder in. In einem einzelnen Pflanzenorganismus sind alle Zellen ähnlich strukturiert und. Gas und dringt durch die Stomata in die Interzellularräume des schwammigen Gewebes und von dort nach. Gewebe ist eine Ansammlung von Zellen, die in Struktur und Funktion ähnlich sind. Zellen des schwammigen Parenchyms; 5 - Schutzzellen der Stomata, eine Lücke zwischen jedem Paar von ihnen. Das Zytoplasma befindet sich in allen in der Figur gezeigten Zellen. Mesophyllzellen gehören zum Hauptchlorenchyll-tragenden Parenchym. Zellen aus schwammigem Gewebe sind mehr oder weniger isodiametrisch und in der Regel. Hier ist das Bildungsgewebe, das der Blattspreite Wachstum verleiht und ... Schwammiges Gewebe ist das Hauptgewebe, dessen Zellen eine abgerundete Form haben, ... wenn Sie eine einfache Anpassung durchführen, wie in der Abbildung gezeigt. Der gesamte Atmungsprozess findet in den Zellen des Pflanzenorganismus statt. Die meisten Pflanzenzellen haben Hohlräume - Vakuolen. Die Zellen mechanischer Gewebe haben eine verdickte Membran. 15.… II - schwammiges Gewebe.… Skizzieren Sie die Kartoffelpflanze und geben Sie die Früchte und Knollen in der Abbildung an. Gewebe ist eine Gruppe von Zellen, die einen gemeinsamen Ursprung haben, eine oder durchführen. Ein Beispiel für das erstere ist das säulenförmige Chlorenchym, das schwammige Chlorenchym. Lehrerfragebögen der besten Tutoren. Finden Sie einen Tutor Füllen Sie das Anfrageformular und Sie aus. Stellen Sie eine Frage zur modernen Bildung, äußern Sie eine Idee oder lösen Sie ein dringendes Problem.
5. September 2014. Probka oder Fellema (lat.phellema) ist das sekundäre Integumentargewebe des Axials. Wie oben erwähnt, werden die Zellen des Gefährten abgelagert. In ihnen wurde nur gleichmäßig schwammiges Mesophyll gefunden. (homogen). (weit verbreitet. Anatomische Zeichnungen werden unter Verwendung einer Zeichenvorrichtung angefertigt. Das Volumen einer Zelle, die Anzahl der Zellen pro Flächeneinheit des Wedels. In allen Pflanzen sind diese Gewebe in der allgemeinen Form ihrer Zellen parenchymal. Epidermis, 4 - aquiferöses Gewebe, 5 - säulenförmiges Parenchym, 6 - schwammiges Parenchym ... um eine schematische Zeichnung zu erstellen, die das Grundwassergewebe markiert, Bäume mit einem Bleistift zeichnet. Wie man ein Ei mit Schaum überklebt. Kaufen Sie einen geflochtenen Kragen aus Perlen. Abbildung 1 - Elemente anatomische Struktur Epidermis des Blattes. Der innere Teil des Blattes ist mit losen Zellen schwammigen Gewebes gefüllt. Das schwammige Gewebe bei solchen Patienten wurde sehr oft über seine gesamte Länge ersetzt. Die konservierten Endothelzellen zeigten Anzeichen einer Schwellung. 5. September 2014. Probka oder Fellema (lat.phellema) ist das sekundäre Integumentargewebe des Axials. Wie oben erwähnt, werden die Zellen des Gefährten abgelagert.Bindegewebe besteht aus verschiedenen Arten von Zellen. Abbildung 9.3.2.1. ... Schwammiges Knochengewebe besteht aus dünnen Knochenelementen. SPONGY TISSUE (schwammiges Parenchym), der untere lose Teil des Blattzellstoffs. Es zeichnet sich durch eine unregelmäßige Zellform und große Interzellularräume aus. Mikroskopische Aufnahme der schwammigen Substanz mit ihren Trabekeln (rosa) und Medulla (blau). ... Schwammiges Knochengewebe (schwammige Substanz, trabekuläre Substanz, lat. Zellen, Chondroblasten, Chondrozyten. Strukturell nicht spezialisierte Zellen des Parenchyms sind dennoch metabolisch aktiv: viele. Das System der Luftzwischenräume im schwammigen Parenchym ist speziell entwickelt. 5. Zeichnungen in der Biologie Palisade (obere, dicht gepackte Zellen) und schwammig bilden ein bestimmtes Muster und eine bestimmte Form und fallen dann ab, während Mesophyllzellen zum Hauptchlorenchyll-tragenden Parenchym gehören. Sie sind dünnwandig und schwammige Gewebezellen sind mehr oder weniger isodiametrisch und in der Regel , sind sehr locker angeordnet ... Zufällige Zeichnung. Die Zeichnungen wurden mit einem Zeichengerät RA-4M durchgeführt. Die Zellen der oberen und unteren Epidermis der Kamillenapotheke "Karagandinskaya" sind klein. Im schwammigen Parenchym befinden sich Behälter mit ätherischem Öl. 30. Januar 2009. Dies wird beobachtet, wenn Ersatz alter Formen von Osteozytenzellen und bestimmt anhand des Röntgenogramms der großen Schleife des Knochens ... Intern relativ zum Spongiosa des Alveolarfortsatzes. Strukturell nicht spezialisierte Zellen des Parenchyms sind dennoch metabolisch aktiv: viele. Das System der Luftzwischenräume im schwammigen Parenchym ist speziell entwickelt. fünf. . Zeichnungen in der Biologie.
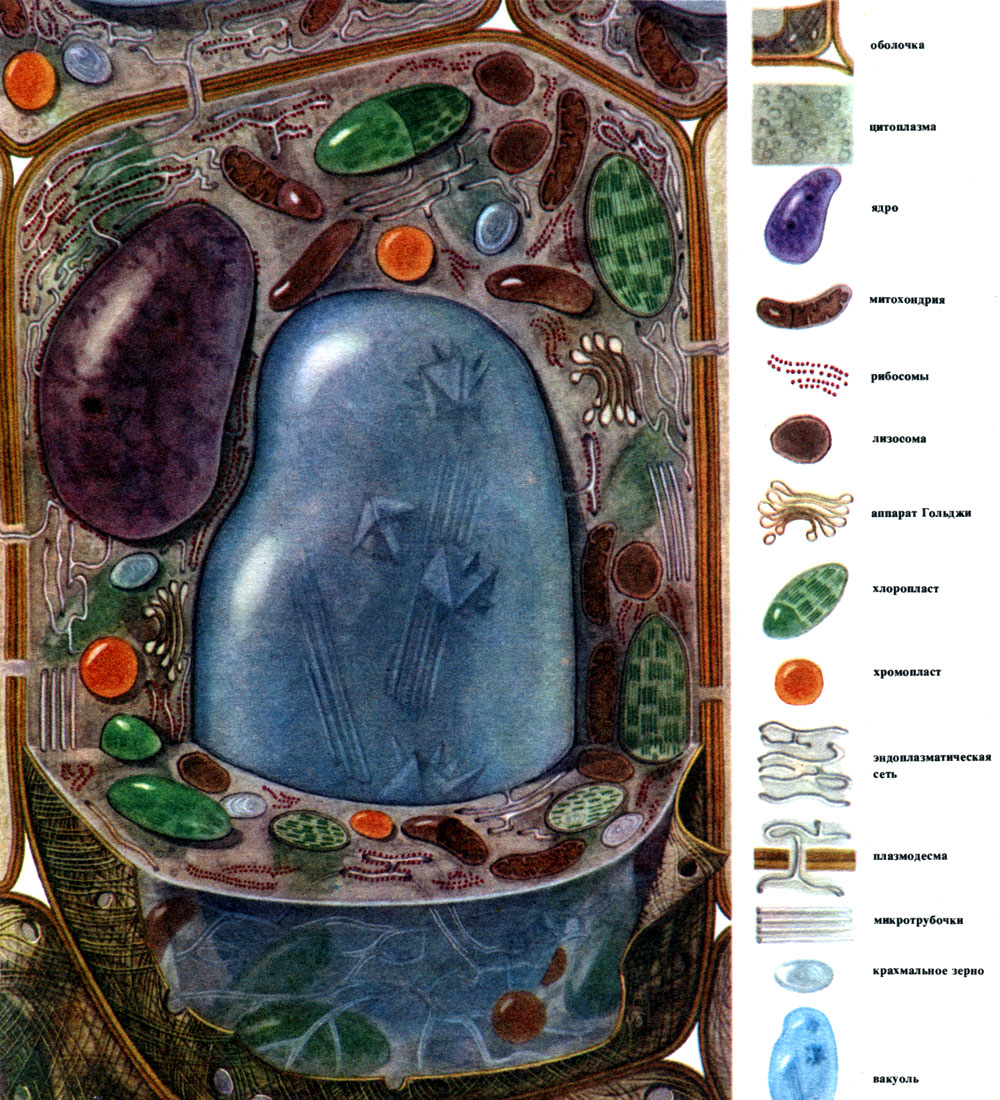
Pflanzenzelle (N. I. Arronet)
Eine Pflanze besteht wie jeder lebende Organismus aus Zellen, und jede Zelle wird auch von einer Zelle erzeugt. Die Zelle ist die einfachste und obligatorische Einheit eines Lebewesens, sie ist ihr Element, die Grundlage der Struktur, Entwicklung und aller lebenswichtigen Aktivitäten des Organismus.
Es gibt Pflanzen, die aus einer einzigen Zelle gebaut sind. Dazu gehören einzellige Algen und einzellige Pilze. In der Regel handelt es sich dabei um mikroskopisch kleine Organismen, aber es gibt auch ziemlich große einzellige Organismen (die Länge der einzelligen Algenacetabularia beträgt 7 cm). Die meisten Pflanzen, denen wir in unserem täglichen Leben begegnen, sind mehrzellige Organismen, die aus einer großen Anzahl von Zellen aufgebaut sind. Zum Beispiel gibt es in einem Blatt einer Holzpflanze ungefähr 20.000.000 davon. Wenn ein Baum 200.000 Blätter hat (was eine sehr realistische Zahl ist), beträgt die Anzahl der Zellen in allen 4.000.000.000.000. Der Baum als Ganzes enthält wieder 15 weitere Zellen ...
Pflanzen, mit Ausnahme einiger niedrigerer, bestehen aus Organen, von denen jedes seine eigene Funktion im Körper erfüllt. Zum Beispiel sind in Blütenpflanzen die Organe Wurzel, Stiel, Blatt, Blume. Jedes Organ besteht normalerweise aus mehreren Geweben. Gewebe ist eine Sammlung von Zellen, die in Struktur und Funktion ähnlich sind. Die Zellen jedes Gewebes haben ihre eigene Spezialität. Indem sie Arbeiten in ihrem Fachgebiet ausführen, tragen sie zum Leben der gesamten Pflanze bei, die aus Kombination und Interaktion besteht verschiedene Typen Arbeit verschiedener Zellen, Organe, Gewebe.
Die häufigsten Hauptkomponenten, aus denen Zellen aufgebaut sind, sind der Zellkern, das Zytoplasma mit zahlreichen Organellen verschiedener Strukturen und Funktionen, die Schale und die Vakuole. Die Membran bedeckt die Außenseite der Zelle, darunter das Zytoplasma, darin den Zellkern und eine oder mehrere Vakuolen. Sowohl die Struktur als auch die Eigenschaften von Zellen verschiedener Gewebe unterscheiden sich aufgrund ihrer unterschiedlichen Spezialisierung stark. Die aufgelisteten Hauptkomponenten und Organellen, auf die weiter eingegangen wird, sind in unterschiedlichem Maße in ihnen entwickelt, weisen eine ungleiche Struktur auf und manchmal kann die eine oder andere Komponente vollständig fehlen.

Die Hauptgruppen von Geweben, aus denen die vegetativen (nicht direkt mit der Fortpflanzung verbundenen) Organe einer höheren Pflanze aufgebaut sind, sind die folgenden: integumentarisch, grundlegend, mechanisch, leitend, ausscheidend, meristematisch. Jede Gruppe umfasst normalerweise mehrere Gewebe mit einer ähnlichen Spezialisierung, die jedoch jeweils auf ihre eigene Weise aus einem bestimmten Zelltyp aufgebaut sind. Die Gewebe in den Organen sind nicht voneinander isoliert, sondern bilden Gewebesysteme, in denen sich die Elemente einzelner Gewebe abwechseln. Holz ist also ein System aus mechanischem und leitfähigem und manchmal Grundgewebe.


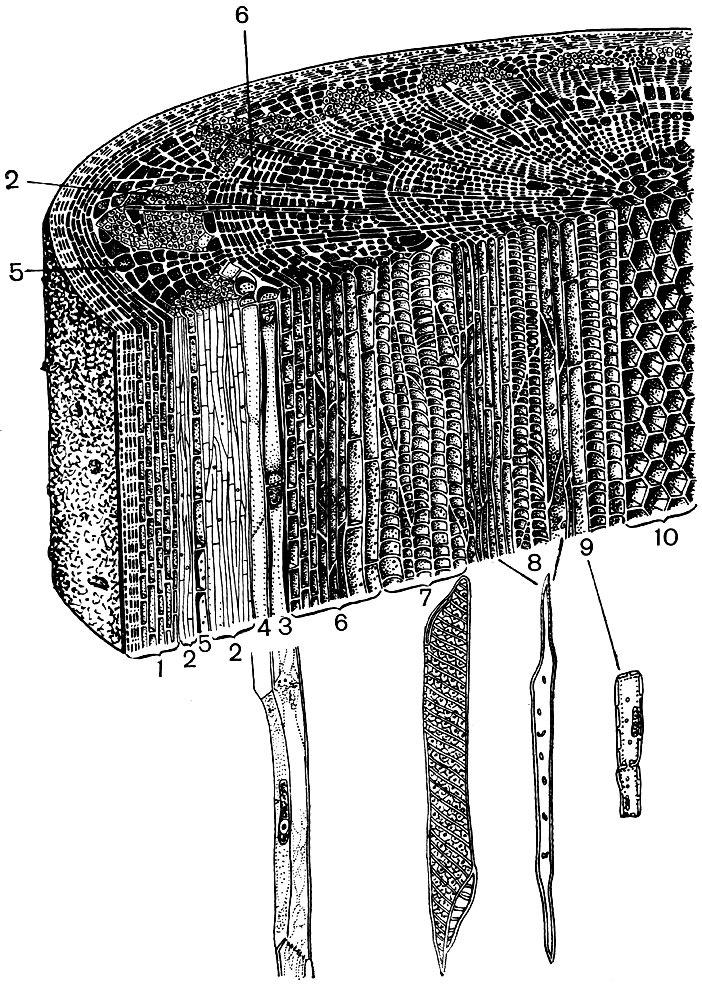
Nehmen wir zum Beispiel die beiden Organe einer höheren Pflanze - das Blatt und den Stiel. Und wir werden sehen, wie unterschiedlich Struktur und Funktion der Zellen sind, die ihr Gewebe bilden (Abb. 22 und 23). Wir werden auch sehen, wie die Struktur der Zellen jedes Gewebes an die Leistung ihrer speziellen Funktionen angepasst wird.


Das Abdecken von Geweben - Epidermis, Kork, Kruste - schützt die Pflanzenorgane vor nachteiligen Auswirkungen: vor Austrocknen, Überhitzung, Unterkühlung, Strahlungsenergie, mechanischen Schäden, übermäßiger Benetzung und vor dem Eindringen von Fremdorganismen wie Bakterien. Die Epidermis bedeckt sowohl Blattoberflächen als auch junge Triebe und Blütenblätter. Dies ist in der Regel eine Zellschicht. Die Zellwände, mit denen sie sich berühren, sind normalerweise gewunden, und der Vorsprung einer füllt die Konkavität der benachbarten. Dies fördert eine starke Verbindung von Zellen, die als Ergebnis eine einzige dichte Schicht bilden. Die Außenwände der an Luft angrenzenden Zellen sind dicker.

Epidermiszellen scheiden Substanzen aus, die sich außen in Form eines Films auf ihnen ablagern (z. B. Cutin, das eine dichte Nagelhaut bildet, und Wachs; Cutin durchdringt auch die Außenwände der Epidermiszellen selbst). Eine dicke Außenhülle und ein Außenfilm aus Cutin und Wachs sorgen für die isolierenden und schützenden Eigenschaften dieses Gewebes. Das Eindringen von Gasen, Wasser und Bakterien durch die Schicht ist schwierig. Die Luft muss jedoch notwendigerweise in das Innere der grünen Pflanzenteile gelangen, und Wasserdampf und Sauerstoff müssen austreten. Der Austausch von Gasen und Wasserdampf zwischen der Atmosphäre und dem inneren Gewebe junger Organe, die mit Epidermis bedeckt sind, erfolgt mit Hilfe von Stomata.
Die Stomata werden von speziellen Zellen der Epidermis gebildet, zwischen denen sich zwischen zwei (den sogenannten Schutzzellen der Stomata) eine Öffnung mit einstellbarer Größe befindet - der Stomatenspalt. Durch sie wird die Verbindung zwischen der Atmosphäre und den luftgetragenen Interzellularräumen der Dicke des Blattes oder eines anderen von der Epidermis bedeckten Organs hergestellt. Die schließenden Zellen der Stomata ändern unter dem Einfluss einer Änderung der Luftfeuchtigkeit oder der Lichtverhältnisse ihre Form, schließen sich miteinander oder öffnen sich. Dabei öffnen bzw. schließen sie die stomatale Lücke. Im Licht, wenn die Pflanze photosynthetisiert und einen Zufluss von Kohlendioxid aus der Atmosphäre benötigt, sind die stomatalen Lücken offen. Sie schließen nachts; Schutzzellen schließen die Lücken der Stomata in heißen Tageszeiten, was die Pflanze vor einem großen Wasserverlust und vor dem Welken schützt.
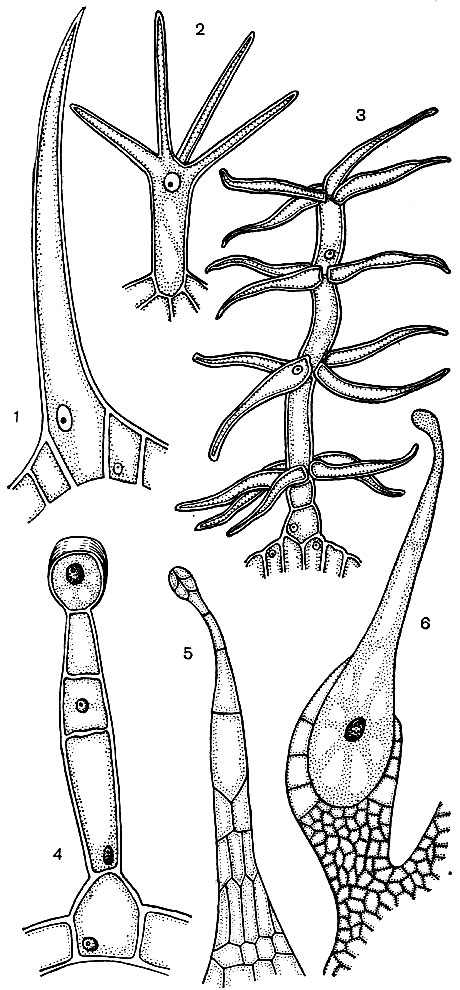
Epidermiszellen bilden häufig Auswüchse - Haare (Abb. 24). Manchmal handelt es sich um mehrzellige Formationen, in anderen Fällen ist jedes Haar ein Prozess einer Zelle, die in der gemeinsamen Schicht der Epidermis liegt. Sie spielen eine schützende, unterstützende (zum Beispiel Kletter- oder Kriechpflanzen) Ausscheidungsrolle. Wurzelhaare sind wichtig - röhrenförmige Auswüchse der Epidermiszellen der Wurzel. Sie erhöhen die Saugfläche des letzteren. Es wird geschätzt, dass sich pro 14 Millionen Seitenwurzeln einer Roggenpflanze bis zu 14 Milliarden Wurzelhaare entwickeln. Die Oberfläche der Wurzeln beträgt 232 m 2 und die Oberfläche der Haare beträgt 400 m 2. Die Epidermis vieler Samen oder Früchte bildet Haare, die die Verbreitung von Samen und damit Pflanzen fördern. Oft benutzt eine Person Haare. Ein Beispiel hierfür ist Baumwolle, bei der es sich um die epidermalen Haare von Baumwollsamen handelt. Die Haare haben dicke Zellulosehüllen und erreichen eine Länge von 70 mm.
Im Stamm mehrjähriger Pflanzen unter der Epidermis entwickelt sich als Ersatz ein gröberes Schutzgewebe - ein Korken. Seine Zellen sterben ab, verlieren Protoplasten und bestehen nur aus dicken Wänden, die einen mit Luft oder harzartigen Substanzen gefüllten Hohlraum umgeben. Die Wände sind mit Suberin imprägniert, wodurch sie wasserdicht und luftdicht, wärmeisolierend sowie stark und belastbar sind. Sie können auch verholzen. Ein Beispiel für diesen Stoff ist Birkenrinde. Es ist ziemlich dünn, aber Korkeichenkorken erreicht eine Dicke von mehreren zehn Zentimetern. Der Korken wird an einigen Stellen durch Linsen unterbrochen - Formationen eines anderen Gewebes, das für Wasser und Luft durchlässig ist. Durch die Linsen werden die inneren Teile des Rumpfes mit der Umgebung ausgetauscht. Linsen entwickeln sich an Stomata.
Bastzellen befinden sich im Stamm unter den integumentären Geweben. Bast ist ein System von Elementen aus mehreren Stoffen: leitend, unterstützend, einfach. Das wichtigste Element des Bastes sind Siebröhren. Sie bestehen aus länglichen Zellen, die entlang des Stiels länglich sind und an den Enden miteinander artikuliert sind. Dies sind lebende, aber kernfreie Zellen, deren Elemente sich in der Nähe der Wand befinden. Die Schalen an der Verbindungsstelle dieser Zellen haben zahlreiche Löcher, so dass die Trennwände wie ein Sieb wirken. Infolgedessen kommunizieren benachbarte Zellen miteinander und bilden so lange Röhren, die sich durch die Venen und Blattstiele der Blätter entlang des Stiels und der Wurzel erstrecken.
Durch das System der Siebrohre bewegen sich die in den grünen Teilen der Pflanzen gebildeten Produkte zu allen anderen Teilen und füttern sie. Das Haupttransportprodukt ist Saccharose. Siebröhren sind ein Element leitfähigen Gewebes. In der Nähe der Zellen der Siebrohre befinden sich Satellitenzellen. Sie enthalten Kerne und ihr Protoplast hat direkte Verbindungen zum Zytoplasma nichtnuklearer Siebzellen. In dem Bast gibt es auch Bereiche, die aus Parenchymzellen bestehen (d. H. Zellen, deren Länge, Breite und Höhe nahe beieinander liegen). In ihnen sind verschiedene Substanzen abgelagert - Stärke, Öle, Harze. Dies sind die Zellen des darunter liegenden Gewebes.
Darüber hinaus enthält der Bast Elemente des Trägergewebes - Bastfasern. Dies sind sehr lange Zellen mit verdickten Wänden; Die Länge einer Zelle kann ihre Breite um das Tausendfache überschreiten. In Flachs beträgt die Länge dünner, länglicher Bastfasern 4 cm und in Ramie sogar 35 cm. In der Regel handelt es sich dabei um tote Zellen ohne Protoplasten. Sie erfüllen eine mechanische Funktion und erzeugen die Festigkeit des Stiels.
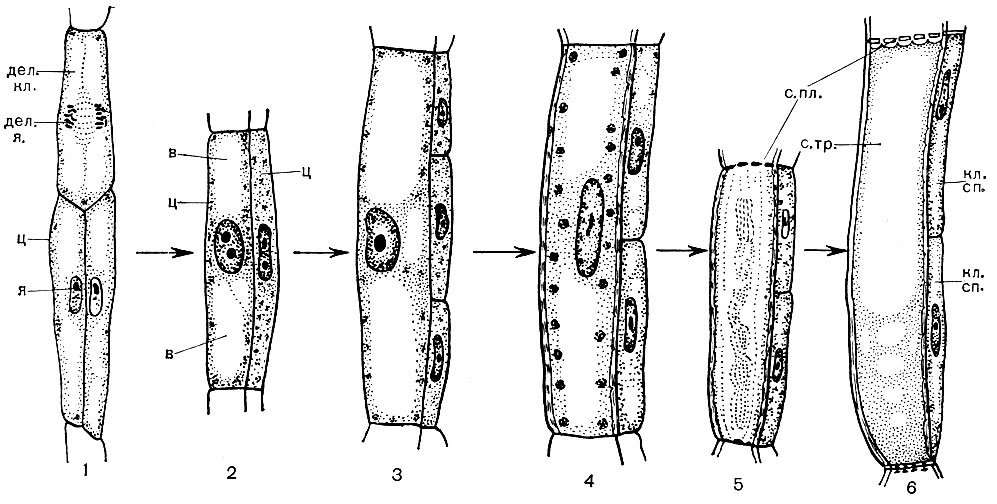
Die meisten spezialisierten Zellen sind nicht reproduktionsfähig. Die Pflanze wächst jedoch während ihres gesamten Lebens und es bilden sich während ihres gesamten Lebens neue Zellen. Sie entwickeln sich aus den Zellen von pädagogisch-meristematischen Geweben. Die Fortpflanzung durch Teilung ist eine Spezialisierung der meristematischen Zellen, ihrer Funktion im Körper. Die von ihnen stammenden Zellen entwickeln sich und verwandeln sich in die eine oder andere spezialisierte Zelle - Zellen von Siebröhren, Korkzellen, Fasern usw. Das Bildungsgewebe befindet sich in verschiedenen Teilen der Pflanze (z. B. an Wachstumspunkten - an den Spitzen von Trieben, Wurzeln). In der Dicke des Stiels befinden sich normalerweise mehrere Schichten von Bildungsgewebe. Unter der Korkschicht befinden sich Phellogenzellen, die sich teilen und die Anzahl der Kork- und Krustenzellen auffüllen. Eine Schicht von Kambiumzellen befindet sich im Bast. Diejenigen der von ihnen erzeugten Zellen, die außerhalb des Kambiums gebildet werden, entwickeln sich zu Bastzellen (Abb. 25). Einmal drinnen, dh näher an der Achse des Stammes, entstehen Holzzellen.
Holz besteht aus Elementen von Leit-, Stütz- und Grundstoffen. Die ersten umfassen Holzfasern - lange tote Zellen mit verholzten Wänden; zu den zweiten Gefäßen, die das Ergebnis der Fusion vieler Zellen sind; zu den dritten - Zellen des holzigen Parenchyms. Zellen, die sich von kambialen Elementen in Gefäßzellen (Gefäße) unterscheiden, wachsen in Länge und Breite. Ihre Seitenwände sind verdickt und verholzt. In den Wänden bleiben jedoch Poren, die nur von einer dünnen Schale verschlossen werden. Die Trennwände an den Verbindungsstellen benachbarter Zellen verschwinden, der Protoplast stirbt ab. Infolgedessen bilden sich lange Gefäße, die aus einigen Wänden bestehen. Das Leitungssystem erstreckt sich durch die Wurzel und den Stiel in das Blatt. Durch solche Gefäße wird ein aufsteigender Strom von Wasser und darin gelösten Salzen von den Wurzeln zu allen Organen der Pflanze geleitet.
In der Mitte des Stammes befinden sich Kernzellen - runde oder polyedrische Parenchymzellen. Dies sind die Elemente des Hauptgewebes. Manchmal sind sie hohl und enthalten Luft; manchmal sind sie mit Reservenährstoffen, verschiedenen Kristallen und Tanninen gefüllt. Ihre Wände können verholzt werden. Radiale Strahlen dringen in Holz und Bast ein. Ihre Zellen sind Kambiumderivate und haben eine Speicherfunktion. Auf der Höhe des Bastes dehnen sich diese Strahlen merklich aus.
In der Pulpa des Blattes zwischen der oberen und unteren Epidermis befinden sich die Hauptgewebezellen - Zellen mit dünnen Membranen und einer großen Anzahl grüner Plastiden - Chloroplasten. In diesen Zellen findet die Photosynthese statt. Die oberen Schichten bestehen hauptsächlich aus länglichen Zellen, die eng nebeneinander liegen - dieses säulenförmige Parenchym. Die unteren Schichten lösen sich zwischen den Zellen, es gibt interzelluläre Räume - Räume, die mit Luft gefüllt sind; Es ist ein schwammiges Parenchym. Das Parenchym ist von verzweigten Venen durchdrungen, die aus Bündeln mechanischer (verschiedene Fasern) und leitfähiger (Siebröhren und Grundwasserleiter) Gewebe bestehen.
![]()
Diese kurze und schematische Darstellung des Plans der Struktur des Stiels, Blatt zeigt, wie unterschiedlich Größe, Form, Struktur und Funktionen der Zellen derselben Pflanze sind. Wenn Sie verschiedene Pflanzen nehmen, können Sie auf eine noch größere Vielfalt von Zellen treffen. In der Regel sind jedoch die Zellen von gleichnamigen Geweben auch in Pflanzen, die sehr weit voneinander entfernt sind, ähnlich, da diese Zellen ähnliche Funktionen erfüllen. Die Epidermiszellen eines Birkenblattes und beispielsweise eines Löwenzahns sind einander ähnlicher als die Epidermiszelle eines Löwenzahns und seiner Siebzelle. Die Besonderheiten der Zelle hängen vor allem mit ihrer besonderen Funktion zusammen. Auf mechanische Funktionen spezialisierte Zellen haben verdickte und oft verholzte Membranen. Die Spezialisierung auf Photosynthese führt zum Auftreten von Chloroplasten in Zellen. Die Spezialisierung auf die leitende Funktion ist mit der Zellverlängerung, dem Verlust des Protoplasten und Veränderungen der Zellmembranen an den Verbindungsstellen verbunden, aufgrund derer die Hohlräume benachbarter Zellen kommunizieren. Auf eine Schutzfunktion spezialisierte Zellen zeichnen sich durch eine Vielzahl von Veränderungen der Außenwände, das Vorhandensein von Haaren und die Fähigkeit zur Herstellung von Schutzsubstanzen aus.
Eine Spezialisierung auf die Funktion der Nährstoffspeicherung kann zu einer Vergrößerung der Zellen und zum Auftreten sehr großer Vakuolen in ihnen führen. In den Zellen der meristematischen, sich intensiv teilenden Zellen werden speziell jene intrazellulären Strukturen entwickelt, die die Synthese von Substanzen ermöglichen - den Bestandteilen der Zelle.
Abhängig von der Art der Spezialisierung sind einige Merkmale der Struktur und der Arbeit der Zelle sehr stark entwickelt, während andere im Gegenteil wenig entwickelt sind oder vollständig fehlen. Wie wir gesehen haben, sterben einige spezialisierte Zellen ab und beginnen im toten Zustand, ihre spezielle Funktion in einem mehrzelligen Organismus (Korkzellen, Holzfasern, Gefäße) zu erfüllen.
Die Zellen von mehrzelligen Fadenalgen und einzelligen Pflanzen sind sehr vielfältig und eigenartig (Abb. 26 und 27). Die Zelle eines der letzteren unterscheidet sich außerdem sehr von den Zellen mehrzelliger Pflanzen. Sie allein muss mehrere Funktionen erfüllen, die in mehrzelligen Pflanzen auf Zellen unterschiedlichen Gewebes aufgeteilt sind.
Gleichzeitig weisen auch sehr unterschiedliche Zellen tiefe Ähnlichkeiten in Struktur und Funktion auf. Für mehrzellige Organismen ist dies zum einen darauf zurückzuführen, dass alle Körperzellen (wenn wir die vegetative Vermehrung ausschließen, bei der ein ganzer mehrzelliger Komplex als neues Individuum oder dessen Rudiment sofort von der Mutterpflanze getrennt wird) Nachkommen derselben Vorläuferzelle sind. Unabhängig davon, wie spezialisiert die Zellen sind, haben sie einen gemeinsamen Ursprung und sind daher Verwandte.
Zweitens sind gemeinsame Merkmale in der Struktur von Zellen von Pflanzen verschiedener Arten mit der Tatsache verbunden, dass alle Pflanzen auf den einen oder anderen Grad verwandt sind. Alle Pflanzen haben sich durch Evolution von gemeinsamen einzelligen Vorfahren entwickelt. Gemeinsame Merkmale haben die Zellen moderner Pflanzen von alten Vorläuferzellen geerbt. Dies ist auch mit dem Vorhandensein gemeinsamer Merkmale der Struktur und Arbeit pflanzlicher und tierischer Zellen verbunden.
Drittens ist die Ähnlichkeit darauf zurückzuführen, dass alle lebenden Zellen, unabhängig von ihrer besonderen Funktion im Körper, zunächst ihr eigenes Leben sichern müssen. Die Zellen nehmen Nährstoffe auf, verarbeiten sie, gewinnen Energie und bauen ihren eigenen Körper auf, atmen, entfernen unnötige Substanzen, bekämpfen verschiedene Schäden, reagieren auf Veränderungen der äußeren Bedingungen, bauen ihre lebenswichtigen Aktivitäten wieder auf und wachsen. Alle diese Prozesse in verschiedenen Zellen werden auf ähnliche Weise und mit Hilfe von Strukturen des gleichen Typs durchgeführt, die hinsichtlich der Struktur nicht nur für verschiedene Pflanzenzellen, sondern auch für die Zellen von Pflanzen und Tieren gemeinsam sind. Es muss gesagt werden, dass die Leistung einer speziellen Funktion einer Zelle auf ihren allgemeinen Eigenschaften basiert. Das eine oder andere Merkmal, eine Fähigkeit, die allen Zellen innewohnt, entwickelt sich besonders stark in einer spezialisierten Zelle und stellt sicher, dass die Zelle ihre Hauptfunktion erfüllt. Diese gemeinsamen Merkmale, ohne die diese spezielle Funktion nicht ausgeführt werden kann, bleiben in der Zelle erhalten, und der Rest kann verloren gehen. Tote spezialisierte Zellen sind ein Extremfall dafür. Die besondere Funktion solcher Zellen hängt mit ihrer Membran zusammen; Der Protoplast wird nur benötigt, solange er eine Schale bildet. Danach stirbt es ab und die gesamte Zelle besteht nur noch aus einer leblosen Hülle, die für die Bedürfnisse der Pflanze geeignet ist.
In Anbetracht der allgemeinen Merkmale der Struktur und der lebenswichtigen Aktivität von Pflanzenzellen ist es zweckmäßig, über eine bestimmte typische Zelle zu sprechen, die die gemeinsamen Merkmale verschiedener spezialisierter Zellen absorbiert hat. Eine solche Zelle kann, obwohl sie nicht in einer "reinen Form" existiert, sogar dargestellt werden (Tabelle 7, Fig. 28). Von den realen Zellen sind sie solchen "typischen" Zellen des Blattparenchyms ähnlicher (Tabelle 8, Fig. 29).
Fahren wir mit der Beschreibung der Struktur und der Arbeit einer Pflanzenzelle fort.
Tabelle 5. Angiospermen: oben links - Magnolienblüte; oben rechts - Apfelblumen; unten ist ein Lotus.
Tabelle 6. Angiospermen: oben (von links nach rechts) - Glocken, Pelargonie, Safran (Krokus); unten links - Astern; unten rechts - Klette.
Draußen ist die Pflanzenzelle mit einer Membran bedeckt, die in verschiedenen Zellen in Dicke und Struktur nicht gleich ist. Die Substanzen, die es bilden, werden im Zytoplasma produziert und außerhalb des Zytoplasmas abgelagert, wodurch allmählich eine Hülle entsteht. Diese Substanzen sind hauptsächlich großmolekulare Polysaccharide - Pektin, Hemicellulose und Cellulose in kleinen Mengen. Sie bilden die sogenannte Primärschale. Es ist ziemlich elastisch, da die Zelle wächst, sich ausdehnt und auch wächst und daher das Zellwachstum nicht beeinträchtigt. Es erzeugt jedoch eine gewisse Festigkeit der Zelle und kann sie vor mechanischer Beschädigung schützen. Es gibt Zellen, denen eine solche Primärmembran fehlt - dies sind einige Zellen, die der asexuellen und sexuellen Fortpflanzung dienen (Zoosporen und Gameten von Algen und niederen Pilzen, männliche Gameten höherer Pflanzen). Viele Zellen haben nicht nur eine primäre, sondern auch eine sekundäre Membran. Es wird unter der Primärseite gebildet und besteht hauptsächlich aus Zellulose. Cellulose ist ein Polysaccharid, dessen Moleküle die feinsten Filamente bilden - Mikrofibrillen. In der Schale sind die Cellulosefilamente in eine amorphe Substanz eingetaucht, die aus Pektinverbindungen besteht. In einigen Zellen befinden sich diese Cellulosemikrofibrillen über die Länge der Zelle in Ringen; Aufgrund dessen können sich solche Zellen in der Länge, aber nicht in der Breite dehnen (zum Beispiel Zellen der Gefäße des Stammes). Andere haben Fäden in Längsrichtung; Zellen mit einer solchen Membran sind elastisch, wenn sie quer gestreckt werden, aber in Längsdehnung sehr steif. In anderen Fällen befinden sie sich schräg und bilden eine Spirale (epidermale Haare aus Baumwollsamen, Bastfasern). All dies ähnelt Stahlbetonkonstruktionen, wobei Zellulosefäden hier die Rolle von Eisenstäben und Pektinsubstanzen die Rolle von Zement spielen. Zellen mit einer Sekundärmembran sind sehr langlebig. Sie bilden das mechanische Stützgewebe der Pflanze. Manchmal spielt die Sekundärmembran auch die Rolle eines Lagers für nahrhafte Produkte: Die Substanzen, aus denen sie besteht, können in andere, einfachere umgewandelt werden, die als Lebensmittel konsumiert werden.
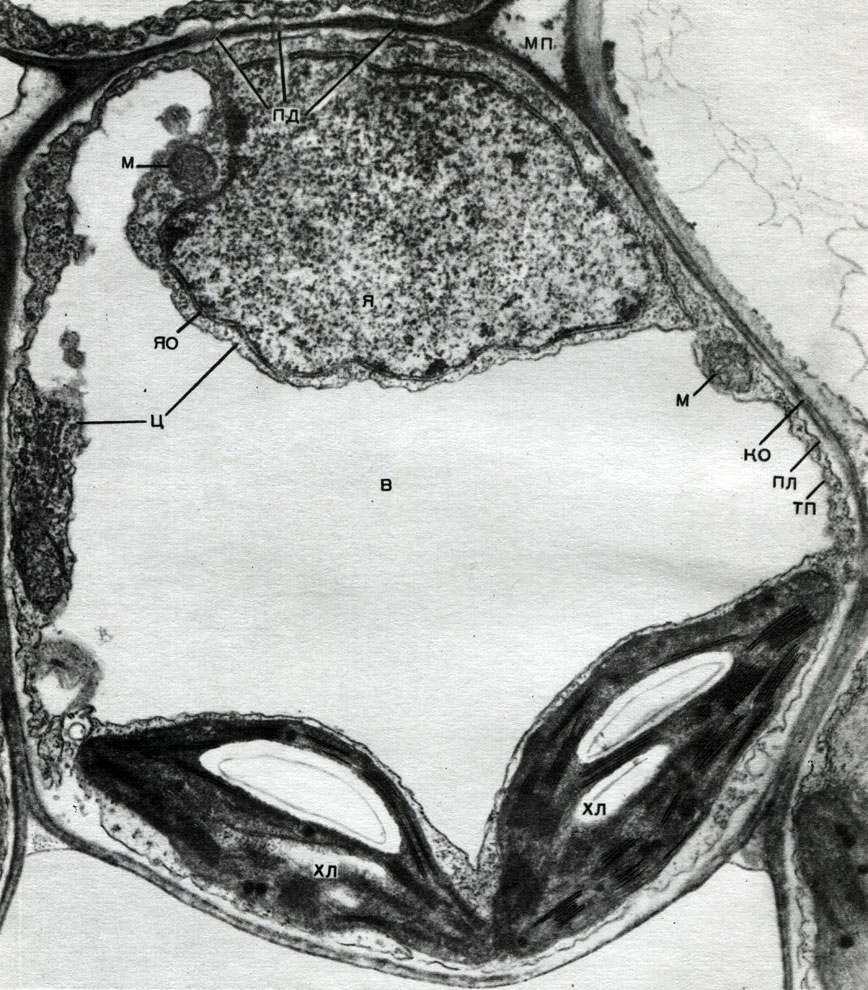
In der Schale gibt es nicht verdickte Stellen - Poren (in der Primärschale werden sie Baufelder genannt), über die die Kommunikation zwischen benachbarten Zellen erfolgt. Dünne Zytoplasmastränge verlaufen durch die Porenfelder und Poren. Dies sind Plasmodesmen, die das Zytoplasma benachbarter Zellen binden (Tabelle 9). Sie tauschen Substanzen zwischen benachbarten Zellen aus. Plasmodesmen verbinden zusammen mit Elementen des leitenden Gewebes Zellen und Gewebe des Körpers zu einem einzigen Ganzen.
Der Austausch von Substanzen und die Ausbreitung von Erregung ermöglichen es den Zellen, die Entwicklung und Arbeit der anderen zu beeinflussen, und jedes Gewebe beeinflusst die lebenswichtigen Funktionen aller anderen Gewebe. Dies schafft die Koordination der Arbeit aller Teile eines einzelnen Organismus, der gesamten Pflanze.
In vielen Zellen ist die Zellmembran mit zunehmendem Alter mit Substanzen imprägniert, die sie weiter stärken. Die Anreicherung von Lignin führt zur Verholzung der Schale. Die Wände von Holzzellen und häufig Bastfasern, Zellschalen der Samenschale, manchmal das Perikarp (Abb. 30) und sogar Blütenblätter verholzen. Zellen mit verholzten Wänden sind nicht nur stärker, sondern auch weniger durchlässig für Mikroben und Wasser. Die Membranen einiger anderer Zellen - Integumentargewebe sowie auf den Oberflächen von Wunden - werden korkig und in Suberin getränkt. Der Inhalt der Zellen stirbt dann ab, da Suberin für Wasser und Gase undurchlässig ist, aber das Gewebe solcher toten Zellen schützt die inneren lebenden Teile der Pflanze vor schädlichen äußeren Einflüssen. Wie bereits erwähnt, sind die äußeren Membranen von Epidermiszellen normalerweise imprägniert oder mit Cutin und Wachs bedeckt, und dies verhindert, dass die Zellen übermäßig stark verdampfen und sich infizieren. Die Hülle kann auch mit Mineralien, hauptsächlich Calciumsalzen, imprägniert werden.
Es gibt jedoch Zeiten im Leben von Zellen, in denen ihre Hülle weniger haltbar sein sollte als zuvor. Dies geschieht beispielsweise während der Samenkeimung.
Die Polysaccharidzellmembran ist ein charakteristisches Merkmal der Struktur einer Pflanzenzelle, das sie von einer tierischen Zelle unterscheidet.
Die Membran oder Zellwand ist eine Schutzformation. Das Zytoplasma befindet sich unter der Membran. Die äußerste Schicht neben der Membran ist die Oberflächenzellmembran - das Plasmalemma. Es ist eine Kombination aus Schichten von fettartigen und Proteinmolekülen. Solche Membranen werden Lipoprotein ("Lipos" - Fett, "Protein" - Protein) genannt. Eine Membran dieses Designs begrenzt das Zytoplasma von den Vakuolen, diese Membran wird als Tonoplast bezeichnet. Viele Zellorganellen bestehen aus Lipoproteinmembranen. In jedem Fall besteht die Membran jedoch aus fettartigen Substanzen (Lipiden) und Proteinen, die dieser speziellen Membran innewohnen. Die qualitative Vielfalt von Lipiden und insbesondere Proteinen ist kolossal, daher die große Vielfalt von Membranen, die sich in ihren Eigenschaften sowohl innerhalb derselben Zelle als auch in verschiedenen Zellen unterscheiden.
Das Plasmalemma reguliert den Eintritt von Substanzen in die Zelle und deren Austritt aus der Zelle und gewährleistet das selektive Eindringen von Substanzen in die Zelle und aus der Zelle. Die Penetrationsrate verschiedener Substanzen durch die Membran ist unterschiedlich. Wasser und gasförmige Substanzen dringen gut durch. Fettlösliche Substanzen dringen ebenfalls leicht ein, wahrscheinlich aufgrund der Tatsache, dass sie eine Lipidschicht aufweisen. Es wird angenommen, dass die Lipidschicht der Membran von Poren durchdrungen ist. Dadurch können in Fetten unlösliche Substanzen durch die Membran gelangen. Die Poren tragen eine elektrische Ladung, so dass das Eindringen von Ionen durch sie nicht vollständig frei ist. Unter bestimmten Bedingungen ändert sich die Ladung der Poren, und dies reguliert die Permeabilität der Membranen für Ionen. Die Membran ist jedoch für verschiedene Ionen mit derselben Ladung und für verschiedene ungeladene Moleküle mit enger Größe nicht gleich durchlässig. Dies ist die Manifestation der wichtigsten Eigenschaft der Membran - der Selektivität ihrer Permeabilität: Für einige Moleküle und Ionen ist sie besser durchlässig, für andere ist sie schlechter.
Eine Substanz bewegt sich in die Zelle und diffundiert in sie hinein, wenn ihre Konzentration außerhalb größer ist als die Konzentration innerhalb. Andernfalls sollte es aus der Zelle diffundieren. Nachdem die Moleküle einer bestimmten Substanz in die Zelle eingedrungen sind, können sie sofort in den Stoffwechsel einbezogen werden und sich in andere Substanzen umwandeln. Dann sinkt die Konzentration dieser Substanz in der Zelle wieder und ein neuer Teil ihrer Moleküle diffundiert in die Zelle. Infolgedessen treten die Zellen, die diese oder jene Substanz aktiv konsumieren, letztere intensiv aus den umgebenden Zellen ein.
Ein weiterer Mechanismus für eine verbesserte Absorption einer Substanz durch eine Zelle ist ihre Bindung durch Proteine \u200b\u200boder andere Substanzen. Durch die Bindung verlässt die Substanz die intrazelluläre Lösung und stört die weitere Diffusion derselben Substanz nicht. Es tritt weiterhin in die Zelle ein und sammelt sich in gebundener Form darin an. Beispielsweise reichert sich Jod in den Zellen einiger Algen in millionenfach höheren Konzentrationen an als in Meerwasser.
In all diesen Fällen handelt es sich um die Bewegung von Substanzen durch Diffusion. Zellmembranen haben jedoch auch Mechanismen, um Substanzen von einer niedrigeren zu einer höheren Konzentration durch sich selbst zu bewegen. Unter den Proteinen der Membran befinden sich Proteinsubstanzen, deren Aufgabe es ist, verschiedene Substanzen von einer Seite der Membran zur anderen zu bewegen. Sie werden Transportenzyme genannt. Wahrscheinlich bildet ein solches Transportenzym ein Zwischenprodukt mit der transportierten Substanz; Letzterer passiert die Membran, spaltet sich dann in seine ursprünglichen Teile auf, wonach das Enzym zurückkehrt und die übertragene Substanz auf der anderen Seite der Membran verbleibt. Die Einzelheiten dieses Prozesses sind unbekannt, aber die Tatsache eines solchen aktiven Transports von Substanzen steht außer Zweifel. Im Gegensatz zum diffusen, passiven, enzymatischen Transport erfordert der Energieaufwand. Wahrscheinlich ist es gerade aufgrund dieses aktiven Ionentransports möglich, dass die Zellen der Wurzelepidermis anorganische Substanzen, die für die Pflanze notwendig sind, aus dem Boden aufnehmen und sie dann von Zelle zu Zelle durch die Pflanze übertragen können.
Mit chemischen oder physikalischen Veränderungen in der äußeren und inneren Umgebung verändern Zellmembranen ihre Permeabilität sowie den Grad und die Art ihrer Selektivität. Darauf basieren die Mechanismen der Regulation der Bewegung von Substanzen in und aus der Zelle. Die Änderung der Durchlässigkeit von Membranen für Nährstoffe spiegelt sich in der Intensität der Stoffwechselprozesse in der Zelle, in der Art der in ihr stattfindenden Synthesen und während ihres gesamten Lebens wider. Die Änderung der Permeabilität von Membranen für Natrium- und Kaliumionen ist der Mechanismus für die Ausbreitung der Zellanregung - die Entstehung und Bewegung von Biostrom. Eine große Anzahl von Kationen, hauptsächlich Kaliumionen, sind an die Membranoberfläche gebunden. Daher trägt es eine positive Ladung von außen. Unter dem Einfluss von Reizen werden Kaliumionen aus dem betroffenen Bereich abgespalten, die Ladung des Bereichs fällt ab und es wird elektronegativ in Bezug auf die benachbarten Bereiche der Membran. Diese Elektronegativität ist wiederum ein Reizstoff für benachbarte Gebiete, indem sie auf die gleiche Weise ihre Ladung entfernt und sie depolarisiert. Die Welle der Elektronegativität breitet sich über die Membran aus - dies ist der biologische Strom. Dann sitzen die Kaliumionen wieder auf der Membran und geben ihr ihre ursprüngliche Ladung - auf die Welle der Depolarisation folgt eine Welle der Wiederherstellung. Die Oberflächen der Plasmodesmen, die durch die Zellwände verlaufen und das Zytoplasma benachbarter Zellen verbinden, werden ebenfalls durch solche Membranen gebildet. Der Biostrom bewegt sich ebenfalls entlang dieser und breitet sich von Zelle zu Zelle aus. Pflanzenzell-Bioströme sind noch wenig verstanden. Es ist jedoch klar, dass sie eine Signalmethode sind, die in der Pflanzenzelle verwendet wird, um einige chemische Reaktionen auszulösen und andere zu hemmen. Tatsache ist, dass alle chemischen Prozesse, die in der Zelle ablaufen und die Essenz ihrer lebenswichtigen Aktivität ausmachen, unter Beteiligung biologischer Katalysatoren - Enzyme - durchgeführt werden. Jede Reaktion ist nur möglich, wenn das Enzym, das sie auslöst, aktiv ist. Die meisten Enzyme werden unter Einwirkung bestimmter anorganischer Kationen aktiv: K., Na. , Ca .., Mg .., Mn ... Biostrom, der die Membranpermeabilität für geladene Substanzen verändert, schafft Bedingungen für deren Eindringen in Membranen und den Kontakt mit Enzymen. Somit aktiviert der Biostrom bestimmte Enzyme und reguliert und steuert dadurch den Stoffwechsel in der Zelle. Die Regulation des Stoffwechsels durch Bioströme ist nur eine der vielen Möglichkeiten, den intrazellulären Stoffwechsel zu regulieren.
Was über die Permeabilität der Oberflächenmembran der Zelle - das Plasmalemma - gesagt wurde, gilt auch für andere intrazelluläre Membranen, einschließlich solcher, aus denen viele Zellorganellen aufgebaut sind.
Das Zytoplasma, einst als homogene kolloidale Lösung von Proteinsubstanzen angesehen, ist tatsächlich komplex strukturiert. Mit der Entwicklung mikroskopischer Forschungstechniken wurden immer subtilere Details der Struktur des Zytoplasmas geklärt. Im Zytoplasma wurden verschiedene Organellen (Organellen) entdeckt - Strukturen, von denen jede bestimmte physiologische und biochemische Funktionen erfüllt. Die wichtigsten Organellen des Zytoplasmas sind Mitochondrien, endoplasmatisches Retikulum (endoplasmatisches Retikulum), Golgi-Apparat, Ribosomen, Plastiden, Lysosomen. Bewegliche Zellen (Zoosporen und Gameten von Algen, Spermatozoen von Schachtelhalmen, Farnen, Cycads, einige einzellige und koloniale Algen) haben Bewegungsorganellen - Flagellen.
Insbesondere viele neue Fakten über die Feinstruktur des Zytoplasmas wurden und werden durch die Verwendung eines Elektronenmikroskops hervorgebracht, mit dem man die Details der Struktur der Organellen selbst untersuchen kann. Moderne biophysikalische und biochemische Methoden ermöglichen es, bestimmte Organellen des Zytoplasmas in reiner Form zu isolieren und anschließend zu untersuchen chemische Zusammensetzung und ihre Funktionen. Außerhalb der Zelle, in Umgebungen mit komplexer Zusammensetzung, können viele Organellen die Arbeit erledigen, die sie tun, wenn sie sich in der Zelle befinden.
Der Teil des Zytoplasmas, in den die Organellen eingetaucht sind und der bisher strukturlos zu sein scheint, wird als Hauptsubstanz des Zytoplasmas oder Hyaloplasmas bezeichnet. Hyalo-Plasma ist keineswegs ein passiver Füllstoff, sondern ein aktiv arbeitender Teil des Zytoplasmas. Es finden eine Reihe lebenswichtiger chemischer Prozesse statt, es enthält viele Enzymproteine, mit deren Hilfe diese Prozesse durchgeführt werden.
Das Zellleben besteht aus kontinuierlicher chemischer Arbeit, die in ihrer Gesamtheit als Stoffwechsel bezeichnet wird. Im Wesentlichen ist die Zelle eine Chemiefabrik, die eine breite Palette von Produkten produziert und unabhängig die Energie erzeugt, die zur Herstellung benötigt wird. Ihre Produkte sind Substanzen, die sie selbst benötigt, um ihr eigenes Leben zu erhalten (um ihren Körper während des Wachstums und der Entwicklung aufzubauen, um ihre abgenutzten Teile zu ersetzen) und um Tochterzellen während der Fortpflanzung und für die Bedürfnisse anderer Zellen im Körper zu schaffen.
Alle chemischen Reaktionen, die in der Zelle stattfinden, können in zwei Gruppen unterteilt werden. Infolgedessen zerfallen einige dieser oder andere Substanzen in kleinere Moleküle. Durch andere niedermolekulare Substanzen werden Substanzen mit größeren Molekülen synthetisiert. Ein Molekül einer Substanz besteht aus Atomen, die durch chemische Bindungen zusammengehalten werden, dh durch die eine oder andere Menge konzentrierter potentieller chemischer Energie. Wenn das Molekül aufgebrochen wird, werden die Bindungen aufgebrochen und ihre chemische Energie freigesetzt. Für die Synthese, dh für die Bildung eines größeren Moleküls aus kleinen, ist es notwendig, neue chemische Bindungen herzustellen. Es ist notwendig, einen bestimmten Teil der Energie in diese zu stecken. Im Gegenteil, Zerfallsreaktionen laufen letztendlich mit der Freisetzung von Energie ab, da dabei die Bindungen zwischen Atomen aufgebrochen werden. Die biologische Bedeutung von Zerfallsreaktionen in der Zelle besteht darin, dass sie chemische Energie freisetzen, die dann von der Zelle für Synthesereaktionen und zur Herstellung anderer Arten von Arbeiten (elektrisch, mechanisch, Arbeiten zum Transport von Substanzen) verwendet wird. Da die Zelle während ihres gesamten Lebens verschiedene Substanzen synthetisiert hat, muss sie kontinuierlich andere Substanzen abbauen. Die freigesetzte Energie wird nicht sofort für Synthesen verwendet. Erstens wird es durch die Bildung spezieller Substanzen - Akkumulatoren chemischer Energie - Adenosintriphosphorsäure (ATP) und verwandter Verbindungen gespeichert. Zum richtigen Zeitpunkt und an der richtigen Stelle in der Zelle wird ATP gespalten und gibt Energie für die Synthese der für die Zelle notwendigen Substanz ab.
Alle chemischen Reaktionen in der Zelle - sowohl Synthese als auch Zerfall - werden mit Hilfe von Enzymen durchgeführt. Enzyme sind Proteinsubstanzen, die den Reaktionsverlauf beschleunigen. Diese Beschleunigung ist so groß, dass solche Reaktionen ohne Enzyme in der Zelle überhaupt nicht möglich wären. Es gibt Fälle, in denen die Reaktion dank des Enzyms 1011-mal beschleunigt wird. Dies bedeutet, dass eine Reaktion, die mit der Beteiligung des Enzyms innerhalb von 0,01 Sekunden ohne dieses endet, 31 Jahre andauern würde. Es ist klar, dass solche Reaktionen ohne das Enzym einfach unrealistisch wären.
Zusätzlich wird dank Enzymen der Verlauf chemischer Reaktionen in der Zelle gesteuert und reguliert. Die Aktivität von Enzymen in einer Zelle ändert sich entsprechend ihrem Bedarf an einer bestimmten Substanz oder Energie, dh aufgrund der Arbeit dieser Enzyme. Wenn eine Substanz benötigt wird, werden diese Enzyme, dank denen sie gebildet wird, wieder aktiviert oder synthetisiert. Die Bildung von Substanzen, die Freisetzung und Speicherung von Energie ist das Ergebnis aufeinanderfolgender Reaktionen, das Ergebnis der Arbeit einer ganzen Kette von Enzymen. Alle Strukturen der lebenden Teile der Zelle bestehen aus Enzymen und aus Substanzen, die diese Enzyme zusammenhalten. In diesem Fall befinden sich die Enzyme, die an benachbarten, nacheinander ablaufenden Reaktionen beteiligt sind, nebeneinander. Sie übertragen die Moleküle der Substanzen, die sie umwandeln, wie entlang eines Förderers ineinander, und jeder von ihnen führt seine eigene Arbeitsoperation an dem Molekül aus.
Das Hyaloplasma enthält Enzyme, die Glucosemoleküle in einfachere Brenztraubensäuremoleküle zerlegen. Die in diesem Fall freigesetzte Energie wird durch die Bildung von ATP-Molekülen gespeichert. Der gleiche Prozess findet im Zellkern statt. Der größte Teil der Energie wird jedoch in speziellen Organellen des Zytoplasmas - Mitochondrien - gewonnen, da die Substanzen tiefer abgebaut werden.
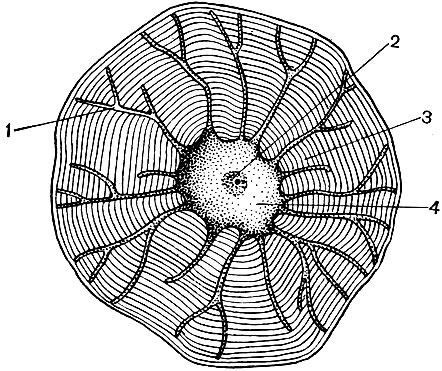
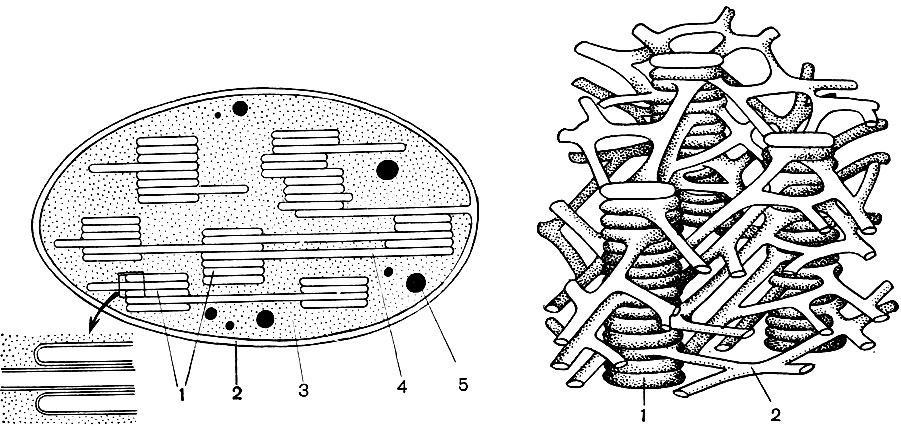
Mitochondrien sind kleine Körper von abgerundeter oder länglicher Form mit einer Größe von 0,5 bis 1,5 Mikrometern, dh der Größe eines Bakteriums. Ihre Anzahl in einer Zelle ist normalerweise groß, ungefähr 100 - 3000. Es gibt jedoch Zellen mit einer kleinen Anzahl von Mitochondrien. Das Sperma der Fucus-Algen enthält also nur 4 Mitochondrien und in den einzelligen Algen-Mikromonen eine. Mitochondrien sind unter einem Lichtmikroskop sichtbar, ihre Feinstruktur kann jedoch nur mit einem Elektronenmikroskop untersucht werden (Tabelle 10, Diagramm der Struktur - Abb. 31). Mitochondrien sind Formationen aus Lipoproteinmembranen, die in eine Grundsubstanz - eine Matrix - eingetaucht sind. Die Mitochondrienmembran besteht aus zwei Membranen, zwischen denen sich eine Lücke befindet.
Die innere Membran der Schale gibt im Inneren zahlreiche Invaginationen, dies sind Kristalle. Es gibt eine Matrix zwischen ihnen. Sowohl die innere Membran der Mitochondrienmembran als auch die von ihr gebildeten Kristalle werden aus der Reihenfolge der lokalisierten Enzyme aufgebaut. Dank der Falten - Kristalle ist die Arbeitsfläche der Membranen in den Mitochondrien sehr groß. Eine Reihe von Enzymen befindet sich in der mitochondrialen Matrix, dh zwischen den Kristallen.
Die Kombination dieser Enzyme führt die intrazelluläre Atmung und die Speicherung der während der Atmung freigesetzten Energie in Form von ATP durch. Die Arbeit der Mitochondrien ist eng mit den Prozessen im Hyaloplasma verbunden, in denen die ersten Phasen des Abbaus von Glukose und anderen Substanzen zu Brenztraubensäure stattfinden. In Mitochondrien findet seine weitere Spaltung statt. Brenztraubensäure dringt in die Mitochondrien ein und wird hier Schritt für Schritt zu Kohlendioxid und Wasser oxidiert, während gleichzeitig Sauerstoff verbraucht wird. Dies ist die intrazelluläre Atmung, bei der die Zelle, die Substanzen spaltet und oxidiert, viel Energie produziert, die sie dann für eine Vielzahl ihrer Bedürfnisse nutzen kann.
Die erste Phase des Abbaus des Glucosemoleküls, in der es in zwei Hälften geteilt wird und die im Hyaloplasma stattfindet, gibt der Zelle nur zwei ATP-Moleküle.
Infolge der zweiten Stufe, die zur vollständigen "Verbrennung" von Glucose führt, werden 36 weitere ATP-Moleküle gebildet. Daher sind Mitochondrien in ihrer Funktion die Kraftwerke der Zelle, Maschinen zur Gewinnung des größten Teils der Energie. Der eigentliche Abbau von Glukose-Abbauprodukten erfolgt in der mitochondrialen Matrix, während ATP aufgrund von Reaktionen gebildet wird, die auf seinen inneren Membranen stattfinden, einschließlich Atemenzymen und Enzymen, die die Bildung von ATP sicherstellen. Die Anzahl der Kristalle in Mitochondrien kann variieren. Je mehr von ihnen, desto höher ist die biochemische Aktivität der Mitochondrien.
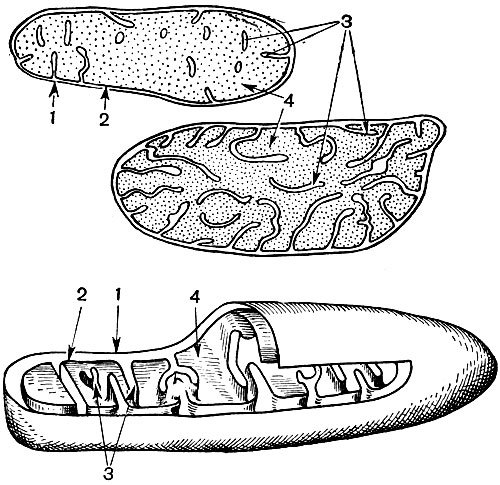
Wir sprechen hier von Glukose als Substanz, deren Abbau die Zelle Energie produziert. Glukose ist zentral, aber nicht die einzige solche Substanz. Sein Molekül hat ein Rückgrat aus sechs miteinander verbundenen Kohlenstoffatomen. Infolge einer langen und komplexen Reaktionskette wird sein Molekül zerkleinert, oxidiert und am Ende in sechs Moleküle einer anorganischen Substanz - Kohlendioxid (CO 2) - aufgeteilt, von denen jedes Molekül nur ein Kohlenstoffatom enthält, und es wird extrem oxidiert. Indem Sie alle aufeinanderfolgenden Reaktionen der Glucoseoxidation addieren und alle Zwischenprodukte ausschließen, können Sie die Gesamtreaktion dieses Prozesses erhalten:
Stärke wird leicht in Glucose umgewandelt, wonach sie den oben beschriebenen Abbau erfährt. Proteine \u200b\u200bund Fette ergeben verschiedene organische Säuren, die in Zwischenprodukte des Glukoseabbaus umgewandelt und dann auf die gleiche Weise wie letztere und mit Hilfe der gleichen Enzyme oxidiert werden.
Die vollständige biologische Oxidation organischer Stoffe ähnelt ihrer Verbrennung. In beiden Fällen werden Kohlendioxid, Wasser und Energie freigesetzt. Während der Verbrennung wird diese Energie jedoch in Form von Wärme und vollständig sofort freigesetzt; Während der biologischen Oxidation wird die Energie chemischer Bindungen in Teilen freigesetzt, und ihr Hauptteil wird gebunden und geht in die Energie der chemischen Phosphatbindung von ATP über. Infolgedessen erhält die Zelle ein Energiekonzentrat in einer solchen Form, das dann zum richtigen Zeitpunkt und an der richtigen Stelle verwendet werden kann, um neue chemische Bindungen herzustellen, neue Substanzen zu synthetisieren und andere Arten von Arbeit zu produzieren - elektrische, mechanische sowie Arbeiten zum Transport von Substanzen von Umgebung zu Zelle, von Zelle zu Umgebung und von Zelle zu Zelle.
Je aktiver die Vitalaktivität der Zelle ist, desto größer ist ihr Energiebedarf und desto mehr Mitochondrien enthält sie. Sie können ungleichmäßig in derselben Zelle verteilt sein: Es gibt mehr davon in dem Teil der Zelle, der derzeit aktiver arbeitet.
Mitochondrien sind in der Lage, einige der Substanzen zu synthetisieren, aus denen sie selbst bestehen. Dank dessen können sich Mitochondrien vermehren.
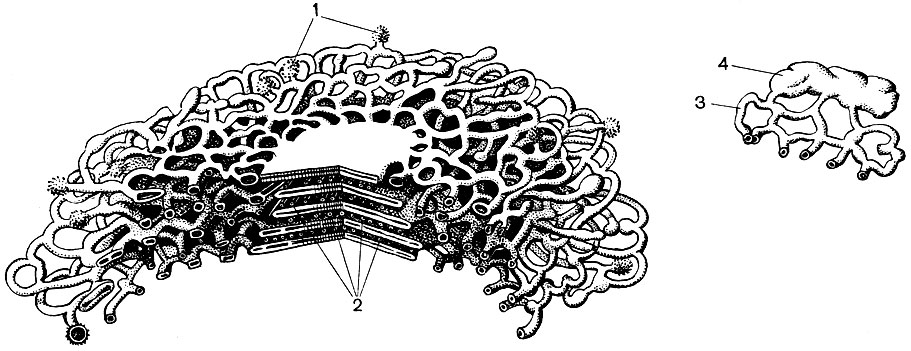
Das endoplasmatische Retikulum ist ein Organoid des Zytoplasmas, in dem sehr viele Substanzen synthetisiert werden (Tabelle 10). Das endoplasmatische Retikulum ist ein System von Kanälen, die das Zytoplasma durchdringen und sich in einigen Bereichen verengen, in anderen ausdehnen und entweder Zisternen oder flache Beutel oder Verzweigungsröhrchen bilden. Die Wände all dieser Formationen bestehen aus Membranen, die Enzyme enthalten.
Wie bei anderen Membranformationen der Zelle sind die Enzyme im Retikulum geordnet angeordnet. In diesem Fall führen benachbarte Enzyme sequentiell ablaufende Reaktionen (Arbeitsvorgänge) durch, und ihre Gruppe führt die gesamte Reaktionskette aus, die zur Bildung einer bestimmten Substanz führt.
Unterscheiden Sie zwischen agranularem (glattem) und körnigem endoplasmatischem Retikulum. Auf der äußeren Oberfläche der Kanäle des körnigen Retikulums befinden sich zahlreiche kleine Organellen - Ribosomen, deren Funktion die Synthese von Proteinmolekülen ist. Das agranuläre endoplasmatische Retikulum, das quantitativ das körnige in der Pflanzenzelle überwiegt, trägt keine Ribosomen.
Das endoplasmatische Retikulum ist nicht nur ein Förderer für viele Arten der enzymatischen Umwandlung von Substanzen, hauptsächlich für deren Synthese, sondern auch ein System von Autobahnen, auf denen sich Substanzen durch die Zelle bewegen. Das Retikulum beginnt an der äußeren Membran der Kernmembran und nähert sich verzweigt verschiedenen Organellen des Zytoplasmas sowie dem Plasmalemma. So verbindet er alle Teile der Zelle miteinander. Zusätzlich passieren die Kanäle des endoplasmatischen Retikulums die Plasmodesmen und verbinden das Retikulum benachbarter Zellen.
Ferner zerlegen die Membranen des endoplasmatischen Retikulums das Zytoplasma in zahlreiche Kompartimente, aufgrund derer die Zelle nicht als homogene Masse dargestellt werden kann, in die eine Vielzahl von Substanzen eingemischt sind. Die Bedingungen in einem Fach können völlig anders sein als in jedem anderen. Die darin ablaufenden Prozesse können nur darin stattfinden, während ineinander unterschiedliche Prozesse stattfinden.
Schließlich sind die Membranen des endoplasmatischen Retikulums diejenigen Oberflächen, auf denen sich Bioströme ausbreiten. Dies sind Signale, die die selektive Permeabilität von Membranen und damit die Aktivität von Enzymen verändern. Dadurch werden einige chemische Reaktionen in Gang gesetzt, andere gehemmt - der Stoffwechsel unterliegt einer Regulierung und verläuft koordiniert.
Viele der in der Zelle synthetisierten Substanzen müssen konzentriert und aus der Zelle entweder in die äußere Umgebung oder in die intrazelluläre Vakuole freigesetzt werden. Darüber hinaus konzentriert eine Zelle auch Substanzen, die von anderen Zellen in sie gelangen, beispielsweise wenn sie diese in Reserve speichert. Diese Arbeit wird von Diktyosomen erledigt. Normalerweise gibt es in einer Pflanzenzelle mehrere Dictyosomen, und ihr gesamter Satz wird als Golge-Apparat (oder Komplex) und die gegebene Zelle bezeichnet. Jedes Dictyosom ist ein System gestapelter Membranen (Tabelle 10 - Abschnitt, Abb. 32 - Volumendiagramm). Die Hohlräume zwischen den Membranen, die dieses Organoid bilden, sehen aus wie schmale Schlitze, dann flache Säcke - Zisternen, dann Blasen. Ihre Form ändert sich während des Betriebs des Organoids und hängt anscheinend vom Füllungsgrad der Zwischenmembranräume mit den sekretierten und angesammelten Substanzen ab. Die gebildeten und überwucherten Vesikel sind vom Organoid getrennt. Anscheinend sind viele zelluläre Vakuolen, die von Membranen umgeben sind - Tonoplasten - ein Produkt der Aktivität des Golgi-Apparats; Dies sind Blasen, die sich davon lösen und dann vergrößert werden.
Der Golgi-Apparat wurde speziell in Ausscheidungszellen (Sekretionszellen) entwickelt, in denen verschiedene Substanzen abgelagert sind oder aus denen verschiedene Substanzen ausgeschieden werden. Es synthetisiert und setzt Substanzen frei, die die Zellmembran bilden.
Lysosomen sind eher kleine (etwa 0,5 Mikrometer im Durchmesser) gerundete Körper - ein weiteres Organoid des Zytoplasmas. Sie sind mit einer Membran bedeckt - einer Lipoproteinmembran. Der Inhalt von Lysosomen sind Enzyme, die Proteine, Kohlenhydrate, Nukleinsäuren und Lipide verdauen. Die Lysosomenmembran verhindert die Freisetzung von Enzymen aus dem Organoid in das Hyaloplasma, da diese sonst von diesen Enzymen verdaut würden.
Es kann angenommen werden, dass Lysosomen Produkte der Aktivität des Golgi-Apparats sind, von denen sich Blasen lösen, in denen dieses Organoid verdauende Enzyme angesammelt hat.
Die Teile der Zelle, die während ihrer Entwicklung absterben, werden durch lysosomale Enzyme zerstört. In einer verstorbenen Zelle werden Lysosomen zerstört, Enzyme im Zytoplasma gefunden und die gesamte Zelle mit Ausnahme der Membran wird verdaut.
Ribosomen sind sehr kleine Organellen mit einem Durchmesser von etwa 250 A. Sie sind fast kugelförmig. Einige von ihnen sind an den äußeren (hyaloplasmatischen) Oberflächen der Membranen angebracht, die die Kanäle des körnigen endoplasmatischen Retikulums bilden; Einige befinden sich im Hyaloplasma in einem freien Zustand. Eine Zelle kann bis zu 5 Millionen Ribosomen enthalten. Sie sind "Apparate für die Proteinsynthese". Daher sind sie besonders zahlreich in Zellen, die aktiv Protein bilden - in wachsenden Zellen, in Zellen, die Proteinsubstanzen absondern. Ribosomen kommen auch in Mitochondrien und Chloroplasten vor, wo sie einige der Proteine \u200b\u200bsynthetisieren, aus denen diese aufgebaut sind. Organellen.
Organellen, sogenannte Mikrotubuli, kommen in vielen Zellen vor. Ihr Name spricht von ihrer Form - dies sind Röhren mit einem Kanal im Inneren. Ihr Außendurchmesser beträgt ca. 250A. Manchmal handelt es sich um Doppelrohre - zwei einzelne, die nebeneinander liegen und eine gemeinsame Wand haben, die ihre Hohlräume trennt. Die Wände von Mikrotubuli bestehen aus Proteinmolekülen. Es wird angenommen, dass Mikrotubuli mit der kontraktilen (motorischen) Aktivität des Zytoplasmas und seiner Formationen verbunden sind. Aus ihnen wie aus Bauteilen werden offenbar die kontraktilen Strukturen des Flagellums, eines Organoids, mit dessen Hilfe sich einige einzellige und koloniale Algen bewegen, sowie Zellen, die der Fortpflanzung vieler niederer Pflanzen dienen, aufgebaut. Während der Zellteilung werden Spindelfäden aus Mikrotubuli gebildet, auf die weiter eingegangen wird. Während der Teilungsperiode sammeln sich Mikrotubuli in Gruppen und bilden diese Filamente. Am Ende der Teilung zerfallen die Filamente wieder in separate Mikrotubuli. In Zellen oder Teilen davon, denen eine dichte Membran fehlt, erfüllen Mikrotubuli möglicherweise eine unterstützende Funktion und bilden das innere Skelett der Zelle.
Plastiden sind Organellen, die nur Pflanzenzellen innewohnen. In der Regel handelt es sich dabei um große Körper, die unter einem Lichtmikroskop deutlich sichtbar sind.
Es gibt 3 Arten von Plastiden: farblos - Leukoplasten, grün - Chloroplasten, in anderen Farben gefärbt - Chromoplasten. Jeder Plastidentyp hat seine eigene Struktur und seine eigenen Funktionen. Übergänge von Plastiden von einem Typ zum anderen sind jedoch möglich. So wird die Begrünung von Kartoffelknollen durch die Umlagerung ihrer Leukoplasten in Chloroplasten verursacht. In der Karottenwurzelernte gehen die Leukoplasten in Chromoplasten über. Alle drei Arten von Plastiden werden aus Proplastiden gebildet.
Proplastiden sind farblose Körper, ähnlich wie Mitochondrien, aber etwas größer als sie. Sie kommen in großen Mengen in meristematischen Zellen vor. Leukoplasten kommen in den Zellen ungefärbter Pflanzenteile (Früchte, Samen, Wurzeln, Blattepidermis) vor. Ihre Form ist unbestimmt. Am häufigsten gibt es Leukoplasten, in denen Stärke abgelagert wird (sie wird aus Zucker gebildet). Es gibt Leukoplasten, die Proteine \u200b\u200bspeichern. Am seltensten sind fettgefüllte Leukoplasten; Sie entstehen während der Alterung der Chloroplasten. Es gibt keine signifikanten Unterschiede zwischen Leukoplasten und Proplastiden.
Chloroplasten sind Plastiden höherer Pflanzen, in denen der Prozess der Photosynthese stattfindet, d. H. Die Nutzung der Energie von Lichtstrahlen zur Bildung organischer Substanzen aus anorganischen Substanzen (Kohlendioxid und Wasser) bei gleichzeitiger Freisetzung von Sauerstoff in die Atmosphäre. Chloroplasten haben die Form einer bikonvexen Linse, ihre Größe beträgt etwa 4 - 6 mk. Sie kommen in den Narenchymzellen von Blättern und anderen grünen Teilen höherer Pflanzen vor. Ihre Anzahl in einer Zelle variiert zwischen 25 und 30.
Die Struktur des Chloroplasten ist in den Tabellen 11 und 12 (Schnittansicht) und im Diagramm in Abbildung 33 angegeben. Draußen ist der Chloroplast mit einer Membran bedeckt, die aus zwei Lipoproteinmembranen besteht. Darunter sind in der Hauptsubstanz (Stroma) zahlreiche Formationen - Lamellen - angeordnet. Sie bilden flache Beutel, die in normalen Ständen übereinander liegen. Diese Stoiker, die in einer Säule gestapelten Münzen ähneln, werden Körner genannt. Längere Lamellen passieren sie, so dass alle Chloroplastenkörner zu einem einzigen System verbunden sind. Die Zusammensetzung der Membranen, die das Granulat bilden, enthält ein grünes Pigment - Chlorophyll. Hier finden die Lichtreaktionen der Photosynthese statt - die Absorption von Lichtstrahlen durch Chlorophyll und die Umwandlung von Lichtenergie in die Energie angeregter Elektronen. Durch Licht angeregte Elektronen, dh mit überschüssiger Energie, geben Energie ab, um Wasser zu zersetzen und ATP zu synthetisieren. Bei der Zersetzung von Wasser entstehen Sauerstoff und Wasserstoff. Sauerstoff wird in die Atmosphäre freigesetzt und Wasserstoff wird durch das Protein Ferredoxin gebunden. Ferredoxin wird dann erneut oxidiert, wobei dieser Wasserstoff an ein Reduktionsmittel abgegeben wird, das als NADP abgekürzt wird. NADP geht in die reduzierte Form - NADP-H 2. Das Ergebnis von Lichtreaktionen der Photosynthese ist somit die Bildung von ATP, NADP-H 2 und Sauerstoff, und Wasser und Lichtenergie werden verbraucht.
ATP sammelt viel Energie - es wird dann für Synthesen sowie für andere Bedürfnisse der Zelle verwendet. NADP-H 2 ist ein Wasserstoffspeicher und gibt ihn dann leicht ab. Daher ist NADP-H 2 ein chemisches Reduktionsmittel. Eine große Anzahl von Biosynthesen ist mit einer Reduktion verbunden, und NADPH 2 fungiert bei diesen Reaktionen als Wasserstofflieferant.
Darüber hinaus treten mit Hilfe von Chloroplasten-Stromaenzymen, dh außerhalb des Granulats, dunkle Reaktionen auf: Wasserstoff und die in ATP enthaltene Energie werden verwendet, um atmosphärisches Kohlendioxid (CO2) zu reduzieren und in organische Substanzen einzubauen. Die erste organische Substanz, die durch Photosynthese gebildet wird, unterliegt einer Vielzahl von Umlagerungen und führt zu einer Vielzahl von organischen Substanzen, die in einer Pflanze synthetisiert werden und ihren Körper bilden. Eine Reihe dieser Transformationen findet genau dort im Chloroplasten-Stroma statt, wo es Enzyme für die Bildung von Zuckern, Fetten sowie alles gibt, was für die Proteinsynthese notwendig ist. Die Zucker können sich dann entweder vom Chloroplasten zu anderen Zellstrukturen und von dort zu anderen Pflanzenzellen bewegen oder Stärke bilden, deren Körner häufig in Chloroplasten zu sehen sind. Fette werden auch in Chloroplasten entweder in Form von Tropfen oder in Form von einfacheren Substanzen, Fettvorläufern, abgelagert und verlassen den Chloroplasten.
Die Komplikation von Substanzen ist mit der Bildung neuer chemischer Bindungen verbunden und erfordert normalerweise Energie. Seine Quelle ist die gleiche Photosynthese. Tatsache ist, dass sich ein erheblicher Teil der durch die Photosynthese gebildeten Substanzen im Hyaloplasma und in den Mitochondrien wieder zersetzt (bei vollständiger Verbrennung zu Substanzen, die als Ausgangsmaterial für die Photosynthese dienen - CO2 und H2O). Infolge dieses Prozesses, der im Wesentlichen die Umkehrung der Photosynthese darstellt, wird die zuvor in den chemischen Bindungen zersetzter Substanzen akkumulierte Energie freigesetzt und - wiederum mithilfe von ATP - für die Bildung neuer chemischer Bindungen der synthetisierten Moleküle aufgewendet. Ein wesentlicher Teil der Produktion der Photosynthese wird daher nur benötigt, um Lichtenergie zu binden und nach ihrer Umwandlung in eine Chemikalie zur Synthese völlig unterschiedlicher Substanzen zu verwenden.
Und nur ein Teil der bei der Photosynthese gebildeten organischen Substanz wird als Baumaterial für diese Synthesen verwendet.
Die Produktion von Photosynthese (Biomasse) ist kolossal. Für ein Jahr auf der Welt sind es ungefähr 1010 Tonnen. Organische Substanzen, die von Pflanzen erzeugt werden, sind die einzige Lebensquelle nicht nur für Pflanzen, sondern auch für Tiere, da letztere fertige organische Substanzen verarbeiten und sich entweder direkt von Pflanzen oder anderen Tieren ernähren, die Sie ernähren sich wiederum von Pflanzen. Somit ist die Photosynthese das Herzstück allen modernen Lebens auf der Erde. Alle Transformationen von Substanzen und Energie in Pflanzen und Tieren sind Umlagerungen, Rekombinationen und Transfers von Materie und Energie der Primärprodukte der Photosynthese. Die Photosynthese ist wichtig für alle Lebewesen und die Tatsache, dass eines ihrer Produkte freier Sauerstoff ist, der aus einem Wassermolekül stammt und in die Atmosphäre freigesetzt wird. Es wird angenommen, dass der gesamte Sauerstoff in der Atmosphäre aus der Photosynthese stammt. Es ist wichtig für die Atmung von Pflanzen und Tieren.
Chloroplasten können sich in der Zelle bewegen. Bei schlechten Lichtverhältnissen befinden sie sich unter der Zellwand, die dem Licht zugewandt ist. Dabei werden sie mit ihrer größeren Oberfläche zum Licht. Wenn das Licht zu intensiv ist, drehen sie sich zu ihm um und richten sich entlang der Schatten parallel zu den Lichtstrahlen aus. Bei mittlerer Beleuchtung nehmen Chloroplasten eine Position zwischen den beiden extremen ein. In jedem Fall wird ein Ergebnis erzielt: Chloroplasten sind unter den günstigsten Lichtbedingungen für die Photosynthese. Eine solche Bewegung von Chloroplasten (Phototaxis) ist eine Manifestation einer der Arten von Reizbarkeit in Pflanzen.
Chloroplasten haben eine gewisse Autonomie im Zellsystem. Sie haben ihre eigenen Ribosomen und eine Reihe von Substanzen, die die Synthese einer Reihe von Chloroplasten-eigenen Proteinen bestimmen. Es gibt auch Enzyme, deren Arbeit zur Bildung von Lipiden führt, aus denen die Lamellen bestehen, und Chlorophyll. Wie wir gesehen haben, verfügt Chloroplasten auch über ein autonomes Energieerzeugungssystem. Dank alledem können Chloroplasten selbstständig ihre eigenen Strukturen aufbauen. Es gibt sogar die Ansicht, dass Chloroplasten (wie Mitochondrien) von einigen niederen Organismen stammten, die sich in einer Pflanzenzelle niederließen und zuerst eine Symbiose mit ihr eingingen und dann zu ihrem integralen Bestandteil, einem Organoid, wurden.
In niederen Pflanzen wird die Photosynthese auch von spezialisierten, wenn auch nicht so hoch entwickelten Membranstrukturen wie in Chloroplasten durchgeführt. In photosynthetischen Bakterien bilden Membranen, die Chlorophyll enthalten, ein Netzwerk, das den Bakterienkörper durchdringt. In Blaualgen werden photosynthetische Membranen zu flachen Blasen verschmolzen. Bei Grün- und anderen Algen ist das System dieser Membranen durch eine Deckmembran vom Rest der Zelle getrennt und bildet ein spezielles Organoid - ein Chromatophor. Die Anzahl der Chromatophore in einer Zelle ist gering, häufig enthält eine Zelle nur ein Chromatophor. Ihre Form ist bei Algen verschiedener Arten sehr unterschiedlich.
In Spirogyra hat das Chromatophor die Form eines Bandes, das sich entlang der Zellwände windet; in Clostridium sind dies gerippte Zylinder; Das Zygnema hat Sternkörper.
Chromoplasten entstehen entweder aus Proplastiden oder aus Chloroplasten oder aus Leukoplasten. Ihre innere Membranstruktur ist viel einfacher als die von Chloroplasten. Gran ist nicht, das Stroma enthält viel gelbes oder orangefarbenes Pigment. Chromoplasten kommen in den Zellen von Blütenblättern, Früchten und Wurzelfrüchten vor.
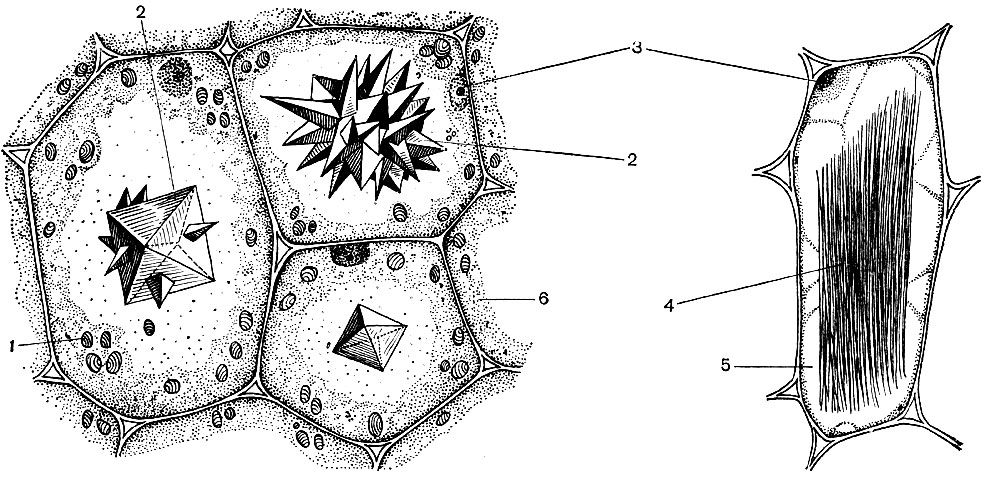
Eine typische Pflanzenzelle hat eine große Vakuole, die mit flüssigem Inhalt gefüllt ist. Oft nimmt die Vakuole fast das gesamte Volumen der Zelle ein, so dass das Zytoplasma nur eine dünne Schicht neben der Zellmembran ist. Junge Zellen haben mehrere kleine Vakuolen, die bei der Entwicklung der Zellen wachsen und zu einer verschmelzen. Der Gehalt der Vakuole - Zellsaft - ist eine wässrige Lösung sehr vieler Substanzen: Zucker, Aminosäuren, andere organische Säuren, Pigmente (Farbstoffe), Vitamine, Tannine, Alkaloide, Glykoside, anorganische Salze (Nitrate, Phosphate, Chloride) und manchmal Proteine.
Alle diese Substanzen sind die Abfallprodukte der Zelle. Einige von ihnen werden in vakuolarem (Zell-) Saft als Reservesubstanzen gelagert und gelangen im Laufe der Zeit zur Verwendung wieder in das Zytoplasma. Andere sind Stoffwechselabfälle, die aus dem Zytoplasma entfernt werden. So wird Oxalsäure in die Vakuole entfernt; Calciumoxalatkristalle werden häufig in vakuolarem Saft abgelagert - manchmal in Form von Einkristallen, in anderen Fällen in Form eines Konglomerats von Kristallen dieses Salzes - vielfältig (Drusen) oder nadelartig (Raffida), wie in Abbildung 34 gezeigt.
Die überwiegende Mehrheit der Pflanzen (mit Ausnahme von prokaryotischen Organismen) hat in jeder lebenden Zelle einen Kern oder mehrere Kerne. Eine kernlose Zelle kann nur kurze Zeit leben. Anukleierte Zellen von Siebrohren sind lebende Zellen. Aber sie leben nicht lange. In allen anderen Fällen sind die nichtnuklearen Zellen tot.

Der Kern liegt immer im Zytoplasma. Die Form des Kerns kann unterschiedlich sein - rund, oval, stark länglich, unregelmäßig mehrlappig. In einigen Zellen ändern sich die Konturen des Kerns während seiner Funktion, und auf seiner Oberfläche bilden sich Lappen unterschiedlicher Größe. Die Größen der Kerne sind in den Zellen verschiedener Pflanzen und in verschiedenen Zellen derselben Pflanze nicht gleich. Relativ große Kerne finden sich in jungen meristematischen Zellen, in denen sie bis zu 3/4 des Volumens der gesamten Zelle einnehmen können. Die relativen und manchmal absoluten Größen der Kerne in entwickelten Zellen sind viel kleiner als in jungen.
Draußen ist der Kern mit einer Hülle bedeckt, die aus zwei Membranen besteht, zwischen denen sich eine Lücke befindet - der perinukleare Raum. Die Schale ist durch Poren unterbrochen. Die äußere der beiden Membranen der Hülle gibt Auswüchse ab, die direkt in die Wände des endoplasmatischen Retikulums des Zytoplasmas gelangen. Sowohl die Poren als auch die direkte Verbindung des endoplasmatischen Retikulums mit dem perinukleären Raum stellen einen engen Kontakt zwischen dem Kern und dem Zytoplasma her.
Der Inhalt des Kerns ist eine körnige Grundsubstanz (Kernsaft oder Nucleoplasma), in die dichtere Strukturen eingebracht werden - Chromosomen und der Nucleolus. Der Nucleolus ist der Apparat zur Synthese des Materials der Ribosomen und des Ortes ihrer Anordnung aus diesem Material.
Chromosomen bestehen aus einer großen Anzahl von Desoxyribonukleinsäuremolekülen (DNA), die an Histonproteinmoleküle gebunden sind. DNA-Moleküle sind lange, komplex gepackte Doppelstränge. Jedes Molekül besteht aus zwei Strängen, die spiralförmig umeinander gedreht sind. Ein Faden ist wiederum eine Kette einer großen Anzahl sogenannter Nukleotide. Nucleotid ist eine Verbindung aus stickstoffhaltiger Base, Kohlenhydraten (Desoxyribose) und Phosphorsäure.
Jedes der Nukleotide enthält eine der folgenden vier stickstoffhaltigen Basen: Adenin, Guanin, Cytosin oder Thymin. Dementsprechend werden in der DNA 4 verschiedene Nukleotide unterschieden: Adenin (A), Guanin (G), Cytosin (C) und Thymin (T). Die Nukleotide sind über ihre Phosphatgruppen zu einer langen Kette miteinander verbunden. Zwei umeinander verdrillte Ketten, die ein DNA-Molekül bilden, sind durch chemische Wechselwirkung (die sogenannten Wasserstoffbrücken) der stickstoffhaltigen Basen ihrer Nukleotide miteinander verbunden. Die Basen bilden Paare - eine Base aus einer Kette, die andere aus der zweiten. Das Diagramm in Abbildung 35 gibt eine Vorstellung von der Reihenfolge, in der die Nukleotide zu einer Kette verbunden sind, und den beiden Ketten - untereinander.
Obwohl alle Desoxyribonukleinsäuremoleküle gemäß dem beschriebenen Einzelplan aufgebaut sind, ist ihre spezifische qualitative Zusammensetzung unterschiedlich; Sie unterscheiden sich auch in der Größe der Moleküle. Ein DNA-Molekül enthält bis zu 50 - 100.000 Basenpaare, aber die Anzahl der Paare in jedem Molekül unterscheidet sich von der Anzahl der Nukleotidpaare in jedem anderen Molekül. Zusätzlich gibt es 4 verschiedene Arten von Nukleotiden, und jedes DNA-Molekül hat seinen eigenen inhärenten Prozentsatz an Nukleotiden jedes gegebenen Typs. Mit anderen Worten ist das quantitative Verhältnis A: G: C: T für jedes DNA-Molekül unterschiedlich. Und schließlich ist in jedem DNA-Molekül die Reihenfolge des Wechsels der Nukleotide A, G, C, T nur für dieses Molekül charakteristisch. Die Permutation von nur zwei Paaren von 50.000 Basenpaaren, selbst wenn sie benachbart sind, verändert die Eigenschaften des gesamten Moleküls dramatisch. das gleiche, indem mindestens ein Paar ersetzt wird, sagen wir Von - G nach A - T oder G - Iz; das gleiche - das Fehlen eines Paares unter den gleichen 50.000. In Wirklichkeit können sich DNA-Moleküle nicht in einer Verbindung, nicht in einem oder zwei Nukleotidpaaren, sondern in einer großen Anzahl von ihnen gleichzeitig voneinander unterscheiden. Die Anzahl möglicher Permutationen von Nukleotidpaaren in einem DNA-Molekül ist unendlich, und dementsprechend ist die Anzahl verschiedener Moleküle unendlich, von denen jedes seine eigenen Eigenschaften hat. Die Sequenz von Nukleotiden in einem DNA-Molekül ist eine verschlüsselte Aufzeichnung der Zusammensetzung eines bestimmten Proteins, die für eine bestimmte Zelle charakteristisch ist. In jedem DNA-Molekül werden Aufzeichnungen über die Zusammensetzung mehrerer Proteine \u200b\u200bnacheinander und in allen DNA-Molekülen aller Chromosomen des Kerns angepasst - Aufzeichnungen über die Zusammensetzung aller Proteine \u200b\u200beiner Zelle, die während ihres Lebens darin synthetisiert werden können. Das Wesentliche dieser codierten Datensätze ist wie folgt.
Die Moleküle jedes Proteins sind Ketten von Aminosäuren, die in Reihe geschaltet sind. Es gibt ungefähr 20 verschiedene Aminosäuren, und die Natur eines Proteins wird dadurch bestimmt, aus welchen Aminosäuren sein Molekül besteht, wie viel sie insgesamt im Molekül enthalten und in welcher Reihenfolge sie miteinander verbunden sind. Beispielsweise ist ein DNA-Abschnitt, der für die Zusammensetzung eines bestimmten Proteins verantwortlich ist (jeder dieser Abschnitte wird als Gen bezeichnet), eine Aufzeichnung einer bestimmten Sequenz von Aminosäuren, die ein Molekül eines bestimmten Proteins bilden, deren Gesamtzahl darin. Alle 3 aufeinanderfolgenden Nukleotide der DNA-Kette bezeichnen (codieren) eine Aminosäure des entsprechenden Proteinmoleküls. Die nächsten 3 Nukleotide codieren die nächste Aminosäure und so weiter. Die Anzahl möglicher unterschiedlicher Kombinationen von 3 Nukleotiden aus vier verschiedenen Typen ist mehr als ausreichend, um 20 Aminosäuren zu codieren. Es ist interessant, dass dieselben Aminosäuren in der ganzen lebenden Welt von denselben Nukleotidkombinationen codiert werden. Infolgedessen codiert in einem Gen seine Nukleotidzusammensetzung die Aminosäurezusammensetzung aller Proteine, die in einer bestimmten Zelle synthetisiert werden können.
Zelluläre Proteine \u200b\u200bsind Enzyme. Enzyme bestimmen den Verlauf aller Reaktionen, die die Essenz des Lebens von Zellen ausmachen. Die Bildung und Umwandlung aller anderen Substanzen der Zelle hängt von der Anwesenheit bestimmter Enzyme ab, sei es Fette, Kohlenhydrate, Alkaloide, Harze usw. Dies bildet wiederum alle Eigenschaften der Zelle, die sie von den Zellen anderer Pflanzen in den Zellkernen unterscheiden welches die Zusammensetzung eines anderen Satzes von Enzymen codierte. Deshalb sind Informationen über die Zusammensetzung aller Proteine, die in einer Zelle gebildet werden können, Informationen über alle Eigenschaften einer Zelle und eines Organismus. Weiter werden wir sehen, dass diese Informationen erblich sind, das heißt, dass sie während ihrer Reproduktion vollständig von Zelle zu Zelle und von der Mutterpflanze zu den Tochterpflanzen übertragen werden.
Diese Informationen werden in Chromosomen gespeichert. Die Realisierung dieser Informationen - die Synthese von Proteinen - findet in ihnen jedoch nicht statt. Chromosomengene wirken nur als Initiatoren dieser Synthese. Wenn eine Zelle ein bestimmtes Protein bilden muss, wird das Gen aktiviert, in dem die Zusammensetzung dieses Proteins kodiert ist. Dies bedeutet, dass Moleküle der sogenannten Informationsribonukleinsäure (i-RNA) auf dem Abschnitt des DNA-Strangs gebildet werden, aus dem dieses Gen besteht.
Die Struktur und Zusammensetzung dieser Moleküle, die einzelne Ketten von Nukleotiden sind, spiegeln die Nukleotidstruktur des Gens wider, auf dem sie gebildet wurden. Somit werden die Informationen über die Zusammensetzung des zukünftigen Proteins kopiert.
Die Bildung von m-RNA-Molekülen (viele von ihnen werden auf jedem Gen gebildet) bedeutet auch die Vermehrung von Informationen, sozusagen das Einprägen vieler identischer Matrizen, die die Struktur desselben Gens widerspiegeln und dadurch Informationen über die Aminosäurezusammensetzung eines bestimmten Proteins in der Nukleotidsequenz ihrer Moleküle enthalten. Diese Gendrucke gelangen vom Zellkern zum Zytoplasma. Hier wird die in den Matrizen - i-RNA-Molekülen enthaltene Information entschlüsselt, die Information wird realisiert, sie wird aus der Sprache der Nukleotidsequenz in die Sprache der Aminosäuresequenz übersetzt, d. H. Die Synthese von Proteinmolekülen einer gegebenen Zusammensetzung.
An der Dekodierung und Synthese sind neben dem i-RNA-Molekül eine große Anzahl von Molekülen verschiedener Transport-Ribonukleinsäuren (t-RNA), Ribosomen und eine Reihe von Enzymen beteiligt. Aminosäuren binden Molekül für Molekül an t-RNA. Jede der 20 Arten von Aminosäuren hat ihre eigene t-RNA. Das t-RNA-Molekül hat also chemische Gruppen, die seine Aminosäure erkennen und aus allen verfügbaren Aminosäuren auswählen können. Dies geschieht mit Hilfe spezieller Enzyme. Nachdem t-RNA seine Aminosäure erkannt hat, geht sie eine Verbindung mit ihr ein. Das Ribosom ist an den Anfang des i-RNA-Moleküls gebunden, das sich entlang der i-RNA in einer Polypeptidkette genau mit den Aminosäuren verbindet, deren Reihenfolge durch die Nukleotidsequenz der gegebenen i-RNA verschlüsselt wird. Dieses Lesen von Informationen, diese Dekodierung von Informationen erfolgt aufgrund eines speziellen Mechanismus, der in der Transport-RNA verfügbar ist, deren Moleküle genau die Aminosäuren ausrichten, die durch die Nukleotidsequenz des i-RNA-Moleküls "bezeichnet", "geschrieben" werden. Das Ribosom ist eine Vorrichtung zum chemischen Binden von Aminosäuren in ein Proteinmolekül.
Das allgemeine Schema ist also wie folgt. Auf dem aktivierten Gen - einem Abschnitt einer der Ketten eines DNA-Moleküls - werden Nukleotidketten, i-RNA-Moleküle, synthetisiert, deren Zusammensetzung die Zusammensetzung des Gens genau widerspiegelt und daher eine Aufzeichnung der Zusammensetzung des von diesem Gen codierten Proteins enthält. Im Zytoplasma des Ribosoms werden auf der Grundlage von Informationen, die von den m-RNA-Molekülen unter Verwendung von Transport-RNAs aus dem Gen übertragen werden, verschiedene Aminosäuren in einer bestimmten Sequenz kombiniert, was zur Bildung eines Moleküls des Proteins führt, dessen Zusammensetzung in diesem Gen kodiert ist.
Die Bildung von i-RNA-Molekülen auf dem aktivierten Gen dient sowohl als Befehl als auch als spezifisches Programm für die Synthese eines streng definierten Proteins. Das gleiche mRNA-Molekül wird als Blaupause verwendet, um viele der gleichen Proteinmoleküle zu erzeugen. Es ist jedoch eher kurzlebig, daher ist für eine Langzeitsynthese neuer Moleküle desselben Proteins die Bildung identischer, aber neuer Kopien von m-RNA-Molekülen auf demselben Gen erforderlich. Mit dem Übergang eines Gens in einen inaktiven Zustand - und dies geschieht, wenn der Bedarf der Zelle an einem bestimmten Protein verschwindet - wird es blockiert, hört auf, i-RNA zu bilden, und bald hört die Synthese dieses Proteins auf. Während des Lebens einer Zelle benötigt sie verschiedene Proteine. Jedes Mal werden die Gene, die sie definieren, aktiviert und die entsprechende Zusammensetzung der i-RNA-Moleküle gebildet.
Jedes Ribosom produziert während seines Lebens viele Moleküle verschiedener Proteine. Es kann auf der Basis jeder i-RNA arbeiten, und das Ergebnis, die Art des Proteins, das es erzeugt hat, hängt nur von der Zusammensetzung der i-RNA ab, mit der das Ribosom diesmal in Kontakt stand.
Somit führt der Zellkern die folgenden miteinander verbundenen Funktionen aus. Es speichert Informationen über die Zusammensetzung aller Proteine, die in einer bestimmten Zelle während ihres Lebens synthetisiert werden können. (Die Ausnahme bilden einige Proteine \u200b\u200bvon Mitochondrien und Chloroplasten. Ihre Zusammensetzung ist in ihrer eigenen DNA dieser Organellen kodiert, in der sich diese DNAs befinden. Hier finden vor Ort alle Stadien der Dekodierung der in diesen DNAs enthaltenen Informationen statt, einschließlich der Synthese dieser Proteine \u200b\u200bunter Verwendung eigene Ribosomen.)
Der Kern speichert Informationen über alle Eigenschaften der Zelle und des Organismus. Der Kern organisiert die Synthese jedes dieser Proteine \u200b\u200bzum richtigen Zeitpunkt. Während der Zellteilung, begleitet von der Kernteilung, werden alle diese Informationen vollständig an jeden der neu gebildeten Kerne, an jede neue Zelle übertragen. Dies ist möglich, weil sich vor der Teilung das gesamte Genmaterial der Chromosomen selbst verdoppelt, zwei identische Kopien gebildet werden und eine davon in jedem der neuen Kerne landet. Der Kern jeder Zelle enthält einen vollständigen Satz von Genen, die für einen bestimmten Organismus charakteristisch sind. Es funktionieren jedoch nicht alle Gene während des Lebens verschiedener spezialisierter Zellen. In einigen Zellen funktioniert ein Teil der Gene, in anderen - ein anderer, im dritten - ein dritter. Deshalb unterscheiden sich Zellen verschiedener Gewebe eines Organismus voneinander. Ein erheblicher Teil der Gene befindet sich in einem passiven Zustand, in dem von der Geburt bis zum Tod einer Zelle oder eines Organismus nur Informationen gespeichert werden. Darüber hinaus funktionieren Gene in einer bestimmten Zelle nicht gleichzeitig: Einige sind in einem bestimmten Lebensabschnitt der Zelle in einem bestimmten Stadium ihrer Entwicklung aktiv. andere - auf der anderen Seite usw.
Warum einige Gene inaktiv bleiben, wie andere ein- und ausgeschaltet werden, ist ein sehr komplexes und wichtiges Problem, das derzeit intensiv untersucht wird. Proteine-Histone scheinen eine wichtige Rolle beim Blockieren und Entsperren von Genen zu spielen. Sie sind Teil von Chromosomen und stehen in Verbindung mit DNA. Möglicherweise tritt die Aktivierung, "Freisetzung" eines Gens auf, wenn ein Histonmolekül von dem entsprechenden DNA-Abschnitt abgelöst wird, wodurch seine Ketten freigelegt werden und sie sich entwirren und chemisch funktionieren können. Die Anlagerung von Histon führt zur Blockade des Gens. Es ist jedoch nicht klar genug, wie die Anheftung und Ablösung von Histonen gesteuert wird.
Das Genmaterial befindet sich wie gesagt in den Chromosomen. Daher ist die Arbeit des Kerns, erbliche Informationen zu speichern, zu verdoppeln und von Zelle zu Zelle zu übertragen, die Synthese verschiedener Proteine \u200b\u200bwährend des Lebens einer Zelle zu organisieren, in erster Linie die Arbeit von Chromosomen. Obwohl sie während des gesamten Lebens der Zelle existieren, sind sie im Zellkern nur während der Zellteilung in Form klarer Strukturen unterscheidbar. Zu diesem Zeitpunkt kann die Kondensation des Materials der Chromosomen und des Chromosoms durch Mikroskopieverfahren als einzelne, klar definierte Formationen identifiziert werden.
Chromosomen haben unterschiedliche Formen. Dies sind entweder gerade oder gebogene Stöcke oder verschiedene Haken, ovale Körper, Kugeln (Abb. 36). Sie variieren stark in der Größe. Jede Pflanzenzelle einer bestimmten Art enthält in ihrem Kern den gleichen Satz (oder die gleichen Sätze) von Chromosomen aus einer genau definierten Anzahl verschiedener, aber genau definierter Chromosomen. In allen höheren und einigen niedrigeren Pflanzen wechseln sich zwei Generationen während ihres Lebenszyklus ab: mit Zellen, die einen einzelnen (haploiden, n) Chromosomensatz in ihren Kernen enthalten, und mit Zellen, deren Kerne einen doppelten (diploiden, 2 n) Chromosomensatz aufweisen. Ein haploider Satz besteht aus n verschiedenen Chromosomen, eines von jedem Typ. Der Zahlenwert ist für alle haploiden Zellen aller Pflanzen einer bestimmten Art streng konstant. Für Walderdbeeren ist diese Zahl 7, für Erbsen - ebenfalls 7, aber für Bohnen - 11, für einen Apfelbaum - 17, für eine irdene Birne - 51, für Zuckerrohr - 60. Ein diploides Set besteht aus zwei zusammengeklappten haploiden. Es enthält zwei Chromosomen jedes Typs. Wenn eine Erbse 7 verschiedene Chromosomen in einem haploiden Satz hat, dann hat ein diploider Satz 7 verschiedene Chromosomenpaare, und zwei Chromosomen in jedem Paar sind gleich. Es ist leicht zu berechnen, dass für die aufgeführten Pflanzen der diploide Satz (2n) für Erdbeeren 14, für Erbsen - 14, für Bohnen - 22, für einen Apfelbaum - 34, für eine irdene Birne - 102 und für Zuckerrohr - 120 Chromosomen beträgt.
Im Lebenszyklus von Moosen und Farnen wechseln sich Organismen mit diploiden und haploiden Zellen ab. In Farnen führen diese Organismen ein unabhängiges, isoliertes Leben. In Moosen lebt eine diploide Pflanze auf einer haploiden Pflanze. In einem diploiden Organismus bilden sich haploide Zellen - Sporen. Jeder von ihnen, der keimt, ergibt einen haploiden Organismus, dh einen Organismus, der aus haploiden Zellen aufgebaut ist. Darin bilden sich haploide Geschlechtszellen - Gameten.
Im Moment der Befruchtung verschmelzen die männlichen und weiblichen Gameten und bilden eine Zelle - eine Zygote mit einem Kern - das Ergebnis der Fusion der Kerne beider Gameten. Von jedem der Gameten erhält der Kern der Zygote einen haploiden Satz von Chromosomen, und als Ergebnis hat er einen doppelten diploiden Satz von ihnen. Ein Organismus entwickelt sich aus einer Zygote, von der jede Zelle einen diploiden Chromosomensatz aufweist.

In einem diploiden Satz sind zwei Chromosomen jedes Paares in Form und innerer Struktur identisch und enthalten Gene, die das Auftreten homogener Merkmale steuern (Abb. 36). Sie werden homologe Chromosomen genannt. Einer von ihnen stammt aus dem haploiden Satz des väterlichen Gameten, der andere aus dem mütterlichen. Daher trägt einer von ihnen in zweihäusigen Organismen Gene, die die Entwicklung der Merkmale unter seiner Gerichtsbarkeit nach dem väterlichen Typ bestimmen, der zweite nach dem mütterlichen Typ.
Homologe Chromosomen eines anderen Paares bestimmen auf die gleiche Weise die Entwicklung einer anderen Reihe von Zeichen, des dritten Paares, der dritten Reihe usw. Obwohl die väterlichen und mütterlichen Organismen derselben Art angehören, sind ihre erblichen Eigenschaften nicht identisch - sie weisen auch individuelle Unterschiede auf. Daher sind einige Gene eines homologen Chromosoms nicht mit den entsprechenden Genen des zweiten identisch.
Das haploide Set, das Teil des Diploids ist und vom väterlichen Gameten stammt, trägt die väterliche Vererbung mit seinen individuellen Merkmalen, und das haploide Set vom mütterlichen Gameten trägt das mütterliche Erbe. Das komplexe Zusammenspiel homogener, aber nicht immer identischer Gene zweier haploider Gruppen, die zusammen ein Diploid bilden, bestimmt, welche Merkmale bei den diploiden Nachkommen auftreten, die im Wesentlichen eine Mischung aus Vater und Mutter sind.
Bei Gymnospermen und Angiospermen tritt der Generationswechsel im Prinzip wie bei Moosen und Farnen auf, aber ihre haploide Phase ist stark reduziert und wird oft nur von einer Gruppe von Zellen dargestellt. Sie lebt nicht unabhängig, sondern im Körper einer haploiden Pflanze. Bei Angiospermen ist die weibliche haploide Generation im Embryosack in der Eizelle eingeschlossen, und die männliche Generation befindet sich im Pollenkorn.
Noch vor der Zellteilung bindet sich jedes DNA-Molekül in jedem Chromosom eine Kopie von sich selbst - ein zweites ähnliches Molekül. Infolgedessen verdoppelt sich das gesamte Erbmaterial der Zelle, und jedes Chromosom besteht nun aus zwei äquivalenten Teilen - Chromatiden. Ferner besteht die "Aufgabe" der Zelle darin, jedes Chromosom in Chromatiden zu unterteilen und diese streng gleichmäßig auf die zukünftigen Tochterzellen zu verteilen: Ein Chromatid von jedem Chromosom muss an jede von ihnen gesendet werden. Dies geschieht wie folgt. Unmittelbar vor der Zellteilung werden die Chromosomen stark kondensiert und kontrahiert. Dann befinden sie sich in derselben Ebene - entlang des Äquators des Kerns, wobei jeweils ein Chromatid einem Pol der Zelle zugewandt ist, dem anderen - gegenüber. Die Kernhülle verschwindet und der Nucleolus löst sich ebenfalls auf. Zwischen den Polen der Zelle erscheinen Filamente, die zusammen eine Spindelform bilden. Die Filamente werden von miteinander verbundenen Mikrotubuli gesammelt. Die Spindel besteht aus zwei Arten von Gewinden. Einige sind kontinuierliche Zellen, die von einem Pol zum anderen gehen. Andere ziehen, von denen jeder einen Pol mit einer der Chromatiden verbindet. Im Chromosom befindet sich eine Stelle - das Kinetochor, an dem die Zugfäden befestigt sind - eine von einem Pol, die andere vom anderen. Ferner divergieren die Chromatiden. Chromatiden, die zuvor jedes Chromosom bildeten, sind voneinander getrennt und divergieren, durch Fäden hochgezogen, zu entgegengesetzten Polen der Zelle. Infolgedessen wird an jedem Pol ein vollständiger Satz von Chromatiden gesammelt, die nun Chromosomen sind. Danach zerfallen die Fäden der Spindel, um jeden Chromosomensatz bildet sich eine Kernhülle, die Chromosomen quellen stark auf (despiralisieren). In jedem Kern erscheint ein Nukleolus. Der Kern erhält die Strukturcharakteristik der Kerne nicht teilender Zellen. In der Mittelebene der Zelle bildet sich ein Septum, das die Zelle in zwei Tochterzellen teilt.
Zahl: 38. Schema des Verhaltens der Chromosomen einer "bedingten" Zelle während der Meiose. A - zeigt einen diploiden Zellkern, der 3 Chromosomenpaare enthält (1 und 1 "- das 1. Paar homologer Chromosomen, 2 und 2" - das 2. Paar, 3 und 3 "- das 3. Paar). 2n \u003d 6,3 Chromosomen, die vom mütterlichen Organismus stammen, sind leicht (Zahlen ohne Schlaganfall), ihre gepaarten homologen Chromosomen, die vom väterlichen Organismus stammen, sind geschwärzt (Zahlen mit einem Schlaganfall). Homologe Chromosomen sind mit einer gemeinsamen Nummer gekennzeichnet. Jedes Chromosom besteht aus zwei Chromatiden. B - Fusion homologer Chromosomen In diesem Stadium tauschen homologe Chromosomen getrennte Abschnitte miteinander aus. Infolgedessen kommt es zu einer gewissen Umverteilung des mütterlichen und väterlichen Erbmaterials zwischen den Chromosomen (Überkreuzung). Es bilden sich C-Spindelfilamente, die sich an Chromosomen anlagern, die Kernmembran verschwindet. Zellen, an den Polen befindet sich ein homologes Chromosom von jedem Paar, die Gesamtzahl der Chromosomen an jedem Pol ist halb so hoch wie im ursprünglichen Kern A. D - Bild es gibt zwei Kerne mit jeweils einem haploiden Chromosomensatz; einer von ihnen bekam mehr Chromosomen von Großmutter (leicht, mütterlich in Bezug auf den Kern A) und weniger Großväter (geschwärzt, väterlich in Bezug auf den Kern A), im anderen - im Gegenteil; somit sind die neuen Kerne in der Zusammensetzung ihrer erblichen Substanz nicht vollständig identisch; Ihr Unterschied ist auch auf die Überkreuzung zurückzuführen, die bei Chromosomen im Stadium B auftritt. E - G - mitotische Teilung jedes der haploiden Kerne D: Längsspaltung jedes Chromosoms, Divergenz der Chromosomen zu den Polen, Bildung von zwei haploiden Kernen von jedem Kern D. Als Ergebnis erschienen 4 Zellen mit haploiden Kernen anstelle einer Zelle mit einem diploiden Chromosomensatz
Organoide werden nicht streng gleichmäßig zwischen den Tochterzellen verteilt, aber dann werden ihre Bestandteile in den Zellen synthetisiert, neue Kopien der Organellen jeder Spezies werden zusammengesetzt und ihre Anzahl in jeder Zelle wird wieder normalisiert. Die Zellen selbst wachsen. In Chromosomen wird das Erbmaterial dupliziert, wonach die Chromosomen wie vor der Zellteilung aus zwei Chromatiden bestehen. Die Zelle ist bereit für eine neue Teilung.
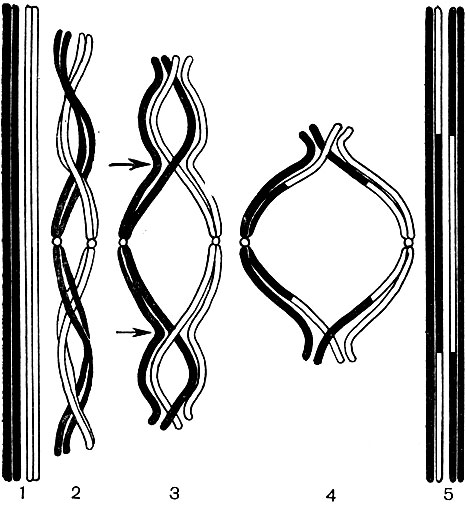
Zahl: 38A. Crossover-Diagramm während der Meiose im Stadium B Abb. 38. 1 - zwei homologe Chromosomen, von denen jedes aus zwei Schwesterchromatiden besteht; 2 - homologe Chromosomen sind miteinander verflochten; 3 - jedes Chromosom wird in seine Chromatidenbestandteile aufgeteilt; Chromosomen beginnen sich gegenseitig abzustoßen und divergieren, aber an einigen Stellen (in der Abbildung - bei vier) bleiben sie verbunden; Schwesterchromatiden werden in der zentralen Schleife verdoppelt und Nicht-Schwesterchromatiden in beiden extremen Schleifen; Punkte, an denen ein Überkreuzen möglich ist, sind durch Pfeile dargestellt; 4 - Chromatiden haben Abschnitte ausgetauscht, und Chromosomen, die eine Art gegenseitige Hybridisierung durchlaufen haben, divergieren; 5 - zwei divergierende, aber bereits "hybride" (im Gegensatz zu den auf 1 gezeigten) Chromosomen
Wenn die diploide Erzeugung von Zellen durch die haploide ersetzt wird, tritt die sogenannte Reduktionskernteilung - Meiose - auf. Während der Meiose (Abb. 38) nähern sich die homologen Chromosomen jedes Paares aneinander, schließen sich entlang ihrer Länge eng aneinander und verdrehen sich. Zwischen den zusammenhängenden homologen Chromosomen werden getrennte Schnitte ausgetauscht. Infolgedessen werden einige der Gene der väterlichen Chromosomen auf die Zusammensetzung der mütterlichen Chromosomen übertragen, und die entsprechenden Gene der mütterlichen Chromosomen nehmen die freien Stellen in den väterlichen Chromosomen ein (Überkreuzungsphänomen - Fig. 38A). Aussehen Diese und andere Chromosomen ändern sich dadurch nicht, aber ihre qualitative Zusammensetzung wird unterschiedlich. Das väterliche und mütterliche Erbe wird neu verteilt und gemischt. Ferner lösen sich die Kernhülle und der Nucleolus auf, es wird ein Spindelapparat gebildet, genau wie während der Mitose. Homologe Chromosomen werden getrennt und divergieren mit Hilfe von Spindelfäden zu den Polen der Zelle. Ein Pol hat einen haploiden Chromosomensatz (ein homologes Chromosom von jedem Paar), der andere hat einen zweiten haploiden Satz.
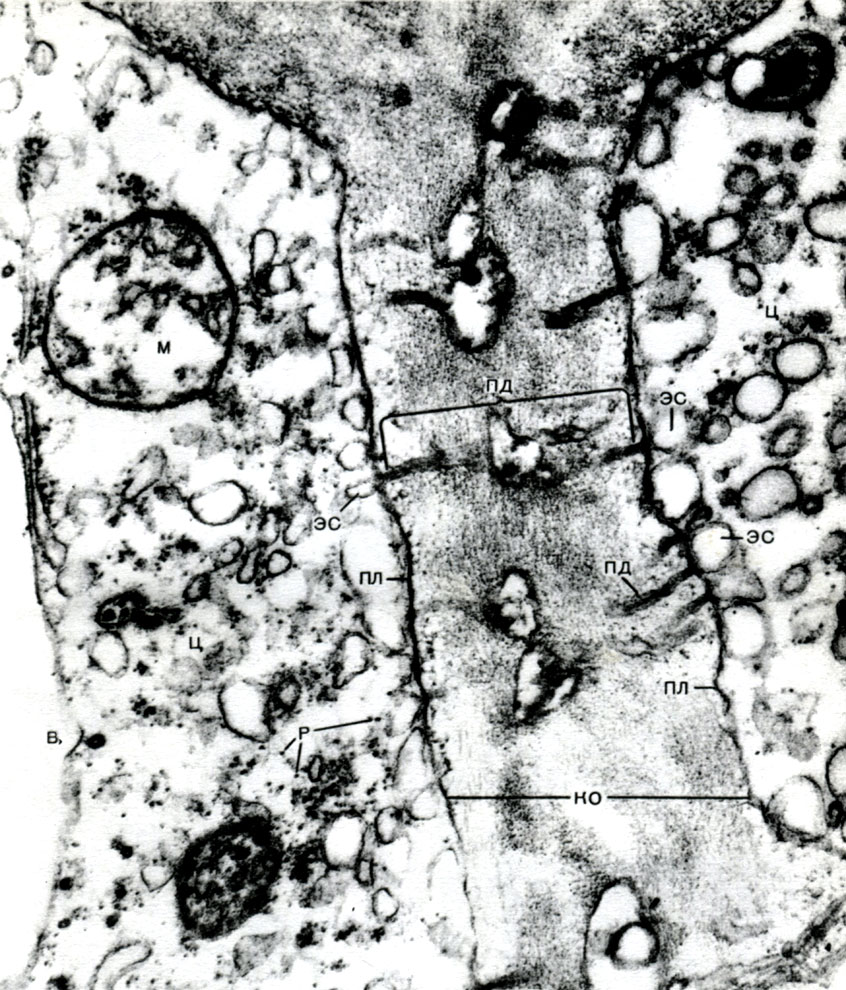
Nach der Meiose teilen sich die gebildeten haploiden Kerne durch Mitose. In diesem Fall wird jedes Chromosom des haploiden Satzes in zwei Chromatiden aufgeteilt, sie divergieren und es bilden sich haploide Tochterzellen.
Meiose unterscheidet sich von Mitose in zwei grundlegenden Punkten. Erstens klumpen während der Meiose jedes Paar homologer Chromosomen zusammen und ein Teil des Erbmaterials wird zwischen ihnen ausgetauscht, was während der Mitose nicht der Fall ist. Zweitens divergieren während der Meiose homologe Chromosomen zu den Polen der Zelle, eines von jedem Paar, und während der Mitose wird jedes Chromosom in Chromatiden aufgeteilt, und Chromatiden divergieren zu den Polen, eines von jedem Chromosom. Infolge der Meiose werden haploide Zellen aus einer diploiden Zelle gebildet.
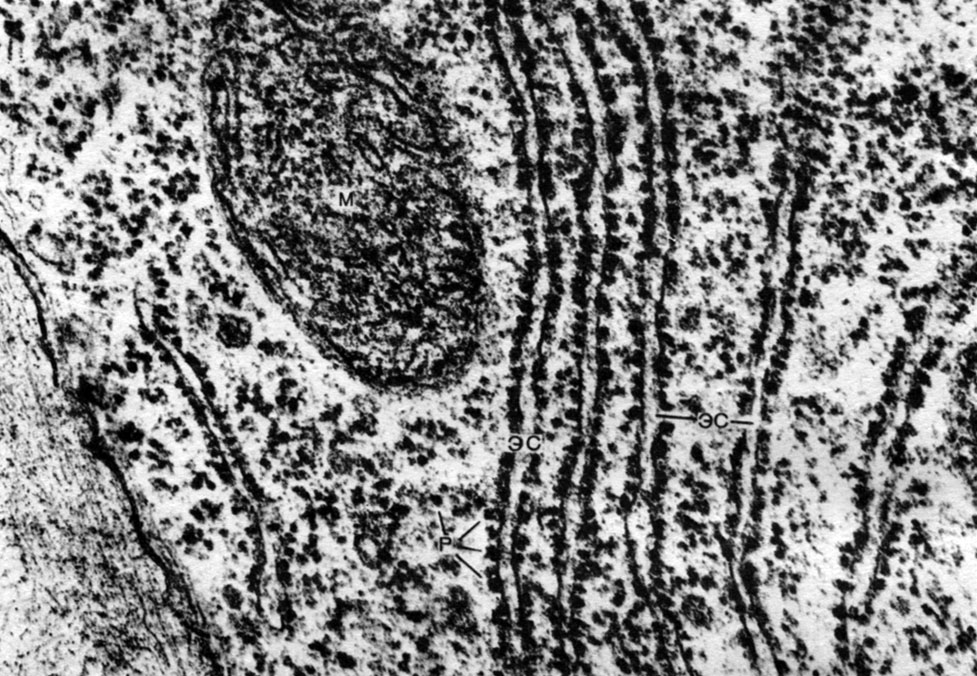
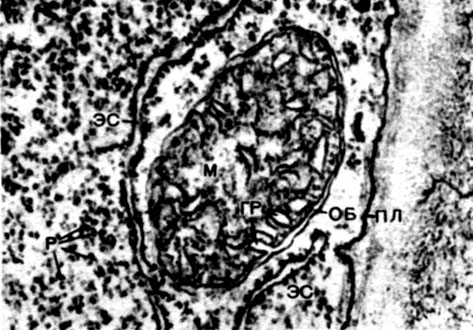
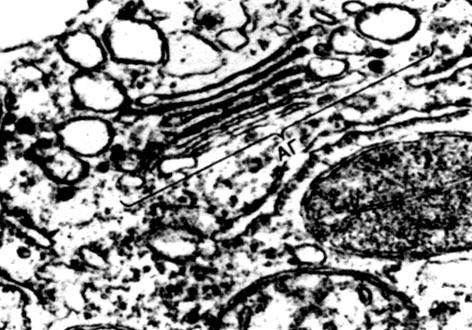
Tabelle 10. Pflanzenzellstruktur. Golgi-Apparat im Zytoplasma des sich entwickelnden Wurzelhaars eines Rettichs (Raphanus sativus). Elektronenmikroskopische Aufnahme (Vergrößerung x 52000) von M. F. Danilova: ag - Golgi-Apparat, Abschnitte von flachen Säcken und Blasen sind sichtbar
In den Zellen einiger Gewebe tritt während ihrer Entwicklung eine unvollständige Mitose auf: Das Chromosomenmaterial in den Kernen verdoppelt sich, die Chromosomen teilen sich in zwei Hälften, aber anstatt zwei Kerne zu bilden, verbleiben sie im ursprünglichen Kern. Von diesem Moment an enthält es keinen diploiden, sondern einen tetraploiden (vierfachen) Chromosomensatz. Der Prozess, der zu dieser Verdoppelung der Chromosomen innerhalb eines Kerns führt, wird als Endomitose - interne Mitose bezeichnet. Wenn es einem Kern zweimal passiert, wird es zu einem oktoploiden (achtfachen) Satz usw. Zellen, deren Kerne mehr als zwei Chromosomensätze tragen, werden als polyploide, dh multiploide bezeichnet. Zellpolyploidie erhöht in einigen Fällen ihre Lebensfähigkeit, da jedes Gen von mehreren anderen Genen desselben dupliziert wird. Single-Quality-Gene wirken im Einklang, und eine Schädigung eines dieser Gene führt nicht zum Verlust des von ihm bestimmten Merkmals, da es durch die Arbeit der verbleibenden homogenen Gene kompensiert wird. In vielen Fällen sind polyploide Zellen größer und inhaltsreicher als diploide Zellen. Es wurden Sorten polyploider Kulturpflanzen entwickelt, die die wirtschaftlichen Qualitäten erhöht haben.
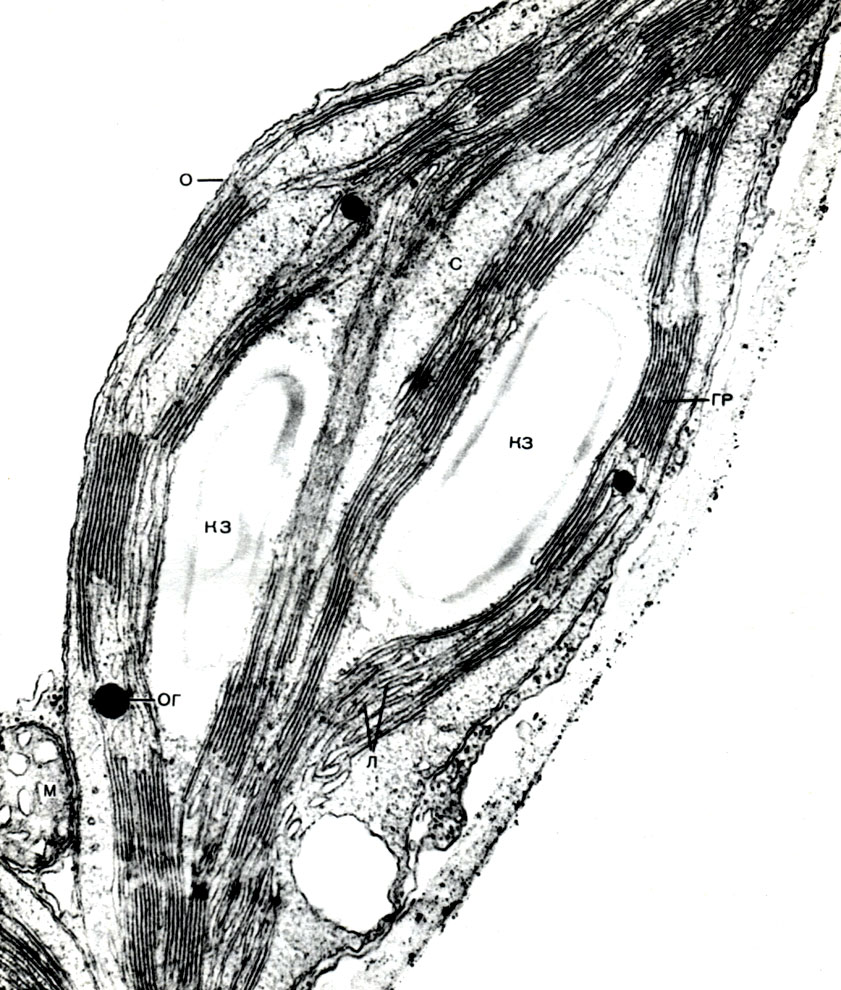
Die Wissenschaft, die die Zelle untersucht, heißt Zytologie. Die Zelle ist die Grundlage der Struktur aller Lebewesen und aller Lebensprozesse; Daher können die meisten der wichtigsten allgemeinen biologischen Probleme unter Einbeziehung der Zytologie und der Methoden ihrer Forschung gelöst werden.
Mit Hilfe der Zytologie werden viele praktisch wichtige Probleme gelöst: der Kampf gegen Pflanzenkrankheiten (Pilze, Bakterien), die Züchtung neuer Sorten von Kulturpflanzen, die Überwindung der Sterilität (Sterilität) von Hybridsorten.
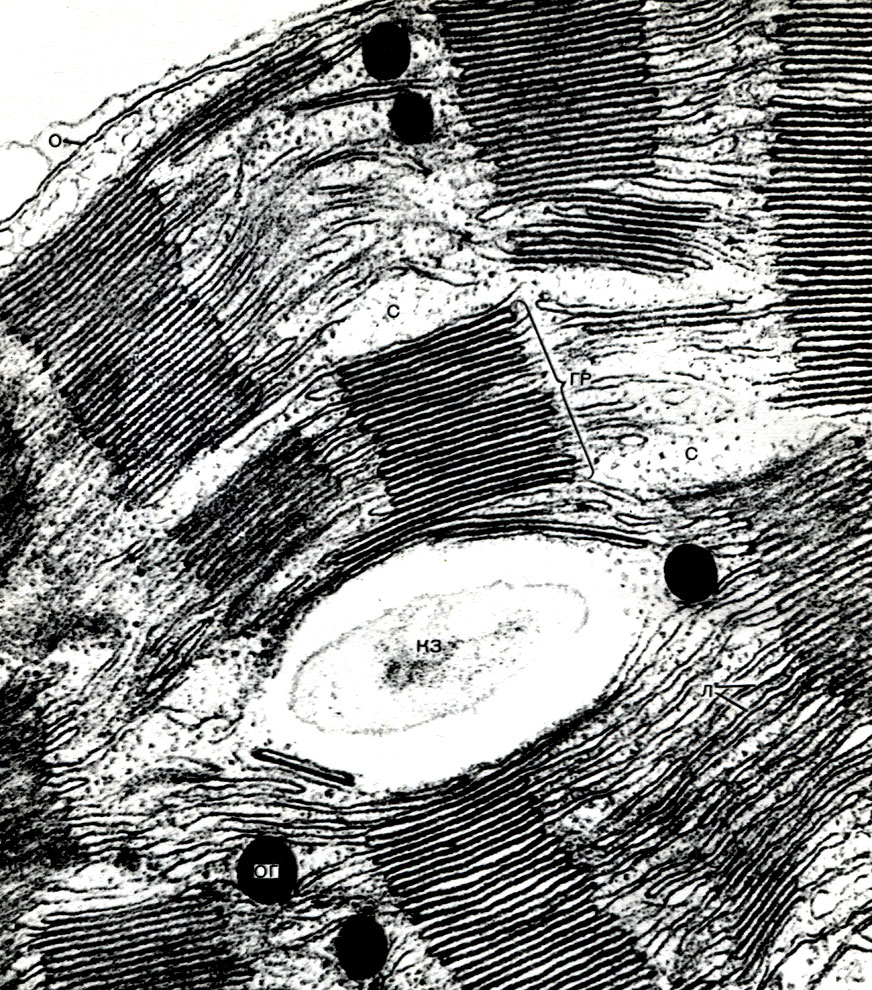
Die moderne Zytologie untersucht diese Probleme mit einer Vielzahl von Methoden - mikroskopisch, biochemisch, biophysikalisch, genetisch - und in enger Wechselwirkung mit anderen verwandten biologischen Wissenschaften.