VIRUSURI, cei mai mici agenți patogeni ai bolilor infecțioase. Tradus din virusul latin înseamnă „otravă, principiu otrăvitor”. Până la sfârșitul secolului al XIX-lea. termenul „virus” a fost folosit în medicină pentru a se referi la orice agent infecțios care cauzează boli. Acest cuvânt și-a dobândit semnificația modernă după 1892, când botanistul rus DI Ivanovsky a stabilit „filtrabilitatea” agentului cauzator al bolii mozaicului tutunului (mozaicul tutunului). El a arătat că seva celulară de la plantele infectate cu această boală, a trecut prin filtre speciale care prind bacteriile, păstrează capacitatea de a provoca aceeași boală la plantele sănătoase. Cinci ani mai târziu, un alt agent de filtrare, agentul cauzal al febrei aftoase la bovine, a fost descoperit de bacteriologul german F. Löffler. În 1898, botanistul olandez M. Beijerink a repetat aceste experimente într-o versiune extinsă și a confirmat concluziile lui Ivanovsky. El a numit „principiul otrăvitor filtrabil” care cauzează mozaicul tutunului „virus filtrabil”. Acest termen a fost folosit de mulți ani și a fost redus treptat la un singur cuvânt - „virus”.
În 1901, chirurgul militar american W. Reed și colegii săi au constatat că agentul cauzal al febrei galbene este, de asemenea, un virus filtrabil. Febra galbenă a fost prima boală umană identificată ca virală, dar a durat încă 26 de ani pentru ca originea sa virală să fie definitiv dovedită.
Se acceptă în general că virusurile au apărut ca urmare a izolării (autonomizării) elementelor genetice individuale ale celulei, care, în plus, au primit capacitatea de a fi transmise de la organism la organism. Într-o celulă normală, mai multe tipuri de structuri genetice se mișcă, de exemplu, matricial sau informațional, ARN (ARNm), transpozoni, introni și plasmide. Astfel de elemente mobile ar fi putut fi predecesorii sau progenitorii virușilor.
Virusurile sunt organisme vii? În 1935, biochimistul american W. Stanley a izolat virusul mozaicului tutunului sub formă cristalină, demonstrând astfel natura sa moleculară. Rezultatele obținute au stârnit dezbateri aprinse despre natura virușilor: sunt organisme vii sau doar molecule activate? Într-adevăr, în interiorul celulei infectate, virușii se manifestă ca componente integrale ale sistemelor vii mai complexe, dar în afara celulei sunt nucleoproteine \u200b\u200binerte din punct de vedere metabolic. Virușii conțin informații genetice, dar nu o pot implementa independent fără a avea propriul mecanism de sinteză a proteinelor. Când trăsăturile structurii și reproducerii virușilor au fost clarificate, întrebarea dacă acestea sunt în viață și-a pierdut treptat semnificația.
STRUCTURA VIRUSELOR
Complet în structură și infecțios, adică capabil să provoace infecție, o particulă virală în afara celulei se numește virion. Nucleul („nucleul”) virionului conține o moleculă și, uneori, două sau mai multe molecule de acid nucleic. Învelișul proteic care acoperă acidul nucleic virionic și îl protejează de influențele nocive ale mediului se numește capsidă. Acidul nucleic virionic este materialul genetic al virusului (genomul acestuia) și este reprezentat de acidul dezoxiribonucleic (ADN) sau acidul ribonucleic (ARN), dar niciodată de acești doi compuși simultan. (Chlamydia, rickettsia și toate celelalte microorganisme „cu adevărat vii” conțin atât ADN, cât și ARN.) Acizii nucleici ai celor mai mici viruși conțin trei sau patru gene, în timp ce cei mai mari viruși au până la o sută de gene.
Unii viruși, pe lângă capsidă, au și un înveliș exterior format din proteine \u200b\u200bși lipide. Se formează din membranele celulei infectate care conțin proteine \u200b\u200bvirale încorporate. Termenii „virioni goi” și „virioni neacoperiți” sunt folosiți sinonim. Capsidele celor mai mici și mai simplu aranjați viruși pot consta doar din unul sau mai multe tipuri de molecule proteice. Mai multe molecule din aceleași sau diferite proteine \u200b\u200bsunt combinate în subunități numite capsomere. Capsomerii, la rândul lor, formează structuri geometrice regulate ale capsidei virale. La diferiți viruși, forma capsidei este o trăsătură caracteristică (trăsătură) a virionului.
Virionii cu un tip de spirală de simetrie, precum virusul mozaicului tutunului, au forma unui cilindru alungit; în interiorul învelișului proteic, care constă din subunități separate - capsomere, există o spirală spirală a acidului nucleic (ARN). Virionii cu un tip de simetrie icosaedrică (din grecescul eikosi - douăzeci, hedra - suprafață), ca un poliovirus, au o formă sferică, sau mai bine zis, cu mai multe fațete; capsidele lor sunt construite din 20 de fațete (suprafețe) triunghiulare regulate și arată ca o cupolă geodezică.
Bacteriofagii individuali (virusurile bacteriilor; fagii) au un tip mixt de simetrie. La așa-numitul. Dintre fagii „cu coadă”, capul arată ca o capsidă sferică; un proces tubular lung - „coada” se îndepărtează de el.
Există viruși cu o structură și mai complexă. Virionii variolei (viruși variolici) nu au o capsidă obișnuită, tipică: structurile tubulare și membranare sunt situate între miez și învelișul exterior.
Informațiile genetice codificate într-o singură genă pot fi văzute în general ca instrucțiuni pentru producerea unei proteine \u200b\u200bspecifice într-o celulă. O astfel de instrucțiune este percepută de celulă numai dacă este trimisă sub formă de ARNm. Prin urmare, celulele în care materialul genetic este reprezentat de ADN trebuie să „rescrie” (transcrie) aceste informații într-o copie complementară a proteinelor virusului mARN (vezi și ACIDI NUCLEICI). Virusurile care conțin ADN în modul de replicare diferă de virusurile care conțin ARN.
ADN-ul există de obicei sub formă de structuri dublu catenare: două lanțuri polinucleotidice sunt legate de hidrogen și răsucite în așa fel încât se formează o helică dublă. În schimb, ARN-ul există de obicei sub formă de structuri monocatenare. Cu toate acestea, genomul virușilor individuali este ADN monocatenar sau ARN bicatenar. Catenele (lanțurile) de acid nucleic viral, duble sau simple, pot fi liniare sau închise într-un inel.
Prima etapă a replicării virale este asociată cu penetrarea acidului nucleic viral în celula gazdă. Acest proces poate fi facilitat de enzime speciale care alcătuiesc capsida sau învelișul exterior al virionului, iar învelișul rămâne în afara celulei sau virionul îl pierde imediat după pătrunderea în celulă. Virusul găsește o celulă potrivită pentru reproducerea sa prin contactarea părților individuale ale capsidei (sau învelișului exterior) cu receptori specifici de pe suprafața celulei într-o manieră de „blocare a cheii”. Dacă sunt absenți receptori specifici („de recunoaștere”) de pe suprafața celulei, atunci celula nu este sensibilă la infecția virală: virusul nu pătrunde în ea.
Pentru a-și realiza informațiile genetice, ADN-ul viral care a pătruns în celulă este transcris de enzime speciale în ARNm. ARNm rezultat se mută în „fabricile” celulare de sinteză a proteinelor - ribozomi, unde înlocuiește „mesajele” celulare cu propriile „instrucțiuni” și este tradus (citit), în urma cărora sunt sintetizate proteinele virale. ADN-ul viral în sine este în mod repetat duplicat (duplicat) cu participarea unui alt set de enzime, atât virale, cât și aparținând celulei.
Proteina sintetizată, care este utilizată pentru construirea capsidei, și ADN-ul viral înmulțit în multe exemplare se combină și formează noi virioni „fiice”. Descendenții virali formați părăsesc celula utilizată și infectează altele noi: ciclul de reproducere a virusului se repetă. Unii viruși, în timpul înmuguririi de pe suprafața celulei, captează o parte a membranei celulare, în care proteinele virale au fost inserate „în avans” și astfel dobândesc o membrană. În ceea ce privește celula gazdă, în cele din urmă se dovedește a fi deteriorată sau chiar complet distrusă.
În unele virusuri ADN, ciclul de reproducere în sine în celulă nu este asociat cu replicarea imediată a ADN-ului viral; în schimb, ADN-ul viral este inserat (integrat) în ADN-ul celulei gazdă. În acest stadiu, virusul ca o singură formațiune structurală dispare: genomul său devine parte a aparatului genetic al celulei și chiar se reproduce ca parte a ADN-ului celular în timpul diviziunii celulare. Cu toate acestea, ulterior, uneori după mulți ani, virusul poate apărea din nou - se declanșează mecanismul de sinteză a proteinelor virale, care, combinându-se cu ADN-ul viral, formează noi virioni.
În unele virusuri ARN, genomul (ARN) poate acționa direct ca ARNm. Cu toate acestea, această caracteristică este caracteristică numai pentru virușii cu o catenă de ARN "+" (adică, cu ARN având o polaritate pozitivă). La virușii cu un fir de ARN "-", acesta din urmă trebuie mai întâi să fie "rescris" într-un fir "+"; abia după aceea începe sinteza proteinelor virale și virusul se reproduce.
Așa-numitele retrovirusuri conțin ARN ca genom și au un mod neobișnuit de transcriere a materialului genetic: în loc să transcrie ADN în ARN, așa cum se întâmplă într-o celulă și este tipic pentru virusurile care conțin ADN, ARN-ul lor este transcris în ADN. ADN-ul dublu catenar al virusului este apoi încorporat în ADN-ul cromozomial al celulei. Pe matricea unui astfel de ADN viral, se sintetizează un nou ARN viral care, la fel ca altele, determină sinteza proteinelor virale. A se vedea și RETROVIRUSE.
Dacă virușii sunt într-adevăr elemente genetice mobile care au primit „autonomie” (independență) față de aparatul genetic al gazdelor lor (diferite tipuri de celule), atunci diferite grupuri de viruși (cu genom, structură și replicare diferite) ar fi trebuit să apară independent unul de celălalt. Prin urmare, este imposibil să se construiască pentru toți virușii un singur pedigree care să le lege pe baza relațiilor evolutive. Principiile clasificării „naturale” utilizate în taxonomia animalelor nu sunt potrivite pentru viruși.
Cu toate acestea, un sistem de clasificare a virusului este necesar în practică, iar încercările de creare a acestuia au fost făcute în mod repetat. Cea mai productivă abordare s-a dovedit a fi bazată pe caracteristicile structurale și funcționale ale virușilor: pentru a distinge diferitele grupuri de viruși unul de celălalt, aceștia descriu tipul acidului lor nucleic (ADN sau ARN, fiecare dintre aceștia putând fi monocatenar sau dublu catenar), dimensiunea acestuia (numărul de nucleotide din lanțul acidului nucleic). acizi), numărul moleculelor de acid nucleic dintr-un virion, geometria virionului și caracteristicile structurale ale capsidei și ale învelișului exterior al virionului, tipul gazdei (plante, bacterii, insecte, mamifere etc.), caracteristicile patologiei cauzate de viruși (simptome și natura bolii), proprietățile antigenice ale proteinelor virale și caracteristicile reacției sistemului imunitar al organismului la introducerea virusului.
Un grup de agenți patogeni microscopici numiți viroizi (adică particule asemănătoare virusului) nu se încadrează în sistemul de clasificare a virusurilor. Viroizii provoacă multe boli comune ale plantelor. Aceștia sunt cei mai mici agenți infecțioși, lipsiți chiar și de cea mai simplă înveliș proteic (găsit în toate virusurile); constau doar din ARN monocatenar închis într-un inel.
BOLI VIRALE
Pentru mulți viruși, cum ar fi rujeola, herpesul și parțial gripa, oamenii sunt principalul rezervor natural. Transmiterea acestor viruși are loc prin picături aeriene sau prin contact.
Distribuirea unora boli virale, ca și alte infecții, este plin de surprize. De exemplu, în grupuri de oameni care trăiesc în condiții insalubre, aproape toți copiii din vârstă fragedă poartă poliomielită, de obicei ușoară, și dobândesc imunitate. Dacă condițiile de viață din aceste grupuri se îmbunătățesc, copiii vârsta mai mică poliomielita nu este de obicei bolnavă, dar boala poate apărea la o vârstă mai înaintată, iar apoi este adesea severă.
Mulți viruși nu pot persista în natură mult timp la o densitate redusă de dispersie a speciei gazdă. Lipsa populațiilor de vânători primitivi și colecționari de plante a creat condiții nefavorabile pentru existența unor viruși; prin urmare, este foarte probabil ca unele virusuri umane să apară mai târziu, odată cu apariția așezărilor urbane și rurale. Se presupune că virusul rujeolei a existat inițial în rândul câinilor (ca agent cauzal al febrei), iar variola la om ar fi putut apărea ca urmare a evoluției variolei la vaci sau șoareci. Cele mai recente exemple de evoluție virală includ sindromul imunodeficienței dobândite (SIDA). Există dovezi ale asemănărilor genetice între virusurile imunodeficienței umane și maimuțele verzi africane.
Infecțiile „noi” sunt de obicei severe, adesea fatale, dar pe parcursul evoluției agentului patogen pot deveni mai ușoare. Un bun exemplu este istoria virusului mixomatozei. În 1950, acest virus, endemic în America de Sud și destul de inofensiv pentru iepurii locali, a fost introdus în Australia împreună cu rasele europene ale acestor animale. Boala la iepurii australieni, care nu întâlnise anterior acest virus, a fost fatală în 99,5% din cazuri. Câțiva ani mai târziu, mortalitatea din această boală a scăzut semnificativ, în unele zone până la 50%, ceea ce se explică nu numai prin mutații „atenuante” (slăbind) în genomul viral, ci și prin rezistența genetică crescută a iepurilor la boală și, în ambele cazuri, selecția naturală efectivă a avut loc sub presiune puternică a selecției naturale.
Reproducerea virușilor în natură este susținută de diferite tipuri de organisme: bacterii, ciuperci, protozoare, plante, animale. De exemplu, insectele suferă adesea de viruși care se acumulează în celulele lor sub formă de cristale mari. Plantele sunt adesea atacate de viruși ARN mici și aranjați simplu. Acești viruși nu au nici măcar mecanisme speciale pentru a intra în celulă. Sunt transportate de insecte (care se hrănesc cu seva celulară), viermi rotunzi și prin contact, infectând planta dacă este deteriorată mecanic. Virușii bacterieni (bacteriofagi) au cel mai complex mecanism de livrare a materialului genetic către o celulă bacteriană sensibilă. În primul rând, „coada” fagului, care arată ca un tub subțire, se atașează de peretele bacteriei. Apoi, enzime speciale ale „cozii” dizolvă o secțiune a peretelui bacterian și materialul genetic al fagului (de obicei ADN) este injectat în gaura rezultată prin „coadă”, ca printr-un ac de seringă.
Peste zece grupuri majore de viruși sunt patogeni pentru oameni. Printre virușii care conțin ADN, aceasta este familia de variole (care provoacă variolă, vaccin și alte infecții cu variolă), viruși din grupul herpes (răni de herpes pe buze, varicela), adenovirusuri tractului respirator și ochi), familia papovavirusului (negii și alte creșteri ale pielii), hepadnavirusurile (virusul hepatitei B). Există mult mai mulți viruși care conțin ARN, care sunt patogeni pentru oameni. Picornavirusurile (din latinescul pico - foarte mic, ARN englez - ARN) sunt cei mai mici viruși de mamifere, asemănători unor viruși vegetali; provoacă poliomielită, hepatită A, acută răceli... Mixovirusurile și paramixovirusurile sunt cauza diferitelor forme de gripă, rujeolă și oreion (oreion). Arbovirusurile (din engleză arthropod borne - „purtate de artropode”) - cel mai mare grup de viruși (mai mult de 300) - sunt transportate de insecte și sunt agenții cauzali ai encefalitei transmise prin căpușe și japoneze, febra galbenă, meningoencefalita ecvină, febra Colorado transmise prin căpușe, encefalita scoțiană și alte boli ... Reovirusurile, care sunt agenți cauzali mai degrabă rari ai bolilor respiratorii și intestinale la om, au devenit subiectul unui interes științific special datorită faptului că materialul lor genetic este reprezentat de ARN fragmentat dublu catenar. Vezi și BOLI VENERALE; VARICELĂ; HEPATITA; FLU; FEBRA DENGE; MONONUCLEOZA INFECȚIOASĂ; POJAR; KRASNUHA; MENINGITA; PISC NATURAL; POLIO; BOLI VIRALE RESPIRATOARE; DE PORC; SINDROMUL IMUNODEFICIENȚEI OBȚINUTE (SIDA); ENCEFALITĂ.
Agenții cauzali ai unor boli, inclusiv cele foarte grave, nu se încadrează în niciuna dintre categoriile de mai sus. Pentru un grup special de lent infecții virale mai recent, de exemplu, au fost menționate boala Creutzfeldt-Jakob și Kuru, boli degenerative ale creierului, cu o perioadă foarte lungă de incubație. Cu toate acestea, sa dovedit că acestea nu sunt cauzate de viruși, ci de cei mai mici agenți infecțioși de natură proteică - prioni (a se vedea PRION).
Tratament și prevenire. Reproducerea virușilor este strâns legată de mecanismele de sinteză a proteinelor și acizilor nucleici ai celulei din organismul infectat. Prin urmare, crearea de medicamente care suprimă selectiv virusul, dar nu dăunează organismului, este o sarcină extrem de dificilă. Cu toate acestea, s-a dovedit că, în cele mai mari virusuri ale herpesului și ale variolei, ADN-ul genomic codifică un număr mare de enzime care diferă prin proprietăți de enzimele celulare similare, iar acest lucru a servit ca bază pentru dezvoltarea medicamente antivirale... Într-adevăr, au fost create mai multe medicamente al căror mecanism de acțiune se bazează pe suprimarea sintezei ADN-ului viral. Unii compuși care sunt prea toxici pentru uz general (intravenos sau oral) sunt potriviți pentru utilizare topică, cum ar fi atunci când ochii sunt deteriorați de virusul herpes.
Se știe că organismul uman produce proteine \u200b\u200bspeciale - interferoni. Acestea suprimă translația acizilor nucleici virali și astfel inhibă reproducerea virusului. Datorită ingineriei genetice, interferonii produși de bacterii au devenit disponibile și sunt testați în practica medicală (vezi GENE ENGINEERING).
Cele mai eficiente elemente de apărare naturală ale organismului includ anticorpi specifici (proteine \u200b\u200bspeciale produse de sistemul imunitar), care interacționează cu virusul corespunzător și astfel previn în mod eficient dezvoltarea bolii; cu toate acestea, ei nu pot neutraliza un virus care a intrat deja în celulă. Un exemplu este infecție cu herpes: Virusul herpes persistă în celulele ganglionilor nervoși (ganglioni) unde anticorpii nu pot ajunge la el. Din când în când, virusul este activat și provoacă recidive ale bolii.
De obicei, anticorpii specifici se formează în organism ca urmare a pătrunderii unui agent infecțios. Organismul poate fi ajutat prin creșterea artificială a producției de anticorpi, inclusiv prin construirea imunității în prealabil, prin vaccinare. În acest fel, prin vaccinarea în masă, boala variolei a fost practic eliminată în întreaga lume. A se vedea, de asemenea, VACCAREA ȘI IMUNIZAREA.
Metodele moderne de vaccinare și imunizare sunt împărțite în trei grupe principale. În primul rând, aceasta este utilizarea unei tulpini slăbite a virusului, care stimulează producerea de anticorpi în organism care sunt eficienți împotriva unei tulpini mai patogene. În al doilea rând, introducerea unui virus ucis (de exemplu, inactivat cu formalină), care induce și formarea de anticorpi. A treia opțiune este așa-numita. Imunizarea „pasivă”, adică introducerea de anticorpi „străini” gata preparați. Un animal, cum ar fi un cal, este imunizat, apoi anticorpii sunt izolați din sângele său, purificați și utilizați pentru administrare la un pacient pentru a crea imunitate imediată, dar pe termen scurt. Uneori, anticorpii sunt folosiți din sângele unei persoane care a avut boala (de exemplu, rujeolă, encefalită transmisă de căpușe).
Acumularea de viruși. Pentru a pregăti vaccinuri, este necesar să se acumuleze virusul. În acest scop, se folosesc adesea embrioni de pui în curs de dezvoltare, care sunt infectați cu acest virus. După incubarea embrionilor infectați pentru o anumită perioadă de timp, virusul acumulat în ele datorită reproducerii este colectat, purificat (prin centrifugare sau într-un alt mod) și, dacă este necesar, inactivat. Este foarte important să eliminați toate impuritățile de balast din preparatele virusului care pot provoca complicații grave în timpul vaccinării. Desigur, este la fel de important să vă asigurați că nu mai există niciun virus patogen neinactivat în preparate. În ultimii ani, diferite tipuri de culturi celulare au fost utilizate pe scară largă pentru acumularea de viruși.
METODE PENTRU STUDIAREA VIRUSELOR
Virușii bacterieni au fost primii care au devenit obiectul unor studii detaliate ca cel mai convenabil model cu o serie de avantaje față de alți viruși. Ciclul complet de replicare a fagilor, adică timpul de la infecția unei celule bacteriene până la eliberarea particulelor virale multiplicate din aceasta are loc în decurs de o oră. Alte virusuri se acumulează de obicei în câteva zile sau chiar mai mult. Cu puțin timp înainte de al doilea război mondial și la scurt timp după încheierea acestuia, au fost dezvoltate metode pentru a studia particulele virale individuale. Plăcile cu agar nutritiv, pe care s-a crescut un monostrat (strat continuu) de celule bacteriene, sunt infectate cu particule de fagi folosind diluțiile sale seriale. Reproducându-se, virusul ucide celula „protejată” și pătrunde în celulele vecine, care mor și după acumularea descendenților fagici. Zona celulelor moarte este vizibilă cu ochiul liber ca un punct luminos. Astfel de pete se numesc „colonii negative” sau plăci. Metoda dezvoltată a făcut posibilă studierea descendenților particulelor virale individuale, detectarea recombinării genetice a virușilor și determinarea structurii genetice și a metodelor de replicare a fagilor în detalii care anterior păreau incredibile.
Lucrul cu bacteriofagi a contribuit la extinderea arsenalului metodologic în studiul virusurilor animale. Înainte de aceasta, studiile asupra virusurilor vertebrate au fost efectuate în principal pe animale de laborator; astfel de experimente au fost foarte laborioase, costisitoare și nu prea informative. Ulterior, au apărut noi metode bazate pe utilizarea culturilor de țesuturi; celulele bacteriene utilizate în experimentele de fagi au fost înlocuite cu celule de vertebrate. Cu toate acestea, experimentele pe animale de laborator sunt foarte importante pentru studierea mecanismelor de dezvoltare a bolilor virale și continuă să fie efectuate în prezent.
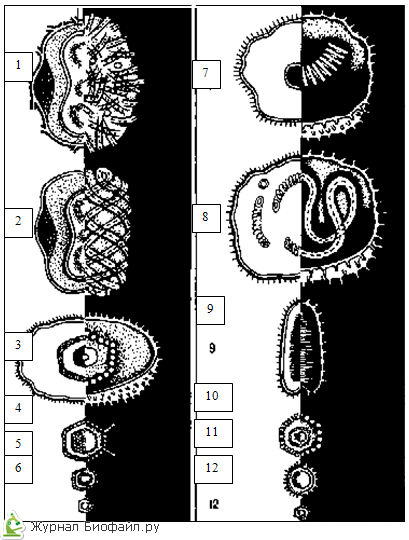
LA TIPUL ICOSAHEDRIC DE SIMETRIE, prezentat în diagrama structurii adenovirusului, capsomerilor sau subunităților proteice ale virusului, formează o teacă proteică izometrică, formată din 20 de triunghiuri regulate.
În cazul unei simetrii spirale, prezentată în diagrama structurii virusului mozaicului tutunului, capsomerilor sau subunităților virusului, formează o spirală în jurul miezului tubular gol.
Simetria COMBINATĂ sau mixtă în viruși poate fi reprezentată de diferite opțiuni. Particula de bacteriofag prezentată în diagramă are un „cap” de formă geometrică regulată și o „coadă” cu simetrie spirală.
Cristalizarea virușilor
În 1932, un tânăr biochimist american, Wendill Stanley, a fost rugat să studieze virușii. Stanley a început prin a strânge o sticlă de suc dintr-o tonă de frunze de tutun infectate cu virusul mozaicului tutunului. A început să studieze sucul folosind metodele chimice de care dispunea. El a expus diferite fracțiuni ale sucului la diferiți reactivi, sperând să obțină o proteină virală pură (Stanley era convins că un virus este o proteină). Odată, Stanley a primit o fracțiune aproape pură a unei proteine \u200b\u200bcare diferă în compoziție de proteinele celulelor vegetale. Omul de știință și-a dat seama că în fața lui se afla ceea ce se străduia atât de mult. Stanley a izolat o proteină extraordinară, a dizolvat-o în apă și a pus soluția în frigider. A doua zi dimineață, într-un balon lichid limpede erau frumoase cristale asemănătoare acelor. Stanley a luat o lingură din aceste cristale dintr-o tonă de frunze. Apoi Stanley a turnat câteva cristale, le-a dizolvat în apă, a umezit tifon cu această apă și a frecat cu ea frunzele plantelor sănătoase. Seva plantelor a suferit o gamă întreagă de influențe chimice. După o astfel de „prelucrare masivă”, cel mai probabil, virușii ar fi trebuit să moară.
Frunzele frecate sunt bolnave. Deci, proprietățile ciudate ale virusului au fost completate de o alta - capacitatea de a cristaliza.
Efectul de cristalizare a fost atât de copleșitor încât Stanley a renunțat la ideea că un virus a fost o creatură pentru o lungă perioadă de timp. Deoarece toate enzimele sunt proteine \u200b\u200bși multe enzime cresc, de asemenea, pe măsură ce organismul se dezvoltă și se pot cristaliza, Stanley a concluzionat că virusurile sunt proteine \u200b\u200bpure, mai degrabă enzime.
Oamenii de știință au devenit în curând convinși că este posibil să cristalizeze nu numai virusul mozaicului tutunului, ci și o serie de alți viruși.
Cinci ani mai târziu, biochimiștii englezi F. Bowden și N. Peary au găsit o eroare în definiția lui Stanley: 94% din conținutul virusului mozaicului tutunului era format din proteine, iar 6% era acid nucleic. Virusul nu era de fapt o proteină, ci o nucleoproteină - o combinație de proteine \u200b\u200bși acid nucleic.
De îndată ce microscopii electronici au devenit disponibili biologilor, oamenii de știință au stabilit că cristalele virale constau în câteva sute de miliarde de particule strâns presate împreună. Există atât de multe particule într-un cristal al virusului poliomielitei încât pot infecta toți locuitorii Pământului de mai multe ori. Când a fost posibil să se examineze particulele virale individuale la microscopul electronic, s-a dovedit că acestea vin în diferite forme, dar învelișul exterior al virușilor constă întotdeauna din proteine \u200b\u200bcare diferă în viruși diferiți, ceea ce le permite să fie recunoscute folosind reacții imunologice, iar conținutul interior este reprezentat de acidul nucleic, care este o unitate a eredității.
Componentele virușilorCel mai mare virus (virusul variolei) se apropie ca mărime de bacteriile mici, cel mai mic (agenți cauzali ai encefalitei, poliomielitei, febrei aftoase) - la moleculele proteice mari. Cu alte cuvinte, virușii au propriii lor giganți și pitici. (vezi Fig. 1) Pentru a măsura virușii, se folosește o convenție numită nanometru (nm). Un nm este o milionime de milimetru. Dimensiunile diferiților viruși variază de la 20 la 300 nm.
Deci, virușii sunt compuși din mai multe componente:
nucleu - material genetic (ADN sau ARN). Aparatul genetic al virusului poartă informații despre mai multe tipuri de proteine \u200b\u200bcare sunt necesare pentru formarea unui nou virus.
un strat proteic numit capsidă. Plicul este adesea construit din subunități repetate identice - capsomere. Capsomerii formează structuri cu un grad ridicat de simetrie.
Membrană lipoproteică suplimentară. Se formează din membrana plasmatică a celulei gazdă. Se găsește numai în virusuri relativ mari (gripă, herpes). Acest înveliș exterior este un fragment al membranei nucleare sau citoplasmatice a celulei gazdă din care virusul pătrunde în mediul extracelular. Uneori, în cojile exterioare ale virusurilor complexe, pe lângă proteine, carbohidrații sunt conținuți, de exemplu, în agenți cauzali ai gripei și herpesului.
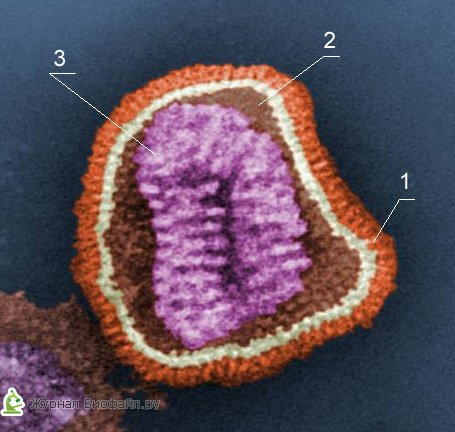
1. Coajă suplimentară
2. Capsomer (strat proteic)
3. Miezul (ADN sau ARN)
Fiecare componentă a virionilor are anumite funcții: stratul proteic îi protejează de efectele adverse, acidul nucleic este responsabil pentru proprietățile ereditare și infecțioase și joacă un rol principal în variabilitatea virușilor, iar enzimele sunt implicate în reproducerea lor.
Virușii structural mai complecși, pe lângă proteine \u200b\u200bși acizi nucleici, conțin carbohidrați și lipide. Fiecare grup de viruși are propriul set de proteine, grăsimi, carbohidrați și acizi nucleici. Unele virusuri conțin enzime în compoziția lor.
Spre deosebire de celulele vii obișnuite, virușii nu consumă alimente sau produc energie. Nu sunt capabili să se reproducă fără participarea unei celule vii. Virusul începe să se înmulțească numai după ce intră într-un anumit tip de celulă. Virusul poliomielitei, de exemplu, poate trăi numai în celulele nervoase ale oamenilor sau în animale foarte organizate, cum ar fi maimuţă. Virușii bacterieni au o structură ușor diferită.
Interacțiunea unui virus cu o celulăVirușii din afara celulei sunt cristale, dar când intră în celulă, „prind viață”. Reproducerea lor are loc într-un mod special, incomparabil. În primul rând, virionii intră în celulă și se eliberează acizi nucleici virali. Apoi detaliile viitorilor virioni sunt „recoltate”. Reproducerea se încheie cu asamblarea de noi virioni și eliberarea lor în mediu.
Întâlnirea virușilor cu celulele începe cu adsorbția sa, adică atașarea la peretele celular. Apoi începe introducerea sau pătrunderea virionului în celulă, pe care ea însăși o realizează. Cu toate acestea, de regulă, pătrunderea virusului în citoplasma celulei este precedată de legarea acestuia cu o proteină receptoră specială situată pe suprafața celulei. Legarea de receptor se realizează datorită prezenței unor proteine \u200b\u200bspeciale pe suprafața particulei virale, care „recunosc” receptorul corespunzător de pe suprafața celulei sensibile. Zeci și chiar sute de virioni pot fi adsorbiți pe o singură celulă. Zona suprafeței celulare, de care este atașat virusul, se aruncă în citoplasmă și se transformă într-un vacuol. Vacuolul, al cărui perete este format dintr-o membrană citoplasmatică, poate fuziona cu alte vacuole sau cu nucleul. Acesta este modul în care virusul este livrat în orice parte a celulei. Acest proces se numește viropexis.
Procesul infecțios începe atunci când virușii care au intrat în celulă încep să se înmulțească, adică se produce reduplicarea genomului viral și auto-asamblarea capsidei. Pentru ca reduplicarea să apară, acidul nucleic trebuie eliberat din capsidă. După sinteza unei noi molecule de acid nucleic, aceasta este îmbrăcată și se formează o capsidă din proteinele virale sintetizate în citoplasma celulei. Acumularea de particule virale duce la ieșirea lor din celulă. Pentru unii viruși, acest lucru se întâmplă prin „explozie”, ca urmare a căreia integritatea celulei este încălcată și aceasta moare. Alte virusuri sunt secretate într-un mod similar celui în devenire. În acest caz, celulele corpului își pot menține viabilitatea pentru o lungă perioadă de timp.
Un alt mod de penetrare în celula în bacteriofagii. Pereții celulari groși nu permit proteinei receptorului, împreună cu virusul atașat, să se scufunde în citoplasmă, așa cum se întâmplă atunci când celulele animale sunt infectate. Prin urmare, bacteriofagul introduce o tijă goală în celulă și împinge prin ea ADN-ul (sau ARN-ul) care se află în capul său. Genomul bacteriofagului intră în citoplasmă, în timp ce capsida rămâne în exterior. În citoplasma celulei bacteriene începe replicarea genomului bacteriofagului, sinteza proteinelor sale și formarea capsidei. După o anumită perioadă de timp, celula bacteriană moare, iar particulele de fagi maturi sunt eliberate în mediu.
Este uimitor modul în care virușii, care sunt de zeci și chiar sute de ori mai mici decât celulele, gestionează cu pricepere și încredere economia celulară. Reproducând, acestea epuizează resursele celulare și adânc, adesea ireversibil, perturbă metabolismul, care în cele din urmă provoacă moartea celulară.
Forma virusurilor de plante este, în general, în formă de tijă și rotund. Dimensiunile virușilor în formă de tijă sunt de 300-480 x 15 nm, iar cei care au o formă rotunjită sunt de 25-30 nm. [...]
Acestea sunt microorganisme care nu au o structură celulară. Dimensiunile unităților structurale ale virușilor (virioni) variază de la 10 la 300 nm. Virionii conțin molecule de acizi ribonucleici (ARN) sau dezoxiribonucleici (ADN), înconjurați de o coajă proteică. Virușii au o varietate de forme: cubice, sferice, în formă de tijă etc. Reproducerea virușilor se realizează prin divizare simplă sau într-un mod mai complex numai în interiorul celulelor unui organism viu. Virușii au specificitate de acțiune, adică anumite grupuri de viruși infectează anumite organisme vii. [...]
Virușii, care sunt mai mici și mai puțin complexi decât celulele, nu pot trăi independent. Sunt doar bucăți de informații genetice foarte ambalate, care pot trăi și se pot reproduce numai infectând o celulă. În acest caz, mii de particule virale se pot forma într-o singură celulă. Se presupune că virușii subjugă cumva mecanismul vieții celulare și îl folosesc în scopuri proprii. Originile evolutive ale virușilor nu sunt pe deplin clare. Ele pot fi considerate ca fiind celule foarte generate sau fragmentele lor. Genele virușilor sunt similare cu genele altor forme și pot fi, de asemenea, mutate. [...]
Acest virus conține aproximativ 20% ARN, iar particulele sale sunt poliedre. Diametrul particulelor din micrografiile electronice obținut prin contrast negativ este de la 26 la 30 nm, care depinde de caracteristicile preparării preparatului. Structura detaliată a acestui virus nu este clară, dar prezintă un interes considerabil, deoarece unele izolate conțin un virus satelit asociat cu acest virus, descris mai jos, precum și în [...]
O altă formă de dependență este caracteristică virusului satelit al virusului necrozei tutunului. Este cel mai mic virus cunoscut. ARN-ul său conține cantitatea de informații suficientă pentru a codifica propria proteină capsidală și, eventual, o polimorază RIK specifică. Pentru alte funcții semnificative, dar până acum necunoscute, op depinde de prezența unui virus necroză de tutun fără legătură. [...]
Virionul virusului este sub formă de fir, are o dimensiune de 600-700 X 12 microni, inactivat la 60-67 ° C, rezistă la îngheț. Transportatorul este necunoscut. [...]
Particulele virusului mozaicului lucernă (ALV) diferă de alte virusuri vegetale prin forma lor baciliformă. Structura acestor viruși are câteva trăsături caracteristice atât virusurilor în formă de tijă, cât și virusurilor izometrice. Cinci componente (b0, 1a, bb, M și B) au fost izolate din preparatul viral VML. Cel puțin patru dintre aceștia s-au dovedit a fi necesari pentru apariția infecției (cap. [...]
Mărimea și forma microbilor. Dimensiunea bacteriilor variază de la zecimi de micron la câțiva microni. În medie, diametrul corpului majorității bacteriilor este cuprins între 0,5-1 microni, iar lungimea medie a bacteriilor în formă de tijă este de 1-5 microni. Rezoluția microscoapelor bacteriologice moderne este de 0,2 microni. Prin urmare, pentru a vedea ultramicrobii (viruși, bacteriofagi), trebuie să utilizați un microscop electronic, care poate crește volumul cu un factor de milioane și are o rezoluție de 0,4 mm. . [...]
Virușii singuri de un fel sau altul sunt formațiuni de diferite forme (rotunde, în formă de tijă sau alte forme), în interiorul cărora există un acid nucleic (ADN sau ARN) închis într-o coajă proteică (capsidă). [...]
ÎN vedere generala virusurile sunt formațiuni submicroscopice formate din proteine \u200b\u200bși acid nucleic și organizate sub formă de particule virale, adesea numite corpusculi virali, virioni, virospori sau nucleocapside. [...]
În unele micrografii electronice, au observat particule discoide de aproape același diametru ca și virusul intact. În aceste particule, era vizibil un canal central, al cărui diametru variază, înconjurat de 10 subunități amplasate radial. [...]
Formele filtrabile de bacterii diferă de virusurile filtrabile prin faptul că se pot dezvolta și pe medii nutritive artificiale. [...]
Bacteriile conțin 1-4% grăsimi, 8-14% proteine \u200b\u200bși 80-85% apă. Urmele conțin fosfor, potasiu, calciu, magneziu, fier și alte elemente. Virușii nu au o structură celulară și au o dimensiune de 10-100 nm. [...]
Agentul cauzal al bolii este virusul mozaicului galben Beap (virusul Phaseolus 2 Smith). Virusul este inactivat la o temperatură de 70 ° C. Afectează toate plantele leguminoase, nu poate fi transmis cu semințe. [...]
Agenții cauzali sunt virusul mozaic ara-bis și virusul inelului de zmeură. Ambii virusuri aparțin aceluiași grup, au particule izometrice cu un diametru de aproximativ: Yu im. Acestea sunt transferate prin mijloace mecanice de contact, nematode ale solului și altoire. Pe frunze sunt pete verzi deschise sau gălbui, de diferite dimensiuni și forme cu margini indistincte. Frunzele sunt mici, deformate, plantele sunt deprimat. În caz de daune grave, plantele din soiuri sensibile mor în decurs de un an. [...]
NOLOGIE pentru extragerea bacteriilor, virușilor și poluanților chimici din apă ", care constă în faptul că microorganismele care trec prin absorbantul de celuloză" se lipesc "în structura sorbentului datorită interacțiunii electrostatice." Ca urmare, „apa devine 100% dezinfectată de viruși, 100% de aproape toate bacteriile și 95-100% de bacteriile E. coli. Impuritățile sunt extrase din apă într-un mod complex: acest lucru se datorează reținerii mecanice a particulelor în structura poroasă a materialului filtrant, datorită sorbției moleculare, interacțiunii electrostatice și schimbului de ioni. " Personal, nu văd nimic PRINCIPAL NOU în această tehnologie, dar există un moment original în Sursa care dă viață. Citez: „Forma părții superioare a filtrului sub forma unei cupole a bisericii are un efect energetic și psihologic benefic asupra persoanelor care beau apă purificată”. Urmează apoi un tabel care compară „Darea vieții” cu tot felul de „acvafore” și „instapuri” (sic), cu care el, desigur, își șterge nasul. [...]
Bacteriofagii și virușii filtrabili nu au structura celulară obișnuită, prin urmare, o celulă organizată nu este ultima unitate de viață. Acest lucru este confirmat de faptele tranziției formelor vizibile de bacterii la „invizibile”, forme necelulare, numite forme filtrabile ale bacteriilor vizibile. [...]
Mozaic de lucernă. Agentul cauzal este virusul mozaic al lucernei (virusul Medicago 2 Smith). Se transmite prin mijloace mecanice de contact, afide, semințe. Simptome: mai întâi, pe frunze apar mici pete rotunjite gălbui, apoi pete alungite sau neregulate între venele laterale, linii galbene deschise sau albicioase de-a lungul venelor. Frunzele sunt mici și deformate. Vara, simptomele sunt adesea mascate. Infecția latentă nu este neobișnuită. Virusul are o gamă largă de gazde: infectează plante sălbatice și cultivate din multe familii: molii, solanacee, astrovacee, dovleac etc. [...]
Pe lângă organismele cu structură celulară, există și forme de viață necelulare - viruși și bacteriofagi. Apropo, virușii au fost descoperiți în 1892 de biologul rus D.I. Ivanov și numele lor în traducere înseamnă „otravă”, ceea ce, în general, în viața de zi cu zi pentru mulți oameni reflectă impactul lor asupra sănătății. [...]
Nu există o limită clară între substanțele vii și cele nevii, lucru confirmat de existența virușilor. Acestea din urmă au semne atât de vii, cât și de nevii. O definiție general acceptată pentru acestea nu a fost încă formulată. De obicei, se crede că virușii sunt cele mai puțin organizate forme de viață care nu au propriul lor metabolism și pot exista doar în interiorul celulelor altor organisme. Nu se înmulțesc în afara celulelor. În același timp, capacitatea virusurilor de a se reproduce, chiar dacă este în contact cu alte celule, este un semn al viețuitoarelor. [...]
Solul conține diverse microorganisme: bacterii, actinomicete sau ciuperci radiante, ciuperci, viruși etc. Majoritatea procesează gunoiul forestier (stratul de humus), îmbunătățesc structura solului, transformă compușii organici în forme digerabile. Odată cu creșterea acidității solului și formarea formelor solubile de metale toxice, activitatea microorganismelor, în special în procesarea deșeurilor forestiere, scade. [...]
Efectul inactivant al produselor de electroliză și al clorului este foarte influențat de cantitatea și forma de clor rezidual (liber sau legat). Studiul dinamicii inactivării produselor de electroliză și a clorului virusului poliomielitei model, fagului Escherichia coli și Escherichia coli a arătat că, în prezența clorului rezidual doar într-o stare legată, la 30 de minute de contact, Escherichia coli a murit complet, iar virusul fagului - doar cu 80 și respectiv 60%. Cu urme de clor rezidual liber, după 20 de minute de contact, E. coli și fagul au fost inactivate cu mai mult de 99%, iar virusul a fost inactivat doar cu 90%. Când conținutul de clor rezidual liber din apă este de 0,1-0,3 mg / l, după 10 minute de contact, acesta a fost complet dezinfectat în raport cu E. coli și fag și, după 30 de minute, a fost detectată doar o cantitate nesemnificativă de viruși activi. Diferența dintre gradul de inactivare a microorganismelor studiate a fost semnificativă statistic în toate cazurile. În condițiile testate de dezinfecție cu produse de electroliză și clor de apă care conține microorganisme în concentrații egale, Escherichia coli a fost mai puțin rezistentă decât fagul, iar fagul a fost mai puțin rezistent decât virusul. Prin urmare, E. coli și fagul pot servi drept indicatori sanitari fiabili ai dezinfecției eficiente a apei cu produse de electroliză și clor împotriva enterovirusurilor. Practic, aceasta se referă la acele cazuri în care, din cauza condițiilor sanitare epidemice nefavorabile, concentrația enterovirusurilor în apa rezervoarelor poate crește semnificativ și poate atinge nivelul de E. coli (E. L. Lovtsevich, L. A. Sergunina, 1968). [...]
Deci, după inventarea antibioticelor, principalul dușman al omului nu a fost cel mai simplu ciuperci și unicelulare, ci virușii. Există primele simptome ale retrovirusurilor care vin să înlocuiască virusurile - forme de viață pre-virale, mai vechi, construindu-și organizarea nu pe baza unei molecule de ADN, ci pe baza ARN. Unul dintre cei mai renumiți reprezentanți ai acestei forme de viață este retrovirusul SIDA. [...]
Microorganismele invizibile la microscop se numesc ultra microbi. Din acest grup de forme ultra-microscopice, bacteriofagii - viruși filtrabili și forme invizibile de bacterii - sunt cele mai importante în practica umană. A fost posibil să se observe ultramicrobii doar la microscopul electronic, ceea ce dă o creștere de până la 45.000 de ori. Virușii (Fig. 85) sunt particule compuse din proteine \u200b\u200bși acid nucleic (ADN sau ARN). Nu au structura celulară obișnuită. Forma necelulară a vieții include și bacteriofagi (Fig. 86), care sunt formațiuni alungite cu un capăt îngroșat. [...]
Procesul infecțios este un complex de reacții într-un macroorganism care apar ca răspuns la introducerea și reproducerea microbilor, virușilor etc. în el. Nu este întotdeauna însoțit de prezența semnelor bolii. De exemplu, cu un microburt sau cu un curs asimptomatic de infecție, nu există semne clinice, deși agentul său patogen este prezent în organism și afectează diferitele sale sisteme, provocând restructurarea imunologică a acestuia din urmă. Dacă procesul infecțios este însoțit de manifestarea semnelor clinice, atunci această formă de infecție se numește boală infecțioasă. Prin urmare, o boală infecțioasă este așa-numita formă manifestă de infecție. [...]
Acești viruși se caracterizează prin similitudine în caracteristicile morfologice, reacții la radiațiile electromagnetice, reproducere etc. Componentele lor principale sunt: \u200b\u200bC, H, N, P, O, carbohidrați și lipide. Se știe că Prak-Pries și toate oncovirusurile sunt instabile termic și sunt distruse la temperaturi de la 50 la 70 ° C, în funcție de tipul de oncovirus. [...]
Acest grup include particule coloidale (minerale și organominere) de soluri și soluri, precum și forme nedisociate și insolubile de substanțe humice care conferă culoare apei. Acestea din urmă sunt spălate în corpuri de apă naturale din soluri de pădure, mlaștină și turbă și se formează, de asemenea, în corpurile de apă, ca urmare a vieții plantelor acvatice și a algelor. Acest grup poate include, de asemenea, viruși și alte organisme care se apropie ca dimensiune de particule coloidale. Deoarece printre ele există organisme patogene (patogene), îndepărtarea lor din apă este un eveniment foarte responsabil. [...]
Al doilea grup de impurități combină particule coloidale hidrofile și hidrofobe minerale și organominerale din soluri și soluri, forme nedisociate și insolubile de substanțe humice și detergenți cu greutate moleculară mare. Stabilitatea cinetică a impurităților hidrofobe se caracterizează prin raportul dintre forțele câmpului gravitațional și mișcarea browniană; stabilitatea lor agregată se datorează stării electrostatice a suprafeței interfazice și formării straturilor difuze sau creării de straturi stabilizatoare pe suprafața particulelor. Acest grup include, de asemenea, viruși și alte microorganisme care au dimensiuni similare particulelor coloidale. [...]
Metoda de centrifugare a gradientului de densitate a lui Bracke poate fi utilizată atât pentru izolarea, cât și pentru cuantificarea virusurilor vegetale. După cum sa dovedit, această metodă este plină de multe posibilități și este utilizată în prezent pe scară largă în domeniul virologiei și biologiei moleculare. Când se efectuează studii prin centrifugare într-un gradient de densitate, tubul centrifugii este parțial umplut cu o soluție, a cărei densitate scade în direcția de la fund la menisc. Zaharoza este cea mai frecvent utilizată pentru a crea un gradient în fracționarea virusurilor vegetale. Înainte de începerea centrifugării, particulele de virus pot fi fie distribuite pe tot volumul soluției, fie aplicate în partea de sus a gradientului. Bracke a propus trei metode diferite de centrifugare cu gradient de densitate. Cu centrifugarea izopicpică (de echilibru), procesul continuă până când toate particulele din gradient ating un nivel în care densitatea mediului este egală cu propria densitate. Astfel, fracționarea particulelor are loc în acest caz în conformitate cu diferențele de densitate a acestora. Soluțiile de zaharoză nu sunt suficient de dense pentru separarea izopicnică a multor viruși. În centrifugarea zonală de mare viteză, virusul este aplicat mai întâi pe un gradient creat anterior. Particulele de fiecare tip sunt sedimentate în același timp printr-un gradient sub forma unei zone sau a unei benzi, la o viteză în funcție de mărimea, forma și densitatea lor. În acest caz, centrifugarea se termină în timp ce particulele continuă să sedimenteze. Centrifugarea zonală de echilibru este similară cu centrifugarea zonală de mare viteză, dar în acest caz centrifugarea continuă până la atingerea stării izopicnice. Rolul gradientului de densitate în centrifugarea de mare viteză este de a inhiba convecția și de a fixa diferite tipuri de molecule în zone specifice. Teoria centrifugării cu gradient de densitate este complexă și nu este bine înțeleasă. Cu toate acestea, în practică, aceasta este o metodă simplă și elegantă, care este utilizată pe scară largă atunci când se lucrează cu virusuri vegetale. [...]
Principala caracteristică a AHC CEC localizată în matrice (precum și a oncovirusurilor din celulă) este prezența unei interfețe între două medii cu conductivitate diferită. În fig. 2.11 arată datele microscopiei electronice care prezintă adenovirusul, virusul Epstein-Barr (EBV) și ЦЭЧ în ShchGK. Fig. 2.11 se poate observa că toate formațiunile de aceeași scară, au o formă apropiată de sferică, constând dintr-un miez și o coajă, în compoziție chimică fiecare coajă conține ioni activi electric, limite clar marcate pentru viruși și CEC cu matricile lor. [...]
Apare pe frunze la începutul primăverii ca o pată galbenă. Până la mijlocul verii, acest simptom dispare, dar frunzele afectate uneori se încrețesc. Fructele sunt mici, adesea neregulate și cu tuberculi de-a lungul cusăturii. Maturizarea lor este întârziată. Agentul cauzal al bolii - virusul mozaicului piersic se transmite prin vaccinare și înmugurire. Se crede că afidele din prune poartă virusul. [...]
În ciuda numeroaselor descoperiri, există încă multe pete goale în imaginea biogenezei. Doar etapele majore pot fi considerate incontestabile. Astfel, nu există nicio îndoială acum că apariția biosferei a fost un eveniment unic, excepțional. Un virus mic și un monstru gigant al mării, o algă unicelulară și o ferigă care a dispărut cu milioane de ani în urmă, sunt toate doar ramuri și frunze pe același copac filogenetic. Formele de viață întotdeauna și peste tot dezvăluie, ca să spunem așa, „rudenie de sânge” și toți copiii ei sunt înrudiți genetic. Încă din ziua când a apărut prima creatură pe Pământ, viața provine doar din viață. [...]
Celula este unitatea structurală și funcțională de bază a tuturor organismelor vii, un sistem viu elementar. Poate exista ca un organism separat (bacterii, protozoare, unele alge și ciuperci) și ca parte a țesuturilor organismelor multicelulare. Numai virușii sunt forme de viață necelulare. [...]
Conform schemei propuse, prima etapă a procesului este formarea complexului enzimă-substrat EI al endonucleazei de restricție EcoR I și a ADN-ului plasmidic dublu catenar. Punctul cheie al schemei este formarea unui complex E-II al restricției endonucleazei EcoR I cu o formă circulară de ADN care conține o ruptură monocatenară, obținută ca rezultat al hidrolizei unei legături fosfodiesterice într-una din firele de ADN. În plus, în funcție de condiții (natura substratului, temperatura etc.) ) fie scindarea celei de-a doua catene de ADN din cadrul aceluiași complex E - II poate apărea odată cu formarea unui complex al enzimei E-III cu o formă liniară de ADN sau disocierea complexului E-P cu formarea unei enzime libere și a ADN-ului circular care conține o rupere cu o singură catena, ceea ce duce la acumularea formei II în soluție. Această schemă a permis explicarea diferențelor în mecanismele de hidroliză a ADN-ului virusului SV 40 pe de o parte și a ADN-ului ColE I și a bacteriofagului G4 pe de altă parte. În cazul ADN-ului virusului SV 40, are loc disocierea complexului enzimă-substrat E-11, ceea ce duce la acumularea formei circulare de ADN în soluție. S-a sugerat că diferențele în mecanismul de hidroliză al acestor molecule de ADN (virusul SV 40; ADN ColE I și bacteriofagul G4) sunt rezultatul interacțiunii enzimei de restricție EcoR I cu diferite secvențe de nucleotide care flancează situl de recunoaștere a enzimei de restricție EcoR I. Cu toate acestea, această ipoteză nu permite explicarea diferențelor în mecanismul de hidroliză al ADN-ului circular ColE I în funcție de temperatură (vezi mai sus). [...]
Boala este cunoscută în multe țări ale lumii. În URSS, a fost găsit în Ucraina, Moldova, Estonia și Georgia și face obiectul unei carantine interne. Sunt afectate pruna, cireașa, mirabela, caisul și piersica. Agentul cauzal al bolii este varicela de prune (\u003d virusul Prunus 7 Smith). Forma virusului este sub formă de fir, dimensiunea 760X20 numită după [...]
Dezvoltarea ulterioară a cercetării privind studiul mecanismului de hidroliză a ADN-ului plasmidic prin endonucleaze de restricție primite în lucrările lui Halford și colab. ... Astfel, mecanismul de reacție este similar cu cel propus pentru hidroliza ADN-ului virusului EU 40 de către enzima de restricție EcoI I. [...]
Pe lângă funcțiile „generale” de mai sus, prezența homeostaziei organismului, există o altă trăsătură foarte importantă: materia vie, ca să spunem așa, creează un alt habitat, și anume posibilitatea ca organismul să se stabilească cu alte ființe vii pentru locuirea permanentă sau temporară. Este un nou habitat biotic creat de viață. Mulți experți consideră că virușii sunt creaturile care locuiesc în acest mediu. Deci, I.A. Shilov (2000) consideră că simplitatea excepțională a structurii lor este un fenomen secundar, și mai probabil este o formă recent apărută de ființe vii care a asimilat complet mediul intracelular în organismele de alte niveluri. A doua confirmare a acestei teze este că virusurile au un grad ridicat de complexitate și diversitate a sistemului genetic. Simplificarea structurii, care a devenit posibilă datorită conexiunilor necondiționate obligatorii ale virușilor cu organismul-gazdă, oferind condiții de viață stabile, a afectat chiar și proprietățile fundamentale inerente majorității covârșitoare a formelor de viață: virușii nu au iritabilitate și nu au aparatul propriu de sinteză a proteinelor. Virușii nu sunt capabili de existență independentă, iar conexiunea lor cu celula nu este doar o conexiune spațială, ci și o conexiune funcțională rigidă cu care celula și virusul reprezintă un fel de unitate. [...]
Tratamentul alcalin pe termen scurt al HPLC la 30 ° C și rezistența ionică ridicată in situ provoacă pauze, ducând la formarea de fragmente de ARN, de dimensiuni destul de uniforme, în care s2 [...]
Numărul persoanelor afectate de malarie, hepatită, HIV și multe alte boli este enorm. Mulți medici cred că nu ar trebui să vorbim despre „victorie”, ci doar despre succesul temporar în lupta împotriva acestor boli. Istoria luptei boli infecțioase este foarte scurt, iar imprevizibilitatea schimbărilor în mediu (în special în cele urbane) poate anula aceste câștiguri. Din acest motiv, „revenirea” agenților infecțioși este înregistrată printre viruși. Mulți viruși se „desprind” de baza naturală și se mută într-o nouă etapă care poate trăi în mediul uman - devin agenți cauzali ai gripei, cancerului viral și a altor boli. Poate că această formă este HIV. [...]
Modificarea greutății moleculare medii a greutății și a razei de rotație a fost evaluată pe baza datelor de împrăștiere a luminii. Pentru ARN HPMC, folosind ambele metode de degradare, au descoperit că raza de rotație a crescut înainte de apariția degradării intense a moleculei, în timp ce raza de rotație și greutatea moleculară medie a ARN-ului TMV a scăzut chiar de la începutul acestui proces. Strazielli și colab. Au explicat această constatare sugerând că ARN-ul HPMC există sub forma unei bucle închise. Cu toate acestea, aceste rezultate pot fi interpretate într-un mod diferit. De exemplu, Hazelkorn a arătat că ARN-ul TMV și ARN-ul HPMT au fost sedimentate împreună în condiții de pH și putere ionică similare cu cele utilizate de Strazielli și colab. Spre deosebire de aceasta, formele circulare și lichge ale ADN-ului fagului cpX174 sunt ușor de distins prin proprietățile lor de sedimentare [1.515]. Kuiper, pe baza datelor de sedimentare în diferite condiții, a sugerat că ARN izolat al virusului mozaicului castravetelui (tulpina Y) poate exista sub două forme: un lanț deschis și o structură circulară. Cu toate acestea, aceste date, precum și în cazul descris mai sus, pot fi explicate în moduri diferite. [...]
ADN-ul bacterian este un compus bogat în polimeri, format dintr-un număr mare de nucleotide - polinucleotide cu o greutate moleculară de aproximativ 4 milioane. O moleculă de ADN este un lanț de nucleotide, unde localizarea lor are o secvență specifică. În secvența de aranjare a bazelor azotate, informațiile genetice ale fiecărei specii sunt codificate. Încălcarea acestei secvențe este posibilă cu mutații naturale sau sub influența factorilor mutageni. În acest caz, microorganismul dobândește sau pierde orice proprietate. El a moștenit trăsăturile schimbării, adică apare o nouă formă de microorganism. În toate microorganismele - procariote și eucariote - purtătorii informațiilor genetice sunt acizii nucleici - ADN și ARN. Doar câțiva viruși fac excepție: nu au ADN, iar informațiile ereditare sunt înregistrate sau reflectate numai în ARN.