ВІРУСИ, дрібні збудники інфекційних хвороб. У перекладі з латинської virus означає «отрута, отруйне початок». До кінця 19 ст. термін «вірус» використовувався в медицині для позначення будь-якого інфекційного агента, що викликає захворювання. Сучасне значення це слово набуло після 1892 коли російський ботанік Д.И.Ивановский встановив «фільтрованість» збудника мозаїчної хвороби тютюну (тютюнової мозаїки). Він показав, що клітинний сік з уражених цією хворобою рослин, пропущений через спеціальні фільтри, що затримують бактерії, зберігає здатність викликати те ж захворювання у здорових рослин. П'ять років по тому інший фільтрівний агент - збудник ящуру великої рогатої худоби - був виявлений німецьким бактеріологом Ф.Лёффлером. У 1898 голландський ботанік М.Бейерінк повторив в розширеному варіанті ці досліди і підтвердив висновки Іванівського. Він назвав «Фільтр отруйна початок», що викликає тютюнову мозаїку, «вірусом, що фільтрується». Цей термін використовувався протягом багатьох років і поступово скоротився до одного слова - «вірус».
У 1901 американський військовий хірург У.Рід і його колеги встановили, що збудник жовтої лихоманки також є вірусом, що фільтрується. Жовта лихоманка була першим захворюванням людини, впізнаним як вірусне, однак знадобилося ще 26 років, щоб її вірусне походження було остаточно доведено.
Прийнято вважати, що віруси походять в результаті відокремлення (автономізації) окремих генетичних елементів клітини, які отримали, крім того, здатність передаватися від організму до організму. У нормальній клітині відбуваються переміщення декількох типів генетичних структур, наприклад матричної, або інформаційної, РНК (мРНК), транспозони, інтронів, плазмід. Такі мобільні елементи, можливо, були попередниками, або прабатьками, вірусів.
Чи є віруси живими організмами? У 1935 американський біохімік У.Стенлі виділив в кристалічній формі вірус тютюнової мозаїки, довівши тим самим його молекулярну природу. Отримані результати викликали бурхливі дискусії про природу вірусів: чи є вони живими організмами або просто активованими молекулами? Дійсно, всередині зараженої клітини віруси проявляють себе як інтегральні компоненти більш складних живих систем, але поза клітиною являють собою метаболічно інертні нуклеопротеїнів. Віруси містять генетичну інформацію, але не можуть самостійно реалізувати її, не володіючи власним механізмом синтезу білка. Коли особливості будови і репродукції вірусів виявилися з'ясованими, питання про те, чи є вони живими, поступово втратив своє значення.
БУДОВА ВІРУСІВ
Повноцінна за будовою і інфекційна, тобто здатна викликати зараження, вірусна частка поза клітиною називається вирионом. Серцевина ( «ядро») віріона містить одну молекулу, а іноді дві або кілька молекул нуклеїнової кислоти. Білковий чохол, що покриває нуклеїнових кислот віріона і захищає її від шкідливих впливів навколишнього середовища, називається капсидом. Нуклеїнова кислота віріона є генетичним матеріалом вірусу (його геномом) і представлена \u200b\u200bдезоксирибонуклеїнової кислотою (ДНК) або рибонуклеїнової кислоти (РНК), але ніколи двома цими сполуками відразу. (Хламідії, рикетсії і всі інші «істинно живі» мікроорганізми містять одночасно ДНК і РНК.) Нуклеїнові кислоти найдрібніших вірусів містять три або чотири гени, тоді як найбільші віруси мають до ста генів.
У деяких вірусів на додаток до капсиду є ще й зовнішня оболонка, що складається з білків і ліпідів. Вона утворюється з мембран зараженої клітини, що містять вбудовані вірусні білки. Терміни «голі віріони» і «позбавлені оболонки віріони» використовуються як синоніми. Капсиди найдрібніших і просто влаштованих вірусів можуть складатися лише з одного або декількох видів білкових молекул. Кілька молекул одного або різних білків об'єднуються в субодиниці, звані капсомерами. Капсомери, в свою чергу, утворюють правильні геометричні структури вірусного капсида. У різних вірусів форма капсида є характерною особливістю (ознакою) віріона.
Віріони зі спіральним типом симетрії, як у вірусу тютюнової мозаїки, мають форму подовженого циліндра; всередині білкового чохла, що складається з окремих субодиниць - капсомеров, знаходиться згорнута спіраль нуклеїнової кислоти (РНК). Віріони з ікосаедрічеськая типом симетрії (від грец. Eikosi - двадцять, hedra - поверхня), як у поліовірусу, мають сферичну, а точніше, багатогранну форму; їх капсиди побудовані з 20 правильних трикутних фасеток (поверхонь) і схожі на геодезичний купол.
У окремих бактеріофагів (вірусів бактерій; фагів) змішаний тип симетрії. У т.зв. «Хвостатих» фагів головка має вигляд сферичного капсида; від неї відходить довгий трубчастий відросток - «хвіст».
Зустрічаються віруси з ще більш складною будовою. Віріони поксвирусов (віруси групи віспи) не мають правильного, типового капсида: між серцевиною і зовнішньою оболонкою у них розташовуються трубчасті і мембранні структури.
Генетичну інформацію, закодовану в окремому гені, загалом можна розглядати як інструкцію з виробництва певного білка в клітині. Така інструкція сприймається клітиною тільки в тому випадку, якщо вона послана у вигляді мРНК. Тому клітини, у яких генетичний матеріал представлений ДНК, повинні «переписати» (транскрибувати) цю інформацію в комплементарную копію мРНКвірусних білків (див. Також НУКЛЕЇНОВІ КИСЛОТИ). ДНК-віруси за способом реплікації відрізняються від РНК-вірусів.
ДНК зазвичай існує у вигляді дволанцюжкових структур: дві полінуклеотідниє ланцюжки з'єднані водневими зв'язками і закручені таким чином, що утворюється подвійна спіраль. РНК, навпаки, зазвичай існує у вигляді одноланцюгових структур. Однак геном окремих вірусів являє собою одноцепочечную ДНК або двухцепочечную РНК. Нитки (ланцюжки) вірусної нуклеїнової кислоти, подвійні або одинарні, можуть мати лінійну форму або замикатися в кільце.
Перший етап реплікації вірусів пов'язаний з проникненням вірусної нуклеїнової кислоти в клітину організму-господаря. Цьому процесу можуть сприяти спеціальні ферменти, що входять до складу капсида або зовнішньої оболонки віріона, причому оболонка залишається зовні клітини або віріон втрачає її відразу після проникнення всередину клітини. Вірус знаходить підходящу для його розмноження клітку, контактуючи окремими ділянками свого капсида (або зовнішньої оболонки) зі специфічними рецепторами на поверхні клітини за типом «ключ - замок». Якщо специфічні ( «узнающие») рецептори на поверхні клітини відсутні, то клітина не чутлива до вірусної інфекції: вірус в неї не проникає.
Для того щоб реалізувати свою генетичну інформацію, яка проникла в клітку вірусна ДНК транскрибується спеціальними ферментами в мРНК. Новоутворена мРНК переміщається до клітинних «фабрикам» синтезу білка - рибосоми, де вона замінює клітинні «послання» власними «інструкціями» і транслюється (прочитується), в результаті чого синтезуються вірусні білки. Сама ж вірусна ДНК багаторазово подвоюється (дупліціруется) за участю іншого набору ферментів, як вірусних, так і належать клітці.
Синтезований білок, який використовується для будівництва капсида, і розмножена в багатьох копіях вірусна ДНК об'єднуються і формують нові, «дочірні» віріони. Сформований вірусне потомство залишає використану клітку і заражає нові: цикл репродукції вірусу повторюється. Деякі віруси під час отпочковиванія від поверхні клітини захоплюють частину клітинної мембрани, в яку «завчасно» прилаштувалися вірусні білки, і таким чином набувають оболонку. Що стосується клітини-господаря, то вона в підсумку виявляється пошкодженою або навіть повністю зруйнованою.
У деяких ДНК-вірусів сам цикл репродукції в клітині не пов'язаний з негайною репликацией вірусної ДНК; замість цього вірусна ДНК вбудовується (інтегрується) в ДНК клітини-господаря. На цій стадії вірус як єдине структурне утворення зникає: його геном стає частиною генетичного апарату клітини і навіть реплицируется в складі клітинної ДНК під час поділу клітини. Однак згодом, іноді через багато років, вірус може з'явитися знову - запускається механізм синтезу вірусних білків, які, об'єднуючись з вірусною ДНК, формують нові віріони.
У деяких РНК-вірусів геном (РНК) може безпосередньо виконувати роль мРНК. Однак ця особливість характерна тільки для вірусів з «+» ниткою РНК (тобто з РНК, що має позитивну полярність). У вірусів з «-» ниткою РНК остання повинна спочатку «переписати» в «+» нитка; тільки після цього починається синтез вірусних білків і відбувається реплікація вірусу.
Так звані ретровіруси містять в якості генома РНК і мають незвичайний спосіб транскрипції генетичного матеріалу: замість транскрипції ДНК в РНК, як це відбувається в клітці і характерно для ДНК-вірусів, їх РНК транскрибується в ДНК. Дволанцюжкова ДНК вірусу потім вбудовується в хромосомну ДНК клітини. На матриці такий вірусної ДНК синтезується нова вірусна РНК, яка, як і інші, визначає синтез вірусних білків. Див. Також ретровірус.
Якщо віруси дійсно є мобільними генетичними елементами, які отримали «автономію» (незалежність) від генетичного апарату їх господарів (різних типів клітин), то різні групи вірусів (з різним геномом, будовою і репликацией) повинні були виникнути незалежно один від одного. Тому побудувати для всіх вірусів єдиний родовід, яка б пов'язала їх на основі еволюційних взаємин, неможливо. Принципи «природною» класифікації, що використовуються в систематиці тварин, не підходять для вірусів.
Проте система класифікації вірусів необхідна в практичній роботі, і спроби її створення робилися неодноразово. Найбільш продуктивним виявився підхід, заснований на структурно-функціональну характеристику вірусів: щоб відрізнити різні групи вірусів один від одного, описують тип їх нуклеїнової кислоти (ДНК або РНК, кожна з яких може бути одноцепочечной або двухцепочечной), її розміри (число нуклеотидів в ланцюжку нуклеїнової кислоти), число молекул нуклеїнової кислоти в одному вирионе, геометрію віріона і особливості будови капсида і зовнішньої оболонки віріона, тип господаря (рослини, бактерії, комахи, ссавці і т.д.), особливості спричиненої вірусами патології (симптоми і характер захворювання), антигенні властивості вірусних білків і особливості реакції імунної системи організму на впровадження вірусу.
У систему класифікації вірусів не цілком вкладається група мікроскопічних збудників хвороб, звана віроїди (тобто вірусоподібні частки). Віроїди викликають багато поширених серед рослин хвороби. Це дрібні інфекційні агенти, позбавлені навіть найпростішого білкового чохла (наявного у всіх вірусів); вони складаються тільки з замкнутої в кільце одноцепочечной РНК.
ВЕРЕСНЯ ЗАХВОРЮВАННЯ
Для багатьох вірусів, наприклад кору, герпесу і частково грипу, основним природним резервуаром є людина. Передача цих вірусів відбувається повітряно-краплинним або контактним шляхом.
поширення деяких вірусних захворювань, Як і інших інфекцій, повно несподіванок. Наприклад, в групах людей, що проживають в антисанітарних умовах, практично всі діти в ранньому віці переносять поліомієліт, зазвичай протікає в легкій формі, і набувають імунітету. Якщо ж умови життя в цих групах поліпшуються, діти молодшого віку зазвичай на поліомієліт не хворіють, але захворювання може виникнути в більш старшому віці, і тоді воно часто протікає у важкій формі.
Багато віруси не можуть довго зберігатися в природі при низькій щільності розселення виду-господаря. Нечисленність популяцій первісних мисливців і збирачів рослин створювала несприятливі умови для існування деяких вірусів; тому мабуть, що якісь віруси людини виникли пізніше, з появою міських і сільських поселень. Передбачається, що вірус кору спочатку існував серед собак (як збудник лихоманки), а натуральна віспа людини могла з'явитися в результаті еволюції віспи корів або мишей. До найбільш «свіжим» прикладів еволюції вірусів можна віднести синдром набутого імунодефіциту людини (СНІД). Існують дані про генетичне подібність вірусів імунодефіциту людини і африканських зелених мавп.
«Нові» інфекції зазвичай протікають у важкій формі, нерідко зі смертельним результатом, але в процесі еволюції збудника вони можуть стати більш легкими. Хороший приклад - історія вірусу міксоматозу. У 1950 цей вірус, ендемічний для Південної Америки і досить нешкідливий для місцевих кроликів, разом з європейськими породами цих тварин був завезений до Австралії. Захворювання австралійських кроликів, які раніше не зустрічалися з цим вірусом, було смертельним в 99,5% випадків. Кілька років по тому смертність від цього захворювання значно знизилася, в деяких районах до 50%, що пояснюється не тільки «аттенуірующімі» (ослабляють) мутаціями в вірусному геномі, а й зрослої генетичної стійкістю кроликів до захворювання, причому в обох випадках ефективна природна селекція відбулася під потужним тиском природного відбору.
Репродукція вірусів в природі підтримується різними типами організмів: бактеріями, грибами, найпростішими, рослинами, тваринами. Наприклад, комахи часто страждають від вірусів, які накопичуються в їх клітинах у вигляді великих кристалів. Рослини нерідко уражаються дрібними і просто влаштованими РНК-вмісними вірусами. Ці віруси навіть не мають спеціальних механізмів для проникнення в клітину. Вони переносяться комахами (які харчуються клітинним соком), круглими хробаками і контактним способом, заражаючи рослина при його механічному пошкодженні. Віруси бактерій (бактеріофаги) мають найбільш складний механізм доставки свого генетичного матеріалу в чутливу бактеріальну клітину. Спочатку «хвіст» фага, що має вигляд тонкої трубочки, прикріплюється до стінки бактерії. Потім спеціальні ферменти «хвоста» розчиняють ділянку бактеріальної стінки і в отвір, що утворився через «хвіст», як через голку шприца, впорскується генетичний матеріал фага (зазвичай ДНК).
Понад десять основних груп вірусів патогенні для людини. Серед ДНК-вірусів це сімейство поксвирусов (викликають натуральну віспу, коров'ячу віспу та інші віспяні інфекції), віруси групи герпесу (герпетичні висипання на губах, вітряна віспа), аденовіруси (захворювання дихальних шляхів і очей), сімейство паповавирусов (бородавки та інші розростання шкіри), гепаднавірус (вірус гепатиту B). РНК-вірусів, хвороботворних для людини, значно більше. Пікорнавіруси (від лат. Pico - дуже невеликий, англ. RNA - РНК) - найдрібніші віруси ссавців, схожі на деякі віруси рослин; вони викликають поліомієліт, гепатит А, гострі простудні захворювання. Міксовіруси і параміксовіруси - причина різних форм грипу, кору та епідемічного паротиту (свинки). Арбовіруси (від англ. Arthropod borne - «переносяться членистоногими») - найбільша група вірусів (більш 300) - переносяться комахами і є збудниками кліщового і японського енцефаліту, жовтої лихоманки, менінгоенцефалітів коней, колорадській кліщовий лихоманки, шотландського енцефаліту овець та інших небезпечних хвороб . Реовіруси - досить рідкісні збудники респіраторних і кишкових захворювань людини - стали предметом особливого наукового інтересу в силу того, що їх генетичний матеріал представлений двухцепочечной фрагментованою РНК. Див. Також ВЕНЕРИЧНІ ХВОРОБИ; ВІТРЯНА ВІСПА; ГЕПАТИТ; ГРИП; Денге ЛИХОМАНКА; МОНОНУКЛЕОЗ ІНФЕКЦІЙНИЙ; КІР; КРАСНУХА; МЕНІНГІТ; ВІСПА Натуральна; ПОЛІОМІЄЛІТ; РЕСПІРАТОРНІ ВЕРЕСНЯ ЗАХВОРЮВАННЯ; Свинка; СИНДРОМ набутого імунодефіциту (СНІД); Енцефаліт.
Збудники деяких хвороб, в тому числі дуже важких, не вкладаються ні в одну з перерахованих вище категорій. До особливої \u200b\u200bгрупи повільних вірусних інфекцій ще недавно відносили, наприклад, хвороба Крейтцфельда - Якоба і куру - дегенеративні захворювання головного мозку, що мають дуже тривалий інкубаційний період. Однак виявилося, що вони викликаються не вірусами, а дрібними інфекційними агентами білкової природи - пріонами (див. Пріони).
Лікування і профілактика. Репродукція вірусів тісно переплітається з механізмами синтезу білка і нуклеїнових кислот клітини в зараженому організмі. Тому створити ліки, що вибірково пригнічують вірус, але не завдають шкоди організму, - завдання надзвичайно важка. Все ж виявилося, що у найбільш великих вірусів герпесу і віспи геномні ДНК кодують велике число ферментів, що відрізняються за властивостями від подібних клітинних ферментів, і це послужило основою для розробки противірусних препаратів. Дійсно, створено кілька препаратів, механізм дії яких заснований на придушенні синтезу вірусних ДНК. Деякі сполуки, занадто токсичні для загального застосування (внутрішньовенно або через рот), годяться для місцевого використання, наприклад при ураженні очей вірусом герпесу.
Відомо, що в організмі людини виробляються особливі білки - інтерферони. Вони пригнічують трансляцію вірусних нуклеїнових кислот і таким чином пригнічують розмноження вірусу. Завдяки генної інженерії стали доступні і проходять перевірку в медичній практиці інтерферони, вироблені бактеріями (див. ГЕННА ІНЖЕНЕРІЯ).
До найдієвішим елементам природного захисту організму відносяться специфічні антитіла (спеціальні білки, які виробляються імунною системою), які взаємодіють з відповідним вірусом і тим самим ефективно перешкоджають розвитку хвороби; проте вони не можуть нейтралізувати вірус, вже проник в клітку. Прикладом може служити герпетична інфекція: Вірус герпесу зберігається в клітинах нервових вузлів (гангліїв), де антитіла не можуть його досягти. Час від часу вірус активується і викликає рецидиви захворювання.
Зазвичай специфічні антитіла утворюються в організмі в результаті проникнення в нього збудника інфекції. Організму можна допомогти, посилюючи вироблення антитіл штучно, в тому числі створюючи імунітет заздалегідь, за допомогою вакцинації. Саме таким способом, шляхом масової вакцинації, захворювання натуральною віспою було практично ліквідовано в усьому світі. Див. Також ВАКЦИНАЦІЯ та імунізації.
Сучасні методи вакцинації та імунізації поділяються на три основні групи. По-перше, це використання ослабленого штаму вірусу, який стимулює в організмі продукування антитіл, ефективно діють проти більш патогенного штаму. По-друге, введення убитого вірусу (наприклад, інактивованої формаліном), який теж індукує утворення антитіл. Третій варіант - т.зв. «Пасивна» імунізація, тобто введення вже готових «чужих» антитіл. Тварина, наприклад коня, імунізують, потім з її крові виділяють антитіла, очищають їх і використовують для введення пацієнту, щоб створити негайний, але нетривалий імунітет. Іноді використовують антитіла з крові людини, який переніс дане захворювання (наприклад, кір, кліщовий енцефаліт).
Накопичення вірусів. Для приготування вакцинних препаратів необхідно накопичити вірус. З цією метою часто використовують розвиваються курячі ембріони, яких заражають даним вірусом. Після інкубування заражених ембріонів протягом певного часу накопичився в них внаслідок розмноження вірус збирають, очищають (центрифугуванням або іншим способом) і, якщо потрібно, інактивують. Дуже важливо видалити з препаратів вірусу все баластні домішки, які можуть викликати серйозні ускладнення при вакцинації. Звичайно, не менш важливо переконатися, що в препаратах не залишилося неінактівірованного патогенного вірусу. В останні роки для накопичення вірусів широко використовують різні типи клітинних культур.
МЕТОДИ ВИВЧЕННЯ ВІРУСІВ
Віруси бактерій першими стали об'єктом детальних досліджень як найбільш зручна модель, що володіє рядом переваг в порівнянні з іншими вірусами. Повний цикл реплікації фагів, тобто час від зараження бактеріальної клітини до виходу з неї розмножилися вірусних частинок, відбувається протягом однієї години. Інші віруси зазвичай накопичуються протягом декількох діб або навіть більш тривалого часу. Незадовго до Другої світової війни і незабаром після її закінчення були розроблені методи вивчення окремих вірусних частинок. Чашки з поживним агаром, на якому вирощено моношар (суцільний шар) бактеріальних клітин, заражають частинками фага, використовуючи для цього його послідовні розведення. Розмножуючись, вірус вбиває «прихистила» його клітку і проникає в сусідні, які теж гинуть після накопичення фагового потомства. Ділянка загиблих клітин видно неозброєним оком як світла пляма. Такі плями називають «негативними колоніями», або бляшками. Розроблений метод дозволив вивчати потомство окремих вірусних частинок, виявити генетичну рекомбінацію вірусів і визначити генетичну структуру і способи реплікації фагів в деталях, які здавалися раніше неймовірними.
Роботи з бактеріофагами сприяли розширенню методичного арсеналу в вивченні вірусів тварин. До цього дослідження вірусів хребетних виконувалися в основному на лабораторних тварин; такі досліди були дуже трудомісткі, дорогі і не дуже інформативні. Згодом з'явилися нові методи, засновані на застосуванні тканинних культур; бактеріальні клітини, що використовувалися в експериментах з фагами, були замінені на клітини хребетних. Однак для вивчення механізмів розвитку вірусних захворювань експерименти на лабораторних тваринах дуже важливі і продовжують проводитися в даний час.
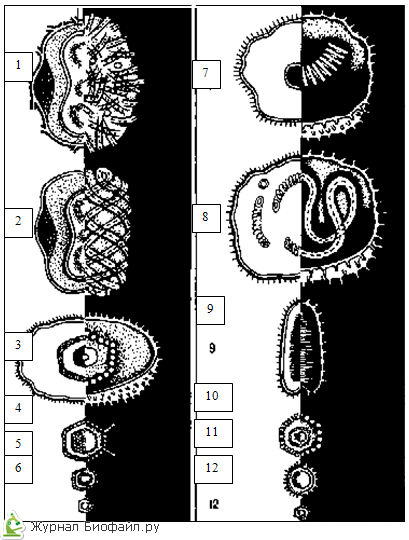
ПРИ ікосаедрічеськая типі симетрії, показаної на схемі будови аденовірусу, капсомери, або білкові субодиниці вірусу, утворюють ізометричний білковий чохол, що складається з 20 правильних трикутників.
У РАЗІ спіральною симетрією, показаної на схемі будови вірусу тютюнової мозаїки, капсомери, або субодиниці вірусу, формують спіраль навколо порожнистої трубчастої серцевини.
Комбінована, або змішана, симетрія у вірусів може бути представлена \u200b\u200bрізними варіантами. Частка бактеріофага, показана на схемі, має «головку» правильної геометричної форми і «хвіст» із спіральною симетрією.
кристалізація вірусів
У 1932 році молодому американському біохіміку Венділлу Стенлі запропонували зайнятися вірусами. Стенлі почав з того, що віджав бутель соку з тонни листя тютюну, уражених вірусом тютюнової мозаїки. Він почав досліджувати сік доступними йому хімічними методами. Різні фракції соку він піддавав впливу всіляких реактивів, сподіваючись отримати чистий вірусний білок (Стенлі був переконаний, що вірус - це білок). Одного разу, Стенлі отримав майже чисту фракцію білка, що відрізнявся за своїм складом від білків рослинних клітин. Вчений зрозумів, що перед ним те, чого він так наполегливо домагався. Стенлі виділив незвичайний білок, розчинив його у воді і поставив розчин в холодильник. На ранок в колбі замість прозорої рідини лежали красиві шовковисті голчасті кристали. З тонни листя Стенлі добув столову ложку таких кристалів. Потім Стенлі відсипав трохи кристаликів, розчинив їх у воді, змочив цією водою марлю і нею натер листя здорових рослин. Сік рослин піддався цілому комплексу хімічних впливів. Після такої "масованої обробки" віруси, швидше за все, повинні були загинути.
Натерті листя захворіли. Отже, дивні властивості вірусу поповнилися ще одним - здатністю кристалізуватися.
Ефект кристалізації був настільки приголомшливим, що Стенлі надовго відмовився від думки, що вірус - це істота. Так як всі ферменти - білки, і кількість багатьох ферментів також збільшується в міру розвитку організму, і вони можуть кристалізуватися, Стенлі уклав, що віруси - чисті білки, швидше за ферменти.
Незабаром вчені переконалися, що кристалізувати можна не тільки вірус тютюнової мозаїки, а й ряд інших вірусів.
Через п'ять років англійські біохіміки Ф. Боуден і Н. Пірі знайшли помилку у визначенні Стенлі.94% вмісту вірусу тютюнової мозаїки складалося з білка, а 6% представляло собою нуклеїнових кислот. Вірус був насправді не білком, а нуклеопротеїн - з'єднанням білка і нуклеїнової кислоти.
Як тільки біологам стали доступні електронні мікроскопи, вчені встановили, що кристали вірусів складаються з тісно притиснутих один до одного декількох сотень мільярдів частинок. В одному кристалі вірусу поліомієліту стільки частинок, що ними можна заразити не по одному разу всіх жителів Землі. Коли ж вдалося розглянути в електронному мікроскопі окремі вірусні частинки, то виявилося що вони бувають різної форми але завжди зовнішня оболонка вірусів складається з білка, які відрізняються у різних вірусів, що дозволяє розпізнавати їх за допомогою імунологічних реакцій, а внутрішній вміст представлено нуклеїнової кислотою, яка є одиницею спадковості.
Складові частини вірусівНайбільші віруси (віруси віспи) наближаються за розмірами до невеликих бактеріям, найдрібніші (збудники енцефаліту, поліомієліту, ящуру) - до великих білкових молекул. Іншими словами, серед вірусів є свої велетні і карлики. (Див. Рис. 1) Для вимірювання вірусів використовують умовну величину, яка називається нанометрів (нм). Один нм становить мільйонну частку міліметра. Розміри різних вірусів варіюють від 20 до 300 нм.
Отже, віруси складаються з декількох компонентів:
серцевина - генетичний матеріал (ДНК або РНК). Генетичний апарат вірусу несе інформацію про декілька типів білків, які необхідні для утворення нового вірусу.
білкова оболонка, яку називають капсидом. Оболонка часто побудована з ідентичних повторюваних субодиниць - капсомеров. Капсомери утворять структури з високим ступенем симетрії.
Додаткова липопротеідна оболонка. Вона утворена з плазматичної мембрани клітини-хазяїна. Вона зустрічається тільки в порівняно великих вірусів (грип, герпес). Ця зовнішня оболонка є фрагментом ядерної або цитоплазматичної мембрани клітини-хазяїна, з якої вірус виходить в позаклітинне середовище. Іноді в зовнішніх оболонках складних вірусів крім білків містяться вуглеводи, наприклад у збудників грипу та герпесу.
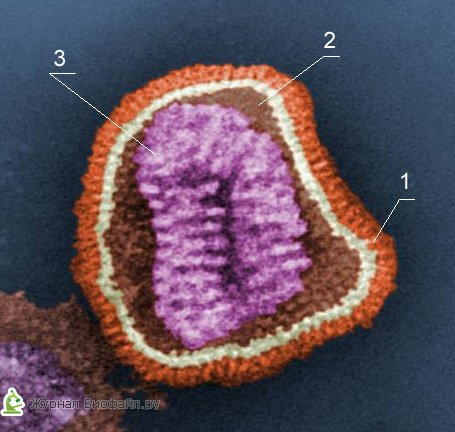
1. Додаткова оболонка
2. капсомеров (білкова оболонка)
3. Серцевина (ДНК або РНК)
Кожен компонент віріонів має певні функції: білкова оболонка захищає їх від несприятливих впливів, нуклеїнова кислота відповідає за спадкові та інфекційні властивості і відіграє провідну роль в мінливості вірусів, а ферменти беруть участь в їх розмноження.
Більш складні за структурою віруси, крім білків і нуклеїнових кислот, містять вуглеводи, ліпіди. Для кожної групи вірусів характерний свій набір білків, жирів, вуглеводів і нуклеїнових кислот. Деякі віруси містять в своєму складі ферменти.
На відміну від звичайних живих клітин віруси не вживають їжі і не виробляють енергії. Вони не здатні розмножуються без участі живої клітини. Вірус починає розмножуватися лише після того, як він проникне в клітку певного типу. Вірус поліомієліту, наприклад, може жити тільки в нервових клітинах людини або таких високоорганізованих тварин, як мавпи. Трохи іншу будову у вірусів бактерій.
Взаємодія вірусу з клітиноюВіруси поза клітиною являють собою кристали, але при попаданні в клітину "оживають". Їх розмноження відбувається особливим, ні з чим не порівнянним способом. Спочатку віріони проникають всередину клітини, і звільняються вірусні нуклеїнові кислоти. Потім "заготовлюються" деталі майбутніх віріонів. Розмноження закінчується складанням нових віріонів і виходом їх в навколишнє середовище.
Зустріч вірусів з клітинами починається з його адсорбції, тобто прикріплення до клітинної стінки. Потім починається впровадження або проникнення віріона в клітку, яке здійснює вона сама. Однак, як правило, проникненню вірусу в цитоплазму клітини передує зв'язування його з особливим білком-рецептором, що знаходяться на клітинній поверхні. Зв'язування з рецептором здійснюється завдяки наявності спеціальних білків на поверхні вірусної частки, які "впізнають" відповідний рецептор на поверхні чутливої \u200b\u200bклітини. На одній клітці можуть адсорбуватися десятки, і навіть сотні віріонів. Ділянка поверхні клітини, до якого приєднався вірус, занурюється в цитоплазму і перетворюється в вакуоль. Вакуоль, стінка якої складається з цитоплазматичної мембрани, може зливатися з іншими вакуолями або з ядром. Так вірус доставляється в будь-яку ділянку клітини. Цей процес називається виропексисом.
Інфекційний процес починається, коли проникли в клітку віруси починають розмножуватися, тобто відбувається редуплікація вірусного генома і самосборка капсида. Для здійснення редуплікації нуклеїнова кислота повинна звільнитися від капсида. Після синтезу нової молекули нуклеїнової кислоти вона одягається, синтезованими в цитоплазмі клітини - вірусними білками - утворюється капсид. Накопичення вірусних частинок призводить до виходу їх з клітини. Для деяких вірусів це відбувається шляхом "вибуху", в результаті чого цілісність клітини порушується і вона гине. Інші віруси виділяються способом, що нагадує брунькування. В цьому випадку клітини організму можуть довго зберігати свою життєздатність.
Інший шлях проникнення в клітину у бактеріофагів. Товсті клітинні стінки не дозволяють білку-рецептора разом з приєдналася до нього вірусом занурюватися в цитоплазму, як це відбувається при інфікуванні клітин тварин. Тому бактеріофаг вводить порожнистий стрижень в клітку і заштовхує через неї ДНК (або РНК), що знаходиться в його голівці. Геном бактеріофага потрапляє в цитоплазму, а капсид залишається зовні. У цитоплазмі бактеріальної клітини починається редуплікація геному бактеріофага, синтез його білків і формування капсида. Через певний проміжок часу бактеріальна клітина гине, і зрілі фагів частинки виходять в навколишнє середовище.
Вражаюче, як віруси, які в десятки і навіть сотні разів менше клітин, вміло, і впевнено розпоряджаються клітинним господарством. Розмножуючись, вони виснажують клітинні ресурси і глибоко, часто незворотньо, порушують обмін речовин, що, в кінцевому рахунку, є причиною загибелі клітин.
Форма вірусів рослин в основному буває палочковидной і округлої. Розміри вірусів палочковидной форми складають 300-480 х 15 нм, а розміри тих, які мають округлу форму, рівні 25-30 нм. [...]
Це - мікроорганізми, які не мають клітинної будови. Розміри структурних одиниць вірусів (віріонів) коливаються від 10 до 300 нм. До складу віріонів входять молекули рибонуклеїнової (РНК) або дезоксирибонуклеїнової (ДНК) кислот, оточені білковою оболонкою. Віруси мають різноманітну форму: кубічну, сферичну, паличкоподібну і ін. Розмноження вірусів здійснюється простим поділом або більш складним шляхом тільки всередині клітин живого організму. Віруси володіють специфічністю дії, т. Е. Окремі групи вірусів вражають певні живі організми. [...]
Віруси, що мають більш дрібні розміри і менш складну структуру, ніж клітини, не можуть жити незалежно. Вони всього лише дуже своєрідно упаковані частки генетичної інформації, здатні жити і розмножуватися тільки інфікувавши якусь клітину. При цьому в одній клітці можуть утворитися тисячі вірусних частинок. Припускають, що віруси якимось чином підпорядковують собі механізм життєдіяльності клітини і використовують його в своїх цілях. Походження вірусів в процесі еволюції не зовсім ясно. Їх можна розглядати як сильно дсгенерірованние клітини або їх фрагменти. Гени вірусів подібні генам інших форм і також можуть піддаватися мутації. [...]
Цей вірус містить близько 20% РНК, і його частинки мають форму багатогранника. Діаметр частинок на електронних мікрофотографіях, отриманих методом негативного контрастування, становить від 26 до 30 нм, що залежить від особливостей приготування препарату. Детальна структура цього вірусу -не з'ясована, але він представляє значний інтерес, оскільки деякі ізоляти містять пов'язаний з цим вірусом вірус-сатолліт, описаний нижче, а також в гл. [...]
Інша форма залежності характерна для вірусу-сателіта вірусу некрозу тютюну. Це найдрібніший з відомих вірусів. У його РНК міститься кількість інформації, достатню для кодування власного капсидних білка і, можливо, специфічної РЙК-поліморази. Щодо інших істотних, але поки невідомих функцій оп залежить від присутності нерідного йому вірусу некрозу тютюну. [...]
Віріон вірусу ниткоподібної форми, розміром 600-700 X 12 мкм, інактивується при 60-67 ° С, витримує промороження. Переносник невідомий. [...]
Частинки вірусу мозаїки люцерни (ВМЛ) відрізняються від інших вірусів рослин своєї бацілловідной формою. Структурі цих вірусів притаманні деякі особливості, характерні як для палочкообразной, так і для ізометричних вірусів. З вірусного препарату ВМЛ було виділено 5 компонентів (Ь0, 1а, Ьь, М і В). Принаймні чотири з них виявилися необхідними для виникнення інфекції (гл. [...]
Розміри і форма мікробів. Розміри бактерій коливаються в межах від десятих часток мікрона до декількох мікрон. В середньому діаметр тіла більшості бактерій знаходиться в межах 0,5-1 мк, а середня довжина становить у паличковидних бактерій 1-5 мк. Роздільна здатність сучасних бактеріологічних мікроскопів дорівнює 0,2 мк. Тому щоб побачити ультрамікробів (віруси, бактеріофаги), потрібно використовувати електронний мікроскоп, що дозволяє збільшити об'єм в мільйони разів і має роздільну здатність 0,4 ммк. . [...]
Поодинокі віруси тих чи інших видів являють собою утворення різної форми (округлої, палочковидной або іншої форми), всередині яких міститься нуклеїнова кислота (ДНК або РНК), укладена в білкову оболонку (капсид). [...]
В загалом вигляді віруси являють собою субмікроскопічес-кі освіти, що складаються з білка і нуклеїнової кислоти і організовані у формі вірусних частинок, що їх називають вірусними корпускулами, вирионами, віроспори або нуклеокап-сидами. [...]
На деяких електронних мікрофотографіях вони спостерігали дисковидні частки майже такого ж діаметру, що і інтактньтй вірус. У цих частинках було видно центральний канал, діаметр якого варіював, оточений 10 радіально розташованими субодиницями. [...]
Фільтрівні форми бактерій відрізняються від фільтруються вірусів тим, що вони можуть розвиватися і на штучних поживних середовищах. [...]
До складу бактерій входить 1-4% жирів, 8 - 14% білків і 80- 85% води. У микроколичествах містяться фосфор, калій, кальцій, магній, залізо та інші елементи. Віруси не володіють клітинною структурою і мають розмір 10 100 нм. [...]
Збудник хвороби - вірус жовтої мозаїки квасолі Веап yellow mosaic virus (Phaseolus virus 2 Smith). Інактивується вірус при температурі 70 ° С. Вражає все бобові рослини, з насінням не передається. [...]
Збудники - вірус мозаїки резуха (ara-bis mosaic virus) і вірус кільцевої плямистості малини (raspberry ringspol virus). Обидва віруси відносяться до однієї групи, мають ізометричні частинки діаметром близько: Ю ім. Переносяться контактно-механічним шляхом, грунтовими нематодами і щепленням. На листі світло-зелені або жовтуваті плями, різної величини і форми з нечіткими краями. Листя дрібні, деформовані, рослини пригнічені. При сильному ураженні рослини сприйнятливих сортів гинуть протягом року. [...]
Нології вилучення бактерій, вірусів і хімічних забруднювачів з води », яка полягає в тому, чю мікроорганізми, що проходять через целюлозний сорбент,« влипають »в структуру сорбенту за рахунок електростатичного взаємодії». В результаті «вода стає стерильною від вірусів на 100%, майже від всіх бактерій - на 100%, і від бактерій кишкової палички - на 95-100%. Домішки витягуються з води складним шляхом: це відбувається за рахунок механічного утримання часток в пористій структурі фільтрувального матеріалу, за рахунок молекулярної сорбції, електростатичного взаємодії та іонного обміну ». Особисто я нічого ПРИНЦИПОВО НОВОГО в цій технології не вбачаю, але є в «Живоносному джерелі» один оригінальний момент. Цитую: «Форма верхньої частини фільтра у вигляді купола церкви благотворно енергетичне і психологічний вплив на людей, що п'ють очищену воду». Потім слід таблиця порівняння «Живоносного» з усякими «Аквафор» і «ін-стапурамі» (так в оригіналі), яким він, зрозуміло, втирає носа. [...]
Бактеріофаги і фільтруються віруси не мають звичайної клітинної структурою, отже, організована клітина не є останньою одиницею життя. Це підтверджується фактами переходу видимих \u200b\u200bформ бактерій в «невидимі», неклітинні форми, Що отримали назву фільтруються форм видимих \u200b\u200bбактерій. [...]
Мозаїка люцерни. Збудник - вірус мозаїки люцерни (ВМЛ, alfalfa mosaic virus, Medicago virus 2 Smith). Передається контактно-механічним шляхом, попелицями, насінням. Симптоми: спочатку з'являються дрібні округлі жовтуваті плями па листі, потім довгасті або неправильної форми плями між бічними жилками, світло-жовтий або білястий рісуіок уздовж жилок. Листя дрібне н деформовані. Влітку симптоми часто маскуються. Нерідко зустрічається латентна інфекція. Вірус має широке коло растеппй-господарів: вражає дикі і культивовані рослини багатьох родин: Метеликові, Пасльонові, Айстрові, Гарбузові і ін. [...]
Крім організмів, що мають клітинну будову, є і неклітинні форми життя-віруси і бактеріофаги. До речі, віруси були відкриті в 1892 р російським біологом Д.І. Івановим, а їх назва в перекладі означає «отрута», що в загальному-то в звичному побуті для багатьох людей відображає їх вплив на стан здоров'я. [...]
Чіткої межі між живими і неживими речовинами немає, що підтверджується існуванням вірусів. Останні мають ознаки як живого, так і неживого. Загальноприйнятого визначення для них поки не сформульовано. Зазвичай вважають, що віруси - це найменш організовані форми життя, що не володіють власним обміном речовин і здатні існувати тільки всередині клітин інших організмів. Поза клітин вони не розмножуються. Разом з тим здатність вірусів відтворюватися, хоча б і в контакті з іншими клітинами, є ознакою живого. [...]
У грунті містяться різні мікроорганізми: бактерії, актиноміцети або променисті грибки, гриби, віруси та ін. Більшість з них переробляє лісову підстилку (гумусовий шар), покращує структуру грунту, переводить органічні сполуки в засвоювані форми. З підвищенням кислотності грунту і утворенням розчинних форм токсичних металів активність мікроорганізмів, особливо в переробці лісової підстилки, знижується. [...]
На інактивує дію продуктів електролізу і хлору великий вплив мають кількість і форма залишкового хлору (вільне володіння або пов'язаний). Вивчення динаміки інактивації продуктами електролізу і хлором модельного вірусу поліомієліту, кишкової палички та фага кишкової палички показало, що при наявності залишкового хлору лише у зв'язаному стані до 30 хв контакту кишкова паличка гинула повністю, а фаги вірус - лише на 80 і 60% відповідно. При слідах вільного залишкового хлору до 20 хв контакту кишкова паличка і фаг інактивована більш ніж на 99%, а вірус - лише на 90%. При вмісті у воді вільного залишкового хлору 0,1-0,3 мг / л до 10 хв контакту спостерігалося повне знезараження її відносно кишкової палички та фага, а до 30 хв виявлялося лише незначна кількість активних вірусів. Різниця між ступенем інактивації досліджуваних мікроорганізмів була в усіх випадках статистично достовірної. При випробуваних умовах знезараження продуктами електролізу і хлором води, що містить мікроорганізми в рівних концентраціях, кишкова паличка виявлялася менш стійка, ніж фаг, а фаг менш стійкий, ніж вірус. Отже, кишкова паличка і фаг можуть служити надійними санітарними показниками ефективного знезараження води продуктами електролізу і хлором щодо ентеровірусів. В основному це відноситься до тих випадків, коли в силу несприятливих епідемічних санітарних умов концентрація ентеровірусів у воді водойм може значно збільшитися і досягти рівня вмісту кишкової палички (Е. Л. Ловцевіч, Л. А. Сергуніна, 1968). [...]
Так, після винаходу антибіотиків головним ворогом людини стали не найпростіші грибки і одноклітинні, а віруси. Є перші симптоми того, що на зміну вірусам грядуть ретровіруси - довірусние, давніші форми життя, що будують свою організацію не на основі молекули ДНК, а на основі РНК. Одним з найбільш відомих представників цієї форми життя є ретровірус СНІДу. [...]
Мікроорганізми, невидимі під мікроскопом, називаються ультрамікробів. З цієї групи ультра-мікроскопічних форм найбільш важливе значення в практичній діяльності людини мають бактеріофаги - фільтрівні віруси і невидимі форми бактерій. Спостерігати ультрамікробів вдалося тільки в електронний мікроскоп, який дає збільшення до 45 000 разів. Віруси (рис. 85) являють собою частинки, що складаються з білкових речовин і нуклеїнових кислот (ДНК або РНК). Вони не мають звичайної клітинної структурою. До неклеточной формі життя відносяться також бактеріофаги (рис. 86), що представляють собою подовжені освіти з потовщеним кінцем. [...]
Тнфекціонний процес - це комплекс реакцій в макроорганизме, що виникають у відповідь на впровадження і розмноження в ньому мікробів, вірусів та ін. Він не завжди супроводжується наявністю ознак хвороби. Наприклад, при мікробоносітельстве або безсимптомному перебігу інфекції клінічні ознаки відсутні, хоча її збудник присутній в організмі і впливає на його різні системи, викликаючи імунологічну перебудову останнього. Якщо інфекційний процес супроводжується проявом клінічних ознак, то таку форму інфекції називають інфекційною хворобою. Отже, інфекційна хвороба є так званої маніфестной формою інфекції. [...]
Ці віруси характеризуються схожістю за морфологічними ознаками, реакцій електромагнітне опромінення, репродукції та ін. Їх основні іческого складові: С, Н, N, Р, О, вуглеводи і ліпіди. Відомо, що прак-Пріес і все онковіруси термічно нестабільні і руйнуються температура від 50 до 70 ° С в залежності від виду онковірус. [...]
У цю групу входять колоїдні (мінеральні і органо) частки грунтів і грунтів, а також недіссоціірованних і нерозчинні форми гумусових речовин, які надають воді забарвлення. Останні вимиваються в природні водойми з лісових, болотистих і торф'яних грунтів, а також утворюються в самих водоймах в результаті життєдіяльності водних рослин і водоростей. До цієї групи можна віднести також віруси і інші організми, що наближаються за розмірами до колоїдних частинок. Так як серед них знаходяться хвороботворні (патогенні) організми, то видалення їх з води є досить відповідальним заходом. [...]
Друга група домішок об'єднує гідрофільні і гідрофобні мінеральні та органо-мінеральні колоїдні частинки ґрунтів і ґрунтів, недіссо-ціірованние і нерозчинні форми високомолекулярних гумусових речовин і детергенти. Кінетична стійкість гідрофобних домішок характеризується співвідношенням сил гравітаційного поля і броунівського руху; Агрегативна стійкість їх обумовлена \u200b\u200bелектростатичним станом міжфазної поверхні і утворенням при цьому дифузних шарів або створенням на поверхні частинок стабілізуючих шарів. До цієї групи належать також віруси та інші мікроорганізми, близькі за розмірами до колоїдних частинок. [...]
Метод центрифугування в градієнті щільності, розроблений Шлюб-ке, можна використовувати як для виділення, так і для отримання кількісних характеристик вірусів рослин. Як виявилося, цей метод таїть в собі багато можливостей і в даний час широко використовується в області вірусології та молекулярної біології. При проведенні досліджень методом центрифугування в градієнті щільності центрифужную пробірку частково наповнюють розчином, щільність якого зменшується в напрямку від дна до меніска. Для створення градієнта при фракціонуванні вірусів рослин найбільш часто використовують сахарозу. Перед початком центрифугування частинки вірусу можуть бути або розподілені в усьому об'ємі розчину, або нанесені на вершину градієнта. Бракке запропонував три різних прийому центрифугування в градієнті щільності. При ізопікпіческом (рівноважному) центрифугировании процес триває до тих пір, поки всі частинки в градієнті не досягнуть рівня, де щільність середовища дорівнює їх власної щільності. Таким чином, фракціонування частинок нроісходіт в цьому випадку відповідно до відмінностями в їх щільності. Розчини сахарози не володіють достатньою щільністю для изопикнических поділу багатьох вірусів. При швидкісному зональному центрифугировании вірус спочатку наносять па попередньо створений градієнт. Частинки кожного типу седіментіруют при, цьому через градієнт у вигляді зони, або смуги, зі швидкістю, яка залежить від їх розміру, форми і щільності. Центрифугування при цьому закінчують, коли частки ще продовжують седіментіровать. Рівноважний зональне центрифугування подібно зі швидкісним зональним центрифугуванням, по в цьому випадку центрифугування триває до досягнення изопикнических стану. Роль градієнта щільності при швидкісному центрифугировании полягає в тому, щоб перешкоджати конвекції я фіксувати різні види молекул в певних зонах. Теорія центрифугування в градієнті щільності складна і не зовсім зрозуміла. На практиці ж це простий і витончений метод, який широко застосовується при роботі з вірусами рослин. [...]
Основною особливістю локалізованого в матриці ЩГК ЦЕЧ (як і онковирусов в клітці) є наявність кордону розділу двох середовищ з різною провідністю. На рис. 2.11 наведені дані електронної мікроскопії, що показують аденовірус, вірус Епштейна-Барр (ЕБВ) і ЦЕЧ в ЩГК. З рис. 2.11 видно, що все освіти одного масштабу, мають форму близьку до сферичної, що складається з ядра і оболонки, в хімічний склад кожної оболонки містяться електрично активні іони, чітко позначені виражені кордону у вірусів і ЦЕЧ зі своїми матрицями. [...]
Виявляється на листках ранньою весною у вигляді жовтої крапчатости. До середини літа ця ознака зникає, але уражені листя іноді стають зморшкуватими. Плоди формуються дрібні, часто неправильної форми і з горбиками уздовж шва. Дозрівання їх затримується. Збудник хвороби - Peach mosaic virus передається при щепленні і окуліровці. Вважають, що переносником вірусу є сливова попелиці. [...]
Незважаючи на багато відкриттів, в картині біогенезу залишається ще чимало білих плям. Можна вважати безперечними лише основні віхи. Так, не викликає тепер сумнівів, що виникненнябіосфери було винятковим, одиничним подією. Мізерно малий вірус і гігантське чудовисько моря, одноклітинна водорість і деревоподібна папороть, який зник мільйони років тому, - всі вони тільки гілки і листя на одному філогенетичному дереві. Форми життя завжди і всюди виявляють, так би мовити, «кровна спорідненість», і все її діти генетично пов'язані між собою. З того дня, коли на Землі з'явилося перше істота, життя відбувається тільки від життя. [...]
Клітка - основна структурно-функціональна одиниця всіх живих організмів, елементарна жива система. Вона може існувати як окремий організм (бактерії, найпростіші, деякі водорості і гриби), так і в складі тканин багатоклітинних організмів. Лише віруси являють собою неклітинні форми життя. [...]
Згідно із запропонованою схемою на першій стадії процесу відбувається утворення фермент-субстратного комплексу EI ендонуклеази рестрикції EcoR I і двухцепочечной плазмідної ДНК. Ключовим моментом схеми є утворення комплексу E-II ендонуклеази рестрикції EcoR I з кільцевої формою ДНК, що містить однонітевую розрив, отриманої в результаті гідролізу фосфодіефірних зв'язку в одному з ланцюжків ДНК У подальшому, залежно від умов (природи субстрату, температури і т. Д. ) може відбуватися або розщеплення другого ланцюга ДНК у складі того ж комплексу Е - II з утворенням комплексу Е-III-ферменту з лінійної формою ДНК або дисоціація комплексу Е-П з утворенням вільного ферменту і кільцевої ДНК, що містить однонітевую розрив, що і призводить до накопиченню форми II в розчині. Ця схема дозволила пояснити відмінності в механізмах гідролізу ДНК вірусу SV 40 з одного боку і ДНК ColE I і бактеріофага G4 з іншого. У разі ДНК вірусу SV 40 відбувається дисоціація фермент-субстратного комплексу Е -11, яка веде до накопичення кільцевої форми ДНК в розчині. Було висловлено припущення, що відмінності в механізмі гідролізу цих ДНК молекул (вірусу SV 40; ДНК ColE I і бактеріофага G4) є результатом взаємодії рестріктази EcoR I з різними нуклеотидними послідовностями, фланкуючими ділянку впізнавання рестриктаз EcoR I. Однак, таке припущення не дозволяє пояснити відмінності в механізмі гідролізу кільцевої ДНК ColE I в залежності від температури (див. вище). [...]
Захворювання відоме в багатьох країнах світу. В СРСР виявлено на Україні, в Молдавії, Естонії та Грузії і є об'єктом внутрішнього карантину. Уражаються слива, алича, мірабель, абрикос і персик. Збудник хвороби - Plum pox (\u003d Prunus virus 7 Smith). Форма вірусу - ниткоподібна, розмір 760X20 ім. [...]
Подальший розвиток дослідження з вивчення механізму гідролізу плазмідної ДНК ендонуклеаза рестрикції отримали в роботах Халфорд з співр. . Таким чином механізм реакції є аналогічним такому, запропонованого для гідролізу ДНК вірусу ЕУ 40 рестриктазой есоі I. [...]
Крім вищевказаних «общеорганізменний» функцій наявність гомеостазу організму існує ще одна дуже важлива особливість: жива речовина як би створює ще одну середовище проживання, а саме можливість заселення організму іншими живими істотами для постійного або тимчасового проживання. Це створена життям нова біотична середовище проживання. До істотам, які заселяють цю середу, багато фахівців відносять віруси. Так, І.А. Шилов (2000) вважає, що виняткова простота їх влаштування є вторинним явищем, навіть швидше це знову виникла форма живих істот, повністю осврівшая внутрішньоклітинну середу в організмах інших рівнів. Другим підтвердженням цієї тези є те, що віруси мають високий ступінь складності і різноманітності генетичної системи. Спрощення будови, що стало можливим завдяки обов'язковим безумовним зв'язків вірусів з господарем-організмом, що забезпечує стабільні умови життя, торкнулося навіть фундаментальні властивості, притаманні переважній більшості форм життя: віруси не мають подразливістю і позбавлені власного апарату синтезу білка. Віруси не здатні до самостійного існування, і їх зв'язок з клітиною-це не тільки просторова, а й жорстка функціональний зв'язок, з якої клітина і вірус представляють певну єдність. [...]
Короткочасна лужна обробка ВЖМТ при 30 ° С і високої іонної силі in situ викликає розриви, приводячи до утворення фрагментів РНК, досить однорідних за величиною, у яких s2 [...]
Кількість людей, уражених малярією, гепатитом, ВІЛ та багатьма іншими хворобами, обчислюється величезними цифрами. Багато медиків вважають, що слід говорити не про «перемогу», а лише про тимчасове успіху в боротьбі з цими хворобами. Історія боротьби з інфекційними хворобами дуже коротка, а непередбачуваність змін у навколишньому середовищі (особливо, в міській) може звести нанівець ці успіхи. З цієї причини «повернення» інфекційних агентів фіксується серед вірусів. Багато віруси «відриваються» від природної основи і переходять в нову стадію, здатну жити в середовищі проживання людини, - стають збудниками грипу, вірусної форми раку та інших хвороб. Можливо, такою формою є ВІЛ. [...]
Про зміну средневесовой молекулярної маси і радіусу обертання судили на підставі даних светорассеяния. Для РНК ВЖМТ, застосовуючи обидва методи деградації, вони знайшли, що радіус обертання збільшувався перед початком інтенсивної деградації молекули, в той час як радіус обертання і средневесовой молекулярна маса РНК ВТМ зменшувалися з самого початку цього процесу. Страціеллі і ін. Пояснили ці дані, припустивши, що РНК ВЖМТ існує в формі замкнутої петлі. Однак ці результати можна інтерпретувати і но-іншому. Наприклад, Хазелькорн показав, що РНК ВТМ і РНК ВЖМТ седіментіровалі спільно в умовах pH і іонної сили, подібних до тих, які були використані Страціеллі і ін.. На противагу цьому кільцева і ліхгейная форма ДНК фага срХ174 легко помітні за своїми седіментаціондьш властивостями 1,515]. Кейпер на підставі даних про седиментації при різних умовах висловив припущення, що ізольована РНК вірусу огіркової мозаїки (штам У) може існувати в двох формах: незамкненою ланцюга і кільцевої структури. Однак ці дані, так само як і в описаному вище випадку, можна пояснити по-різному. [...]
Бактеріальні ДНК - це високополімерні сполуки, що складаються з великого числа нуклеотидів - полінуклеотіди з молекулярною вагою близько 4 млн. Молекула ДНК являє собою ланцюг нуклеотидів, де розташування їх має певну послідовність. У послідовності розташування азотистих основ закодована генетична інформація кожного виду. Порушення цієї послідовності можливо при природних мутаціях або ж під впливом мутагенних чинників. При цьому мікроорганізм набуває або втрачає будь-яке властивість. У нього спадково змінюються ознаки, т. Е. З'являється нова форма мікроорганізму. У всіх мікроорганізмів - прокаріотів та еукаріотів - носіями генетичної інформації є нуклеїнові кислоти - ДНК і РНК. Лише деякі віруси являють собою виняток: у них ДНК відсутня, а спадкова інформація записана або відображена тільки в РНК.