Vírusové choroby vznikali v staroveku, ale virológia ako veda sa začala rozvíjať na konci 19. storočia.
V roku 1892 ruský vedec-botanik DI Ivanovskij, ktorý študoval mozaikové ochorenie tabakových listov, zistil, že túto chorobu spôsobujú najmenšie mikroorganizmy, ktoré prechádzajú cez bakteriálne filtre s jemnými pórmi. Tieto mikroorganizmy sa nazývajú filtrovateľné vírusy (z latinského vírusu - jed). Neskôr sa ukázalo, že bakteriálnymi filtrami prechádzajú ďalšie mikroorganizmy, preto sa filtrované vírusy začali nazývať jednoducho vírusy.
Tvar viriónu a prítomnosť alebo neprítomnosť obalu vo všeobecnosti hovoria len málo o tom, ktoré choroby môžu vírusy spôsobiť alebo ktoré druhy môžu infikovať, ale stále sú užitočným spôsobom na zahájenie klasifikácie vírusov. Obrázok 5 Vírusy môžu mať zložitý tvar alebo sú pomerne jednoduché.
Ktoré z nasledujúcich tvrdení o štruktúre vírusu je správne? Jadro vírusu obsahuje genóm alebo všeobecný genetický obsah vírusu. Vírusové genómy sú zvyčajne malé v porovnaní s baktériami alebo eukaryotmi, ktoré obsahujú iba gény kódujúce proteíny, ktoré vírus nemôže získať z hostiteľskej bunky. Tento genetický materiál môže byť jednovláknový alebo dvojvláknový. Môže to byť aj lineárne alebo kruhové. Zatiaľ čo väčšina vírusov obsahuje jeden segment nukleovej kyseliny, iné majú genómy, ktoré sú zložené z viacerých segmentov.
Veľký príspevok k štúdiu vírusov priniesli sovietski virológovia: M.A. Morozov, N. F. Gamaleya, L. A. Zilber, M. P. Chumakov, A. A. Smorodintsev, V. M. Zhdanov a ďalší.
Vírusy sú nebunková forma existencia živej hmoty. Sú veľmi malé. Podľa obrazného vyjadrenia VM Zhdanova „možno ich veľkosť vo vzťahu k veľkosti priemerných baktérií porovnať s veľkosťou myši vo vzťahu k slonovi“. Vírusy bolo možné vidieť až po vynájdení elektrónového mikroskopu.
Vírusy možno považovať za povinné intracelulárne parazity... Vírus sa musí naviazať na živú bunku, stráviť ju, produkovať bielkoviny a kopírovať genóm a nájsť spôsob, ako z bunky uniknúť, aby mohol infikovať ďalšie bunky a nakoniec aj ďalších ľudí. Vírusy môžu infikovať iba určité druhy hostiteľov a iba určité bunky v danom hostiteľovi. Molekulárny základ tejto špecificity spočíva v tom, že na povrchu hostiteľskej bunky sa musí nachádzať špecifická povrchová molekula známa ako vírusový receptor, aby sa k nemu mohol vírus pripojiť.
V súčasnosti sa na štúdium vírusov používa veľa metód: chemické, fyzikálne, molekulárne biologické, imunobiologické a genetické.
Všetky vírusy sa ďalej delia na infikovanie ľudí, zvierat, hmyzu, baktérií a rastlín.
Vírusy majú širokú škálu foriem a biologických vlastností, ale všetky majú spoločné štrukturálne vlastnosti. Zrelé vírusové častice sa nazývajú virióny.
Okrem toho sú metabolické rozdiely pozorované u rôznych typov buniek založené na diferenciálnej génovej expresii pravdepodobným faktorom, pri ktorom môžu bunky použiť vírus na replikáciu. Bunka musí tvoriť látky potrebné pre vírus, ako sú enzýmy, v ktorých samotný vírusový genóm neobsahuje gény, alebo sa vírus nedokáže replikovať pomocou tejto bunky.
Vírus musí na replikáciu „prevziať“ bunku. Cyklus vírusovej replikácie môže viesť k dramatickým biochemickým a štrukturálnym zmenám v hostiteľskej bunke, ktoré môžu spôsobiť poškodenie buniek. Tieto zmeny, nazývané cytopatické účinky, môžu zmeniť funkciu bunky alebo ju dokonca zničiť. Niektoré infikované bunky, ako napríklad bunky infikované vírusom prechladnutia, zomierajú lýzou alebo apoptózou a súčasne uvoľňujú všetky virióny potomstva. Príznaky vírusové ochorenia sú výsledkom imunitnej odpovede na vírus, ktorý sa snaží vírus riadiť a eliminovať z tela, ako aj pred poškodením buniek vírusom.
Na rozdiel od iných mikroorganizmov obsahujúcich DNA aj RNA obsahuje virión iba jednu z nukleových kyselín - buď DNA alebo RNA.
Nukleová kyselina vírusov môže byť jednovláknová a dvojvláknová. Takmer všetky vírusy obsahujúce RNA majú vo svojom genóme jednovláknovú RNA a tie, ktoré obsahujú DNA, majú dvojvláknovú DNA. V súlade s dvoma typmi genetických látok sa vírusy delia na RNA a DNA. Medzi rodiny obsahujúce DNA patrí 5 rodín, rodiny obsahujúce RNA - 10 rodín.
Počas začínajúceho procesu nie je bunka lýzovaná alebo usmrcovaná okamžite. Najproduktívnejšie vírusové infekcie sa riadia podobnými krokmi v cykle vírusovej replikácie: hniezdenie, penetrácia, nekrytie, replikácia, zhromaždenie a uvoľnenie. Vírus sa viaže na špecifické receptorové miesto na membráne hostiteľskej bunky prostredníctvom väzbových proteínov v kapside alebo proteínov zovretých v jeho obale. Pripojenie je špecifické a vírus sa obvykle pripojí iba k bunkám jedného alebo viacerých druhov a iba k určitým typom buniek v rámci týchto druhov so zodpovedajúcimi receptormi.
* (Tu sú údaje týkajúce sa iba niektorých vírusov patogénnych pre ľudí.)
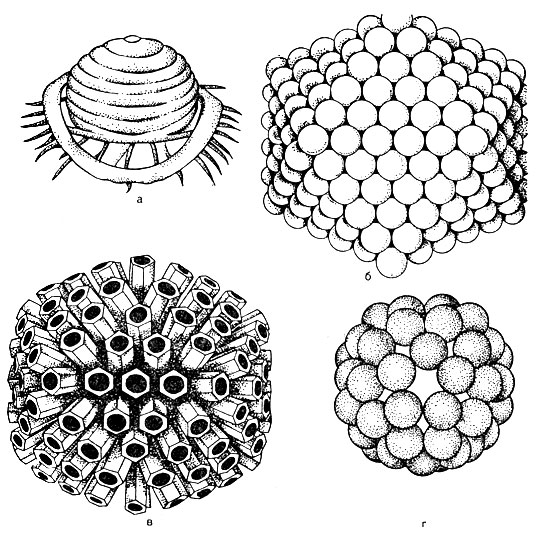
Štruktúra viriónu... V strede viriónu je nukleová kyselina, ktorá je obklopená kapsidom (z gréckeho kanca - box). Kapsida je tvorená proteínovými podjednotkami nazývanými kapsoméry. Zrelý vírus je nukleokapsid v chemickej štruktúre. Počet kapsomérov a spôsob ich balenia (obr. 52) sú pre každý typ vírusu striktne konštantné. Napríklad vírus poliomyelitídy obsahuje 32 kapsomérov, zatiaľ čo adenovírus obsahuje 252 kapsomérov. Capsomeres môžu byť stohované vo forme mnohostena s jednotnými symetrickými okrajmi - kvádrový tvar (napríklad adenovírus). Pre chrípkové vírusy je charakteristický špirálovitý (sférický) tvar. Môže existovať typ symetrie, v ktorej má nukleová kyselina formu pružiny, okolo ktorej sú položené kapsoméry, a v takom prípade má vírus formu tyčinky - vírus, ktorý spôsobuje ochorenie tabakových listov.
Na rozdiel od živočíšnych vírusov sa nukleová kyselina bakteriofága zavádza do hostiteľskej bunky nahá, takže kapsida zostáva mimo bunky. Rastlinné a živočíšne vírusy môžu vstúpiť do buniek endocytózou, v ktorej bunková membrána obklopuje a pohlcuje celý vírus. Niektoré obalené vírusy vstupujú do bunky, keď sa vírusová obálka spojí priamo s bunkovou membránou. Vírusová kapsida sa potom rozkladá vo vnútri bunky a uvoľňuje sa vírusová nukleová kyselina, ktorá je potom dostupná na replikáciu a transkripciu.
Mechanizmus replikácie závisí od vírusového genómu. Z tohto modelu samozrejme existujú výnimky. Ak hostiteľská bunka neposkytuje enzýmy potrebné na vírusovú replikáciu, vírusové gény poskytujú informácie na priamu syntézu chýbajúcich proteínov.
Fág má zložitý typ symetrie: hlava je kvádrová a proces je v tvare tyčinky (v tvare spermií) (pozri obr. 21, 22).
V závislosti na spôsobe balenia sa teda vírusy ďalej delia na kockovitú, sférickú, tyčkovitú a spermatozoidnú formu.
Posledným stupňom vírusovej replikácie je uvoľnenie nových viriónov do hostiteľského organizmu, kde sú schopné infikovať susedné bunky a opakovať replikačný cyklus. Niektoré vírusy sa uvoľňujú, keď hostiteľská bunka zomrie, zatiaľ čo iné vírusy môžu opustiť infikované bunky únikom cez membránu bez priameho zabitia bunky.
Obrázok 6 Keď sú infikované vírusom chrípky, glykoproteíny sa naviažu na hostiteľskú epiteliálnu bunku. Vďaka tomu sa vírus vstrebáva. Obrázok 7 Vírusy spôsobujú u ľudí rôzne choroby, od ľahkých chorôb až po vážne ochorenia.
Niektoré vírusy, ktoré sú zložitejšie, majú škrupinu nazývanú peplos. Vzniká, keď vírus opustí hostiteľskú bunku. V tomto prípade je vírusová kapsida obalená vnútorným povrchom cytoplazmatickej membrány hostiteľskej bunky a je vytvorená jedna alebo viac vrstiev superkapsidového obalu. Iba niektoré vírusy majú taký obal, napríklad vírusy besnoty, herpesu, encefalitídy. Táto škrupina obsahuje fosfolipidy, ktoré sa odbúravajú éterom. Pôsobením na éter je teda možné rozlíšiť vírus s peplosom od vírusu s „holým kapsidom“.
Vakcína sa môže pripraviť pomocou oslabených živých vírusov, usmrtených vírusov alebo molekulárnych podjednotiek vírusu. Živé vírusy vo všeobecnosti vedú k zvýšeniu imunity, ale sú schopné spôsobiť chorobu s nízkou frekvenciou. Zabitá vírusová vakcína a vírusové podjednotky nie sú schopné spôsobiť ochorenie, ale spravidla vedú k menej efektívnej alebo dlhodobej imunite.
Oslabené živé vírusové vakcíny sú navrhnuté v laboratóriách tak, aby spôsobovali u príjemcov málo príznakov a zároveň im poskytovali imunitu proti budúcim infekciám. Poliomyelitída bola jednou z chorôb, ktoré znamenali míľnik v používaní vakcín. Úspech vakcíny proti obrne pripravil pôdu pre rutinnú distribúciu detských vakcín proti osýpkam, príušniciam, ružienke, kiahňam a iným chorobám. Živé vakcíny sa zvyčajne vyrábajú oslabením vírusu divokého typu jeho pestovaním v laboratóriu v tkanivách alebo pri teplotách odlišných od tých, na ktoré je vírus zvyknutý v hostiteľovi.
U niektorých vírusov vyčnievajú z vonkajšej lipidovej vrstvy obalu kapsoméry vo forme tŕňov (tieto tŕne sú tupé). Takéto vírusy sa nazývajú peploméry (napr. Vírus chrípky, pozri obrázok 52).
Nukleová kyselina vírusu je nosičom dedičných vlastností a kapsida a vonkajší obal majú ochranné funkcie, akoby chránili nukleovú kyselinu. Okrem toho uľahčujú vstup vírusu do bunky.
Napríklad môže byť vírus pestovaný v bunkách in vitro, embryách vtákov alebo živých zvierat. Prispôsobenie sa týmto novým bunkám alebo teplote spôsobuje mutácie v genómoch vírusu, čo im umožňuje lepšie rásť v laboratóriu a zároveň narúšať ich schopnosť spôsobiť ochorenie pri opätovnom zavedení do stavu nájdeného v hostiteľovi. Tieto oslabené vírusy teda stále spôsobujú infekciu, ale nerastú veľmi dobre, čo umožňuje vývoj imunitných reakcií v priebehu času, aby sa zabránilo vážnemu ochoreniu.
Nebezpečenstvo použitia živých vakcín, ktoré sú všeobecne účinnejšie ako zabité vakcíny, je nízke, ale významné riziko, že sa tieto vírusy reverznými mutáciami vrátia do svojej pôvodcu spôsobujúcej choroby. K reverzným mutáciám dochádza, keď vakcína mutuje v hostiteľovi tak, že sa číta hostiteľovi a môže znova spôsobiť ochorenie, ktoré sa potom môže rozšíriť na ďalších ľudí v epidémii.
Veľkosti vírusov... Vírusy sa merajú v nanometroch. Ich veľkosť kolíše v širokom rozmedzí od 15-20 do 350-400 nm.
Metódy merania vírusov: 1) filtrácia cez bakteriálne filtre so známou veľkosťou pórov; 2) ultracentrifugácia - veľké vírusy sa zrážajú rýchlejšie; 3) fotografovanie vírusov v elektrónovom mikroskope.
V prípade chrípky mutácia génov pre povrchové molekuly pomáha vírusu uniknúť z ochrannej imunity, ktorú mohol dostať v predchádzajúcej chrípkovej sezóne, a preto je nevyhnutné, aby boli ľudia očkovaní každý rok. Ostatné vírusy, napríklad vírusy, ktoré spôsobujú detské choroby, osýpky, príušnice a ružienka, mutujú tak málo, že sa rok čo rok používa rovnaká vakcína.
Vakcíny a antivírusové lieky na liečbu
V niektorých prípadoch sa môžu na liečbu aktívnej vírusovej infekcie použiť vakcíny. V prípade besnoty, smrteľného neurologického ochorenia prenášaného v slinách vírusov infikovaných besnotou, môže byť progresia ochorenia od pohryznutia zvieraťom do okamihu, keď vstúpi do centrálneho nervového systému, dva týždne alebo dlhšie. To je dostatočný čas na očkovanie osoby podozrivej z uhryznutia besným zvieraťom a zvýšená imunitná odpoveď z očkovania je dostatočná na to, aby sa zabránilo vstupu vírusu do nervového tkaniva.
Chemické zloženie vírusov... Množstvo a obsah vírusov DNA a RNA nie sú rovnaké. V DNA sa molekulová hmotnosť pohybuje od 1,106 do 1,6,108 a v RNA od 2,106 do 9,0,106.
Bielkoviny vo viriónoch sa nachádzajú v zanedbateľnom množstve, pozostávajú z 16 - 20 aminokyselín. Okrem kapsidových proteínov sú s nukleovou kyselinou spojené aj vnútorné proteíny. Proteíny určujú antigénne vlastnosti vírusov a tiež vďaka hustému zhluku polypeptidových reťazcov chránia vírus pred pôsobením enzýmov hostiteľskej bunky.
Týmto spôsobom sa zabráni smrteľným neurologickým následkom choroby a jedinec sa musí zotaviť iba z infikovaného uhryznutia. Tento prístup sa tiež používa na liečbu eboly, jedného z najrýchlejších a najsmrteľnejších vírusov, ktoré infikujú ľudí, hoci zvyčajne infikuje obmedzenú populáciu. Ebola je tiež hlavnou príčinou smrti goríl. Dúfame, že používaním novo vyvinutých vakcín, ktoré zvyšujú imunitnú odpoveď, bude imunitný systém postihnutých ľudí lepšie kontrolovať vírus, čo potenciálne zníži úmrtnosť.
Lipidy a sacharidy sa nachádzajú vo vonkajšom obale komplexných viriónov. Membrána hostiteľskej bunky je zdrojom lipidov a sacharidov. Polysacharidy, ktoré tvoria niektoré vírusy, určujú ich schopnosť spôsobiť aglutináciu erytrocytov.
Vírusové enzýmy... Vírusy nemajú svoj vlastný metabolizmus, takže nepotrebujú metabolické enzýmy. Zistilo sa však, že niektoré vírusy majú enzýmy, ktoré uľahčujú ich penetráciu do hostiteľskej bunky. Napríklad vo vírusu chrípky A bola nájdená neuraminidáza, ktorá štiepi kyselinu neuraminovú obsiahnutú v membránach živočíšnych buniek (erytrocyty atď.). Fágy majú lyzozým, ktorý ničí bunkovú membránu, fosfatázu atď.
Ďalším spôsobom liečby vírusových infekcií je použitie antivírusových liekov. Tieto lieky majú často obmedzenú schopnosť liečiť vírusové ochorenie, ale používajú sa na potlačenie a zmiernenie symptómov širokého spektra vírusových ochorení. U väčšiny vírusov tieto lieky inhibujú vírus blokovaním účinku jedného alebo viacerých jeho proteínov. Je dôležité, aby cieľové proteíny boli kódované vírusovými génmi a aby tieto molekuly neboli prítomné v zdravých hostiteľských bunkách.
Takto je inhibovaný vírusový rast bez poškodenia hostiteľa. Je ich veľa antivírusové liekydostupné na liečbu infekcií, niektorých špecifických pre konkrétny vírus a iných, ktoré môžu ovplyvniť viac vírusov. Antivírusové lieky sú určené na liečbu genitálneho herpesu a chrípky. V prípade genitálneho herpesu môžu lieky ako acyklovir znížiť počet a trvanie epizód aktívneho vírusového ochorenia, pri ktorom sa u pacientov vyvinú vírusové lézie v kožných bunkách.
Detekcia vírusových antigénov... Vírusové antigény v infikovaných hostiteľských bunkách je možné detegovať pomocou imunofluorescenčných techník. Prípravky obsahujúce bunky infikovaný vírusomsú ošetrené špecifickými imunitnými luminiscenčnými sérami. Pri pohľade pod fluorescenčným mikroskopom sa na miestach, kde sa hromadia vírusové častice, pozoruje charakteristická žiara. Typ vírusu je určený korešpondenciou špecifického luminiscenčného séra, ktoré spôsobilo luminiscenciu.
Pretože vírus zostáva po celý život skrytý v nervovom tkanive tela, tento liek nie je liečbou, ale môže zmierniť príznaky choroby. Pri chrípke môžu lieky ako Tamiflu skrátiť trvanie príznakov chrípky o jeden alebo dva dni, ale liek úplne nezabráni prejavom. Ostatné antivírusové liekyako ribavirín sa používajú na liečbu rôznych vírusových infekcií.
Ak sa ktorékoľvek z týchto liekov použije samostatne, vysoká rýchlosť mutácie vírusu umožňuje vírusu rýchlo vyvinúť rezistenciu na liečivo. Vírusy sú entity bez buniek, ktoré sa zvyčajne dajú pozorovať iba pomocou elektrónového mikroskopu. Vírusy sú rozmanité a infikujú archy, baktérie, huby, rastliny a zvieratá. Vírusy pozostávajú z jadra nukleovej kyseliny obklopeného proteínovou kapsidou s alebo bez vonkajšieho lipidového obalu.
Zavedenie vírusu do bunky, jeho interakcia s hostiteľskou bunkou a reprodukcia (reprodukcia) sú zložené zo série po sebe nasledujúcich etáp.
Fáza 1. Začína procesom adsorpcie na úkor viriónov a bunkových receptorov. V komplexných viriónoch sú receptory umiestnené na povrchu obalu vo forme výrastkov styloidu (vírus chrípky), v jednoduchých viriónoch na povrchu kapsidy.
Fáza 2. Prienik vírusu do hostiteľskej bunky prebieha u rôznych vírusov odlišne. Napríklad niektoré fágy svojím procesom prerazia membránu a injikujú nukleovú kyselinu do hostiteľskej bunky (pozri kapitolu 8). Ďalšie vírusy vstupujú do bunky vtiahnutím vírusovej častice pomocou vakuoly, to znamená, že v mieste zavedenia do bunkovej membrány sa vytvorí priehlbina, potom sa jej okraje uzavrú a vírus vstúpi do bunky. Toto stiahnutie sa nazýva viropexis.
Fáza 3. „Odstránenie vírusu“ (dezintegrácia). Pre svoju reprodukciu je vírusová nukleová kyselina zbavená svojich ochranných proteínových obalov (obal a kapsida). Proces vyzliekania sa môže začať počas adsorpcie alebo k nemu môže dôjsť, keď je vírus už v bunke.
Fáza 4. V tomto štádiu dochádza k replikácii (reprodukcii) nukleových kyselín a syntéze vírusových proteínov. Toto štádium nastáva za účasti DNA alebo RNA hostiteľskej bunky.
Fáza 5. Zhromaždenie viriónu. Tento proces je uľahčený samostatným zhromažďovaním proteínových častíc okolo vírusovej nukleovej kyseliny. Syntéza proteínov môže začať okamžite po syntéze vírusových nukleových kyselín alebo po intervale niekoľkých minút alebo niekoľkých hodín. U niektorých vírusov sa samovznikanie vyskytuje v cytoplazme. Iné majú hostiteľské bunky v jadre. K tvorbe vonkajšieho obalu (peplos) dochádza vždy v cytoplazme.
Fáza 6. Uvoľnenie viriónu z hostiteľskej bunky nastáva perkoláciou vírusu cez bunkovú membránu alebo cez otvor vytvorený v hostiteľskej bunke (v tomto prípade hostiteľská bunka zomrie).
Typy interakcie vírus-bunka... Prvý typ, produktívna infekcia, je charakterizovaný tvorbou nových viriónov v hostiteľskej bunke.
Druhým typom - abortívna infekcia - je prerušenie replikácie nukleových kyselín.
Tretí typ je charakterizovaný inkorporáciou vírusovej nukleovej kyseliny do DNA hostiteľskej bunky; existuje forma koexistencie vírusu a hostiteľskej bunky (virogénia). V tomto prípade je zabezpečená synchronicita replikácie vírusovej a bunkovej DNA. Vo fágoch sa to nazýva lyzogenéza.
Mikroskopické vyšetrenie... Pri jednotlivých vírusových infekciách sa v cytoplazme alebo jadrách hostiteľských buniek pozorujú špecifické intracelulárne telieska - inklúzie, ktoré majú diagnostickú hodnotu (telieska Babesh-Negri v prípade besnoty, telieska Guarnieri v prípade kiahní atď.). Veľkosti vírusových častíc a inklúzií telies možno umelo zväčšiť špeciálnymi metódami spracovania prípravkov moridlom a impregnáciou (napríklad metódou postriebrenia podľa Morozova) a pozorovať imerznou mikroskopiou. Menšie virióny, ktoré ležia mimo dohľadu optického mikroskopu, sú detekované iba elektrónovou mikroskopiou. Pokiaľ ide o intracelulárne inklúzie, existujú rôzne uhly pohľadu. Niektorí autori sa domnievajú, že predstavujú súbor vírusov. Iní sa domnievajú, že vznikajú v dôsledku reakcie bunky na zavedenie vírusov.
Genetika vírusu... Modifikácia (nededičné zmeny) vírusov je spôsobená vlastnosťami hostiteľskej bunky, v ktorej sa vírus množí. Modifikované vírusy získavajú schopnosť infikovať bunky podobné tým, v ktorých boli modifikované. Rôzne vírusy sa prejavujú rôznymi spôsobmi. Napríklad vo fágoch sa mení tvar „negatívnych škvŕn“ (kolónie fágov).
Mutácia - vo vírusoch sa vyskytuje pod vplyvom rovnakých mutagénov, ktoré spôsobujú mutáciu baktérií (fyzikálne a chemické faktory). Mutácia nastáva počas replikácie nukleových kyselín. Mutácie ovplyvňujú rôzne vlastnosti vírusov, napríklad citlivosť na teplotu atď.
Genetická rekombinácia vo vírusoch môže byť výsledkom simultánnej infekcie hostiteľskej bunky dvoma vírusmi, zatiaľ čo jednotlivé gény je možné vymieňať medzi dvoma vírusmi a vytvárajú sa rekombinanty obsahujúce gény dvoch rodičov.
Genetická reaktivácia génov niekedy nastáva, keď sa inaktivovaný vírus skríži s úplným vírusom, čo vedie k záchrane inaktivovaného vírusu.
Spontánna a riadená genetika vírusov má veľký význam pre vývoj infekčného procesu.
Odolný voči faktorom prostredia... Väčšina vírusov je inaktivovaná pôsobením vysoké teploty... Existujú však výnimky, napríklad vírus hepatitídy je tepelne odolný.
TO nízke teploty vírusy nie sú citlivé, ultrafialové lúče slnka majú na vírusy inaktivačný účinok. Rozptýlené slnečné svetlo na ne pôsobí menej aktívne. Vírusy sú rezistentné voči glycerolu, čo umožňuje ich dlhodobé udržanie v glyceríne. Sú odolné voči antibiotikám (počas kultivácie vírusov je testovaný materiál ošetrený antibiotikami na potlačenie bakteriálnej flóry).
Kyseliny, zásady, dezinfekčné prostriedky deaktivujú vírusy. Niektoré vírusy inaktivované formalínom si však zachovávajú svoje imunogénne vlastnosti, čo umožňuje používať formalín na výrobu vakcín (vakcína proti besnote).
Náchylnosť zvierat... Spektrum vnímavých zvierat na niektoré vírusy je veľmi široké, napríklad veľa zvierat je citlivých na vírusy besnoty. Niektoré vírusy postihujú iba jeden druh zvieraťa, napríklad vírus psieho moru postihuje iba psy. Existujú vírusy, na ktoré zvieratá nie sú náchylné - napríklad vírus osýpok atď.
Organotropia vírusov... Vírusy majú schopnosť infikovať určité orgány, tkanivá a systémy. Napríklad vírus besnoty napáda nervový systém. Vírus kiahní je dermatropický atď.
Uvoľňovanie vírusov do životného prostredia... Z chorého tela sa môžu vírusy vylučovať stolicou, napríklad vírusom poliomyelitídy a inými enterovírusmi. Vírus besnoty sa vylučuje slinami, vírus chrípky sa vylučuje sliznicou nosohltanu atď.
Hlavné cesty prenosu vírusov... Kvapky vo vzduchu (chrípka, ovčie kiahne), potraviny (poliomyelitída, hepatitída A), kontaktné látky a domácnosť (besnota), prenosné (encefalitída).
Antivírusová imunita... Ľudské telo má vrodenú odolnosť voči určitým vírusom. Napríklad ľudia nie sú náchylní na vírus psieho moru. Zvieratá nie sú náchylné na vírus osýpok. V týchto prípadoch je antivírusová imunita založená na neprítomnosti buniek schopných podporovať reprodukciu vírusov.
Antivírusová imunita je daná bunkovými aj humorálnymi obrannými faktormi, nešpecifickými a špecifickými. Nešpecifické faktory. Silným inhibítorom reprodukcie vírusov je bielkovinová látka - interferón. AT zdravé telo je obsiahnutý v zanedbateľnom množstve a vírusy prispievajú k produkcii interferónu a jeho množstvo sa výrazne zvyšuje. Je nešpecifický, pretože blokuje reprodukciu rôznych vírusov. Má však tkanivovú špecifickosť, to znamená, že bunky rôznych tkanív tvoria iný interferón. Predpokladá sa, že jeho mechanizmus účinku spočíva v skutočnosti, že interferuje so syntézou proteínov v hostiteľskej bunke, a tým zastavuje reprodukciu vírusu.
Medzi špecifické faktory antivírusovej imunity patria protilátky neutralizujúce vírusy, hemaglutinačné a precipitačné protilátky.
Metódy kultivácie vírusov... Vírusy sa množia iba v životaschopných bunkách. Kultivujú sa: v kuracích embryách (obr. 53), v tkanivových kultúrach ľudí a rôznych zvierat, v tele citlivých zvierat, citlivých článkonožcoch.
V prvom období vývoja virológie bola hlavnou metódou štúdia vírusov umelá infekcia zvierat, ale táto metóda je komplikovaná a okrem toho sa ukázalo, že zvieratá sú imunné voči mnohým vírusom.
Pri vývoji virológie malo veľký význam zavedenie metód kultivácie vírusov v kuracích embryách a v kultúre buniek ľudského a zvieracieho tkaniva.
Infekcia kuracieho embrya... Na reprodukciu vírusov sa používajú kuracie embryá vo veku 7 - 12 dní inkubované v termostate pri 37 ° C. Podmienkou pre správny vývoj embrya je dodržiavanie určitej vlhkosti vzduchu, ktorú je možné vytvoriť umiestnením nádoby s vodou do termostatu.
Vhodnosť kuracieho embrya na infekciu je daná prítomnosťou pohybov embrya a rozvinutou sieťou krvných ciev na chorion-alantoidnej membráne pri skenovaní pomocou ovoskopu.
Kultivácia vírusov v kuracích embryách sa uskutočňuje na rôznych miestach infikovaného embrya (pozri obr. 53):
1) na chorion-alantoidnej membráne,
2) do alantoidnej dutiny;
3) do plodovej vody;
4) do žĺtkového vaku.
Kuracie embryá sú infikované v škatuli pomocou sterilných nástrojov. Pred infekciou sa kuracie embryá dvakrát otrú vatovým tampónom navlhčeným v alkohole.
Infekcia na chorion-alantoidnej membráne. Po dezinfekcii sa vajcia z tupého konca opatrne odrežú z časti škrupiny, membrána škrupiny sa odstráni a nájde sa chorion-alantoidná membrána. Na chorion-alantoidnú membránu sa pomocou injekčnej striekačky alebo Pasteurovej pipety nanesie infekčný materiál v množstve 0,1-0,2 ml. Po infikovaní je otvor uzavretý čiapočkou a medzera medzi ním a kuracím embryom je vyplnená parafínom.
Na druhú stranu vajíčka napíšte ceruzkou názov infekčného materiálu a dátum infekcie.
Infekcia do plodovej vody. Vajíčko je ovoskoptované a na bočnej strane je vybrané miesto, kde chorion-alantois nemá veľké krvné cievy. Táto oblasť je označená ceruzkou. Vajcia sú umiestnené na stojane vo vodorovnej polohe, dezinfikované a otvor v škrupine je prepichnutý špeciálnym sterilným oštepom do hĺbky 213 mm, cez ktorý sa v rovnakej vzdialenosti priamo do plodovej vody vpichne ihla s infekčným materiálom. Aby sa zabránilo spätnému toku vstrekovanej kvapaliny, nad airbagom sa urobí prepichnutie, po ktorom sú obidva otvory vyplnené parafínom.
Infekcia do alantoidnej dutiny. Infekcia sa uskutočňuje v zatemnenej škatuli. Zaznamená sa vzduchový priestor, dezinfikuje sa škrupina nad vzdušným priestorom a cez otvor v škrupine smerom k embryu sa vloží injekčná ihla s materiálom. Ak sa ihla dostane do alantoidnej dutiny, potom sa vytieni tieň embrya. Po infikovaní je otvor vyplnený parafínom.
Infekcia v žĺtkovom vaku. Škrupina je dezinfikovaná. Vajce sa položí na stojan tupým koncom doprava tak, aby žĺtkový vak smeroval nahor. Nad vzduchovou komorou v strede je prepichnutý otvor. Cez otvor v škrupine v horizontálnom smere do hĺbky 2 - 3 mm sa vloží ihla injekčnej striekačky, ktorá vstupuje do žĺtkového vaku. Materiál sa vstrekuje v objeme 0,2 - 0,3 ml. Po zavedení materiálu sa otvor voskuje.
Teplotný režim a doba inkubácie závisia od biologických vlastností zavedeného vírusu.
Infikované vajíčka sa kontrolujú každý deň - ovoskopia na kontrolu životaschopnosti embrya. Ak embryá zomrú prvý deň, príčinou je zvyčajne trauma počas infekcie. Takéto vajcia pochádzajú zo skúseností.
Ak je potrebné osobitne preskúmať každú zložku embrya, materiál sa zhromaždí v určitom poradí: alantoidná tekutina sa odsaje, potom sa odreže plodová voda, chorion-alantoidná membrána, oddelí sa plodová membrána, embryo, žĺtkový vak a až potom sa odstráni chorion-alantoidná membrána, ktorá ju oddeľuje od vnútornej povrch škrupiny. Prítomnosť vírusu v infikovanom embryu je určená charakteristickými zmenami v chorio-alantoidnej membráne infikovaného kuracieho embrya.
Vírusy, ktoré nemajú hemaglutinačnú aktivitu, sa detegujú pomocou CSC.
Na detekciu vírusu v alantoidných alebo plodových tekutinách infikovaných embryí sa umiestni RHA (hemaglutinácia je spôsobená alantoickými alebo plodovými tekutinami alebo suspenziou pripravenou z chorion-alantoidnej membrány).
Kultivácia vírusov v bunkovej kultúre... Na akumuláciu vírusov v citlivých bunkových kultúrach sa používajú tkanivá ľudí a rôznych zvierat. Najväčšie praktické uplatnenie dostali jednovrstvové kultúry primárnych trypsinizovaných a transplantovaných bunkových línií.
Jednovrstvové bunkové kultúry sa pestujú v plochých sklenených matracových nádobách. Bunková suspenzia v kvapalnom živnom médiu pri teplote 37 ° C umožňuje získať „in vitro“ vrstvu buniek s určitou histologickou štruktúrou. Prítomnosť vírusov v tkanivových kultúrach sa zisťuje podľa zmeny (degenerácie) buniek. Typ vírusov sa určuje neutralizáciou účinku vírusov pridaním vhodných typovo špecifických sér do očkovaného materiálu.
Tieto metódy umožňujú rýchlejšie zapracovanie výsledkov výskumu a sú nákladovo efektívnejšie. V prípadoch, keď vírusy nespôsobujú cytopatické pôsobenie (degeneráciu) a nevytvárajú sa v kuracích embryách, používajú sa metódy infikovania zvierat (pozri kapitolu 11).
Na kultiváciu vírusov sa používajú kontinuálne bunky, ktoré sa častejšie získavajú z buniek malígnych nádorov.
Jednovrstvové kultúry sa získavajú z ľudských, kuracích a zvieracích embryí.
Výhodou jednovrstvových bunkových kultúr je jednoduchosť techniky a jednoduchosť účtovníctva.
Schopnosť buniek reprodukovať sa mimo tela súvisí so stupňom diferenciácie tkanív. Menej diferencované tkanivá majú väčšiu schopnosť proliferácie (spojivové, epitelové tkanivo).
Podstatou metód na prípravu primárnych tkanivových kultúr je deštrukcia medzibunkového tkaniva a separácia buniek pre následnú výrobu monovrstvy.
Disociácia buniek sa uskutočňuje pôsobením proteolytických enzýmov, najčastejšie trypsínu, na tkanivo. Roztok trypsínu podporuje separáciu buniek pri zachovaní ich reprodukčnej schopnosti. Pestovanie bunkových kultúr si vyžaduje živné médium. Zloženie média je zložité, obsahuje množstvo zložiek: aminokyseliny, glukózu, vitamíny, minerálne soli, koenzýmy atď. Tkanivová kultúra sa získava za prísne aseptických podmienok. Na potlačenie rastu bakteriálnej flóry sa do média pridávajú antibiotiká (500 U penicilínu a 250 U streptomycínu v 1 ml).
Pripravené tkanivo sa naleje do 0,25% roztoku zohriateho trypsínu a inkubuje sa v termostate pri 37 ° C. Počas inkubácie sa tkanivo pravidelne mieša otáčaním banky. Trypsinizované bunky sa centrifugujú pri 800 až 1 000 ot./min počas 5 minút.
Trypsinizácia a centrifugácia sa vykonávajú veľmi opatrne, aby sa nepoškodili bunky. Po centrifugácii sa supernatant odstráni a bunkový sediment sa vloží do malého objemu kultivačného média. Na získanie homogénnej hmoty sa suspenzia buniek prefiltruje cez jednu vrstvu gázy v lieviku (sterilnom). Bunková suspenzia sa kontroluje na sterilitu inokuláciou 0,1 ml do 2 skúmaviek cukrovým bujónom.
Úspešnosť kultivácie buniek závisí od inokulačnej dávky, preto sa po trypsinizácii bunky spočítajú v Goryaevovej komore. Po spočítaní sa suspenzia buniek zriedi živným médiom tak, že 1 ml obsahuje 500 000 - 1 000 000 buniek a naleje sa do skúmaviek a matracov. Skúmavky na tkanivové kultúry sa inkubujú v inkubátore v naklonenej polohe.
Naočkované kultúry sa denne skúmajú pod mikroskopom s malým zväčšením, aby sa určila povaha ich rastu. Normálne proliferujúce bunky sú svetlo sfarbené a rastú v jednej vrstve. Ak sú bunky tmavé, zrnité a nerozmnožujú sa, čo môže byť výsledkom kontaminácie (zlé zaobchádzanie s riadmi alebo kontaminácia prísad), potom sa tieto kultúry z experimentu odstránia.
Výmena živného média 2 - 3 dni po zasiatí zlepšuje intenzitu premnoženia.
Normálne, dobre sa množiace bunky sú infikované testovaným materiálom.
Transplantované kultúry sa získavajú hlavne z malígnych nádorov. Kmeň Hela - kultúra buniek rakoviny krčka maternice ženy menom Helena (získaná v roku 1950); kmeň Hep-2 bol izolovaný od pacienta s rakovinou hrtana. Rast týchto buniek sa udržuje v laboratóriách postupnými pasážami. Ich zvláštnosť spočíva v tom, že sa dlho množia. V súčasnosti tieto bunky prešli tisíckami generácií. Pri prechode strácajú niektoré morfologické a biochemické vlastnosti - prechádzajú mutáciami. Na kultiváciu vírusov v nich však zostávajú celkom vhodné. Kultúru týchto buniek využívajú laboratóriá po celom svete.
K reprodukcii vírusu v bunkovej kultúre dochádza v rôznych časoch, v závislosti od vlastností vírusu a typu buniek.
Prítomnosť vírusu sa hodnotí podľa cytopatického účinku. V mikroskope sa pozoruje degenerácia buniek. Čas cytopatického pôsobenia a jeho podstata závisia od dávky a vlastností vírusu.
U niektorých vírusov sa cytopatický účinok zistí po niekoľkých dňoch (vírus kiahní), v iných po 1-2 týždňoch (vírus hepatitídy atď.).
V súčasnosti sú známe stovky vírusov, ktoré infikujú ľudí. Boj proti vírusovým infekciám sa uskutočňuje rôznymi metódami. Imunizácia je najúčinnejšia. Týmto spôsobom sa eliminovali kiahne a znížil sa výskyt poliomyelitídy. Sociálna prevencia je dôležitá v boji proti vírusovým infekciám - ničenie túlavých psov (boj proti besnote), osobná prevencia atď.
Tieto opatrenia však nemôžu zabezpečiť elimináciu všetkých vírusových ochorení. Vedci vytrvalo hľadajú spôsoby, ako by bolo možné infikovať vírus bez poškodenia bunky, v ktorej sa nachádza.
Je preto prirodzené, že v programe Komunistickej strany Sovietskeho zväzu je virológia označená za jedno z vedúcich odborov prírodnej vedy, ktoré by sa malo v nasledujúcich rokoch prednostne rozvíjať.
Základné metódy výskumu vírusov... 1. Reakcia hemaglutinácie, reakcia oneskorenej hemaglutinácie, reakcia nepriamej hemaglutinácie. Väzbová reakcia komplementu.
2. Reakcia neutralizácie vírusov v tkanivovej kultúre.
3. Metóda imunofluorescencie.
4. Histologická metóda - identifikácia inklúzií (Babeshove telíčka - Negri - s besnotou; Pashenove telíčka - s kiahňami atď.).
5. Biologická metóda.
Vírusová infekcia bunka je komplex procesov vznikajúcich pri interakcii bunky s vírusovým genómom. Extracelulárny virión je inertný a táto inertnosť pretrváva, kým vírusový genóm nezačne fungovať intracelulárne ako samostatná genetická jednotka. Kardinálnou vlastnosťou, ktorá je základom nižšie uvedenej klasifikácie, je vzťah medzi genómami vírusu a bunkou. Ak sa vírusový genóm replikuje nezávisle od replikácie bunkového genómu, ide o autonómny typ vírusovej infekcie. Ak je vírusový genóm zahrnutý do bunkového genómu a replikuje sa spolu s ním, jedná sa o integračný typ. Každý z týchto dvoch hlavných typov vírusovej infekcie možno rozdeliť do niekoľkých foriem podľa vzniku vírusu (produktívna a abortívna infekcia), podľa výsledku infekčného procesu pre bunku (lytická a nelytická infekcia), podľa dynamiky interakcie medzi vírusom a bunkou (akútna a chronická infekcia). ).
Autonómny typ vírusovej infekcie bunky je charakteristický pre väčšinu známych vírusov zvierat a ľudí. Pri tomto type infekcie sa vírusový genóm replikuje nezávisle od replikácie bunkového genómu.
Produktívna autonómna vírusová infekcia je charakterizovaná tvorbou úplného vírusu. Toto je najštudovanejšia forma interakcie medzi vírusmi a bunkami. Základné mechanizmy reprodukcie vírusov diskutované v kapitole 3 boli skúmané a opísané pre túto konkrétnu formu infekcie.
Autonómna abortívna vírusová infekcia nastáva, ak interakcia vírusu s bunkou nemá za následok tvorbu plne hodnotných viriónov. K tomu obvykle dochádza, keď sú bunky infikované chybným vírusom; keď vírus vstúpi do buniek rezistentných voči nemu; keď sú náchylné bunky infikované plne hodnotným vírusom za neprípustných podmienok.
Lytická autonómna vírusová infekcia vedie k bunkovej smrti. Vírus, ktorý spôsobuje bunkovú smrť, sa nazýva cytopatogénny a poškodenie buniek sa označuje pojmom „cytop, togénny účinok“. Povaha cytopatologických zmien vo vírusových infekciách je veľmi rôznorodá. Môže to byť degu
Zlyhanie buniek (vakuolizácia cytoplazmy, jadrová pyknóza, chromozomálne aberácie), tvorba špecifických vírusových intracelulárnych inklúzií, vznik obrovských buniek, ako sú polykaryocyty alebo symplasty. Lýza buniek je konečným štádiom pôsobenia cytopatogénnych vírusov. Pre prejav cytopatogenity vírus nevyžaduje reprodukciu genómu a implementáciu všetkých genetických informácií.
Nelytická autonómna vírusová infekcia je charakteristická pre vírusy, ktoré obsahujú lipoproteínový vonkajší obal a bunky vysielajú pučaním. Retro, ortomomyxo, paramyxo, arenavírusy sú schopné spôsobiť produktívnu nelytickú bunkovú infekciu. V najtypickejších prípadoch je však nelytická povaha charakteristická pre abortívne infekcie, keď genetické vlastnosti vírusov potrebné na implementáciu cytopatogénnych funkcií chýbajú alebo sa neprejavujú.
Podľa dynamiky infekčného procesu sa vírusové infekcie delia na akútne a chronické. Kedy akútna infekcia v infikovanej bunke nastáva tvorba viriónov, po ktorých sa infekčný proces zastaví. Pri chronickej infekcii infikovaná bunka produkuje virióny alebo vírusové zložky až do svojej prirodzenej smrti (nie z danej vírusovej infekcie), zatiaľ čo dcérske bunky zostávajú infikované. Produktívna a neúmyselná lytická infekcia je vždy akútna. Produktívna nelytická infekcia je vždy chronická. Abortívna nelytická infekcia má spravidla chronický priebeh, ale môže sa vyskytnúť aj v akútnej forme.
Integratívny typ vírusovej infekcie je charakterizovaný fyzickým spojením (integráciou) vírusových a bunkových genómov. V tomto prípade sa vírusový genóm replikuje a funguje ako neoddeliteľná súčasť bunkového genómu. Tento typ vírusovej infekcie je pre retrovírusy povinný a je možný v prípade vírusov papa, ado a herpes.
Integračné infekcie možno klasifikovať podľa rovnakých parametrov ako autonómne - produktivita, dynamika a výsledok; integračný vzťah medzi vírusom a bunkou môže byť tiež produktívny a prerušený. Pre retrovírusy je typická produktívna integračná infekcia. Interrupcia, integračná retrovírusová infekcia, sa vyskytuje za rovnakých podmienok, aké sú opísané vyššie pre autonómne abortívne infekcie. Takto sú defektné retrovírusy zbavené
Nové časti genómu (niektoré kmene vírusu Rousovho sarkómu, sarkómové vírusy myší, mačiek, opíc, leukemických vírusov myší Frend a Rauscher atď.) Môžu samy osebe spôsobiť iba abortívnu integračnú infekciu, ktorá sa za prítomnosti vírusu môže zmeniť na produktívnu infekciu. asistent. Príkladom abortívnej integračnej infekcie spôsobenej plne hodnotným retrovírusom v nepriepustných bunkách je infekcia vírus vtákov Rousov sarkóm buniek cicavcov. Zároveň v bunkách nastáva abortívny proces integrácie, ktorý sa vyznačuje zníženou produkciou vírusovej RNA, hromadením neúplného sortimentu vírusových proteínov, narušením ich spracovania a nedostatkom tvorby viriónov.
Integratívne infekcie sú zvyčajne nelytické, ako v prípade retrovírusov typu B, C, D, onkogénnych papov, adeno, herpes vírusov, existujú však aj lytické infekcie spôsobené cytopatogénnym retrovírusom vtáčej retikuloendoteliózy alebo „penivým“ vírusom.
Virogénna integračná infekcia je abortívna infekcia, ktorá sa dá preložiť do produktívnej formy. Bunky s touto formou infekcie môžu spontánne alebo pod vplyvom zvláštnych (fyzikálnych, chemických, biologických) vplyvov začať produkovať infekčný vírus. Tento jav sa nazýva virogénia a bunky sa označujú ako virogénne, analogicky s lyzogenézou a lyzogénnymi bunkami vo fágovo-bakteriálnom systéme.
Viac na tému Infekcia vírusovými bunkami:
- Vírusové infekcie a infekcie s podozrením na vírusovú etiológiu
- Bakteriálne a vírusové infekcie prenášané vzduchom: chrípka, parainfluenza, adenovírusová infekcia, respiračná syncyciálna infekcia. Bakteriálna bronchopneumónia, lobárna pneumónia.