VÍRUSY, najmenšie patogény infekčných chorôb. V preklade z latinčiny vírus znamená „jed, jedovatý princíp“. Do konca 19. storočia. pojem „vírus“ sa v medicíne používa na označenie každého infekčného agens, ktorý spôsobuje chorobu. Toto slovo získalo svoj moderný význam po roku 1892, keď ruský botanik DI Ivanovskij stanovil „filtrovateľnosť“ pôvodcu choroby tabakovej mozaiky (tabaková mozaika). Ukázal, že bunková šťava z rastlín infikovaných touto chorobou, prechádzajúca špeciálnymi filtrami, ktoré zachytávajú baktérie, si zachováva schopnosť spôsobiť rovnaké ochorenie aj u zdravých rastlín. O päť rokov neskôr nemecký bakteriológ F. Löffler objavil ďalšie filtrovateľné činidlo, pôvodcu slintačky a krívačky u hovädzieho dobytka. V roku 1898 holandský botanik M. Beijerinck zopakoval tieto experimenty v rozšírenej verzii a potvrdil závery Ivanovského. „Filtrovateľný jedovatý princíp“, ktorý spôsobuje tabakovú mozaiku, označil za „filtrovateľný vírus“. Tento výraz sa používa už mnoho rokov a postupne sa zredukoval na jediné slovo - „vírus“.
V roku 1901 americký vojenský chirurg W. Reed a jeho kolegovia zistili, že pôvodcom žltej zimnice je aj filtrovateľný vírus. Žltá zimnica bola prvým ľudským ochorením identifikovaným ako vírusové, trvalo však ďalších 26 rokov, kým sa jeho vírusový pôvod definitívne preukázal.
Všeobecne sa uznáva, že vírusy vznikli v dôsledku izolácie (autonómie) jednotlivých genetických prvkov bunky, ktoré navyše dostávali schopnosť prenosu z organizmu na organizmus. V normálnej bunke sa pohybuje niekoľko typov genetických štruktúr, napríklad matricová alebo informačná RNA (mRNA), transpozóny, intróny a plazmidy. Takéto mobilné prvky mohli byť predchodcami alebo predchodcami vírusov.
Sú vírusy živými organizmami? V roku 1935 izoloval americký biochemik W. Stanley vírus tabakovej mozaiky v kryštalickej forme, čím dokázal svoju molekulárnu povahu. Získané výsledky vyvolali búrlivú debatu o podstate vírusov: sú to živé organizmy alebo len aktivované molekuly? Vo vnútri infikovanej bunky sa vírusy skutočne prejavujú ako integrálna súčasť zložitejších živých systémov, ale mimo bunky sú to metabolicky inertné nukleoproteíny. Vírusy obsahujú genetickú informáciu, ale nemôžu ju samostatne implementovať bez vlastného mechanizmu syntézy proteínov. Keď sa objasnili vlastnosti štruktúry a reprodukcie vírusov, otázka, či sú živé, postupne strácala na význame.
ŠTRUKTÚRA VÍRUSOV
Kompletné v štruktúre a infekčné, t.j. schopná spôsobiť infekciu, vírusová častica mimo bunky sa nazýva virión. Jadro („jadro“) viriónu obsahuje jednu molekulu a niekedy dve alebo viac molekúl nukleovej kyseliny. Plášť proteínu, ktorý pokrýva nukleovú kyselinu viriónu a chráni ju pred škodlivými vplyvmi prostredia, sa nazýva kapsida. Nukleová kyselina viriónu je genetický materiál vírusu (jeho genómu) a je predstavovaná deoxyribonukleovou kyselinou (DNA) alebo ribonukleovou kyselinou (RNA), nikdy však týmito dvoma zlúčeninami naraz. (Chlamydia, rickettsia a všetky ostatné „skutočne živé“ mikroorganizmy obsahujú DNA aj RNA.) Nukleové kyseliny najmenších vírusov obsahujú tri alebo štyri gény, zatiaľ čo najväčšie vírusy majú až sto génov.
Niektoré vírusy majú okrem kapsidy aj vonkajší obal pozostávajúci z bielkovín a lipidov. Je tvorený z membrán infikovanej bunky obsahujúcich zabudované vírusové proteíny. Pojmy „nahé virióny“ a „virióny bez plášťa“ sa používajú synonymne. Kapsidy najmenších a najjednoduchšie usporiadaných vírusov môžu pozostávať iba z jedného alebo niekoľkých typov molekúl bielkovín. Niekoľko molekúl rovnakých alebo rôznych proteínov je kombinovaných do podjednotiek nazývaných kapsoméry. Kapsoméry zase tvoria pravidelné geometrické štruktúry vírusovej kapsidy. U rôznych vírusov je tvar kapsidy charakteristickým znakom (znakom) viriónu.
Virióny so špirálovým typom symetrie, ako napríklad vírus tabakovej mozaiky, majú tvar podlhovastého valca; vo vnútri plášťa proteínu, ktorý sa skladá z jednotlivých podjednotiek - kapsomérov, sa nachádza špirálovitá špirála nukleovej kyseliny (RNA). Virióny s ikosahedrálnym typom symetrie (z gréckeho eikosi - dvadsať, hedra - povrch), podobne ako poliovírus, majú sférický, alebo skôr mnohostranný tvar; ich kapsidy sú postavené z 20 pravidelných trojuholníkových faziet (povrchov) a vyzerajú ako geodetická kupola.
Jednotlivé bakteriofágy (bakteriálne vírusy; fágy) majú zmiešaný typ symetrie. Na tzv. Z „chvostových“ fágov vyzerá hlava ako sférická kapsida; dlhý rúrkovitý proces - "chvost" sa z neho odchyľuje.
Existujú vírusy s ešte zložitejšou štruktúrou. Vírusy kiahní (vírusy kiahní) nemajú pravidelnú typickú kapsidu: medzi jadrom a vonkajším plášťom sú umiestnené tubulárne a membránové štruktúry.
Genetické informácie kódované v jednom géne možno všeobecne chápať ako pokyny na produkciu špecifického proteínu v bunke. Takáto inštrukcia je bunkou vnímaná, iba ak je odoslaná vo forme mRNA. Preto bunky, v ktorých je genetický materiál reprezentovaný DNA, musia tieto informácie „prepisovať“ (prepisovať) do komplementárnej kópie proteínov vírusu mRNA (pozri tiež NUKLEOVÉ KYSELINY). Vírusy obsahujúce DNA sa spôsobom replikácie líšia od vírusov obsahujúcich RNA.
DNA zvyčajne existuje vo forme dvojvláknových štruktúr: dva polynukleotidové reťazce sú viazané vodíkom a skrútené takým spôsobom, že vzniká dvojitá špirála. Naproti tomu RNA zvyčajne existuje vo forme jednovláknových štruktúr. Genómom jednotlivých vírusov je však buď jednovláknová DNA, alebo dvojvláknová RNA. Pramene (reťazce) vírusovej nukleovej kyseliny, dvojité alebo jednoduché, môžu byť lineárne alebo uzavreté v kruhu.
Prvý stupeň vírusovej replikácie je spojený s penetráciou vírusovej nukleovej kyseliny do hostiteľskej bunky. Tento proces môžu uľahčiť špeciálne enzýmy, ktoré sú súčasťou kapsidy alebo vonkajšieho obalu viriónu, a obal zostáva mimo bunky alebo ho virión stratí okamžite po preniknutí do bunky. Vírus nájde bunku vhodnú na svoju reprodukciu kontaktom jednotlivých častí svojej kapsidy (alebo vonkajšieho obalu) so špecifickými receptormi na povrchu bunky spôsobom „uzamknutia kľúčom“. Ak na povrchu bunky chýbajú špecifické („rozpoznávajúce“) receptory, potom bunka nie je citlivá na vírusovú infekciu: vírus do nej nepreniká.
Za účelom realizácie svojej genetickej informácie je vírusová DNA, ktorá prenikla do bunky, transkribovaná špeciálnymi enzýmami do mRNA. Výsledná mRNA sa presúva do bunkových „tovární“ na syntézu proteínov - ribozómov, kde nahrádza bunkové „správy“ vlastnými „pokynmi“ a je prekladaná (čítaná), v dôsledku čoho sú syntetizované vírusové proteíny. Samotná vírusová DNA je opakovane duplikovaná (duplikovaná) za účasti ďalšej sady enzýmov, vírusových aj patriacich k bunke.
Syntetizovaný proteín, ktorý sa používa na vytvorenie kapsidy, a vírusová DNA namnožená v mnohých kópiách sa spájajú a vytvárajú nové „dcérske“ virióny. Vytvorený vírusový potomok opustí použitú bunku a infikuje nové: cyklus reprodukcie vírusu sa opakuje. Niektoré vírusy počas pučania z bunkového povrchu zachytia časť bunkovej membrány, do ktorej boli vírusové proteíny vložené „vopred“, a získajú tak membránu. Pokiaľ ide o hostiteľskú bunku, nakoniec sa ukáže, že je poškodená alebo dokonca úplne zničená.
U niektorých DNA vírusov nie je samotný reprodukčný cyklus v bunke spojený s okamžitou replikáciou vírusovej DNA; namiesto toho sa vírusová DNA vloží (integruje) do DNA hostiteľskej bunky. V tomto štádiu vírus ako jedna štrukturálna entita zmizne: jeho genóm sa stáva súčasťou genetického aparátu bunky a počas delenia buniek sa dokonca replikuje ako súčasť bunkovej DNA. Avšak následne, niekedy po mnohých rokoch, sa vírus môže znovu objaviť - spúšťa sa mechanizmus syntézy vírusových proteínov, ktoré v kombinácii s vírusovou DNA vytvárajú nové virióny.
V niektorých RNA vírusoch môže genóm (RNA) priamo pôsobiť ako mRNA. Táto vlastnosť je však typická iba pre vírusy s reťazcom „+“ RNA (tj. S RNA s pozitívnou polaritou). Vo vírusoch s reťazcom „-“ RNA musí byť vlákno „najskôr“ prepísané do vlákna „+“; až potom začne syntéza vírusových proteínov a vírus sa replikuje.
Takzvané retrovírusy obsahujú ako svoj genóm RNA a majú neobvyklý spôsob prepisu genetického materiálu: namiesto prepisu DNA na RNA, ako sa to deje v bunke a je to typické pre vírusy obsahujúce DNA, sa ich RNA prepisuje do DNA. Dvojvláknová DNA vírusu sa potom začlení do chromozomálnej DNA bunky. Na matrici takejto vírusovej DNA sa syntetizuje nová vírusová RNA, ktorá rovnako ako iné určuje syntézu vírusových proteínov. Pozri tiež RETROVIRÚZY.
Ak sú vírusy skutočne mobilné genetické prvky, ktoré dostali „autonómiu“ (nezávislosť) od genetického aparátu svojich hostiteľov (rôzne typy buniek), potom mali vzniknúť nezávisle na sebe rôzne skupiny vírusov (s rôznym genómom, štruktúrou a replikáciou). Preto je nemožné vytvoriť pre všetky vírusy jeden rodokmeň, ktorý by ich spájal na základe evolučných vzťahov. Princípy „prirodzenej“ klasifikácie používané v taxonómii zvierat nie sú vhodné pre vírusy.
Napriek tomu je systém klasifikácie vírusov v praxi nevyhnutný a pokusy o jeho vytvorenie sa opakovane pokúšali. Ukázalo sa, že najproduktívnejší prístup je založený na štrukturálnych a funkčných charakteristikách vírusov: na odlíšenie rôznych skupín vírusov navzájom popisujú typ svojej nukleovej kyseliny (DNA alebo RNA, z ktorých každá môže byť jednovláknová alebo dvojvláknová), jej veľkosť (počet nukleotidov v reťazci nukleovej kyseliny). kyseliny), počet molekúl nukleovej kyseliny v jednom virióne, geometria viriónu a štrukturálne vlastnosti kapsidy a vonkajšieho obalu viriónu, typ hostiteľa (rastliny, baktérie, hmyz, cicavce atď.), vlastnosti patológie vyvolanej vírusmi (príznaky a podstata ochorenia), antigénne vlastnosti vírusových proteínov a vlastnosti reakcie imunitného systému tela na zavedenie vírusu.
Skupina mikroskopických patogénov nazývaných viroidy (t.j. častice podobné vírusom) úplne nezapadá do klasifikačného systému vírusov. Viroidy spôsobujú veľa bežných chorôb rastlín. Toto sú najmenšie infekčné agensy, ktoré neobsahujú ani najjednoduchšie proteínové obaly (vyskytujú sa vo všetkých vírusoch); pozostávajú iba z jednovláknovej RNA uzavretej v kruhu.
Vírusové choroby
Pre mnoho vírusov, ako sú osýpky, herpes a čiastočne chrípka, sú ľudia hlavným prírodným rezervoárom. K prenosu týchto vírusov dochádza vzdušnými kvapôčkami alebo kontaktom.
Distribúcia niektorých vírusové ochorenia, rovnako ako iné infekcie, je plný prekvapení. Napríklad v skupinách ľudí žijúcich v nehygienických podmienkach takmer všetky deti v nízky vek niesť poliomyelitídu, zvyčajne miernu, a získať imunitu. Ak sa zlepšia životné podmienky v týchto skupinách, deti mladší vek detská obrna nie je zvyčajne chorá, ale choroba sa môže vyskytnúť vo vyššom veku a potom je často závažná.
Mnoho vírusov nemôže v prírode dlho pretrvávať pri nízkej hustote rozptýlenia hostiteľského druhu. Nedostatok populácií primitívnych lovcov a zberateľov rastlín vytvoril nepriaznivé podmienky pre existenciu niektorých vírusov; preto je veľmi pravdepodobné, že niektoré ľudské vírusy sa objavili neskôr, keď sa objavili mestské a vidiecke sídla. Predpokladá sa, že vírus osýpok pôvodne existoval u psov (ako pôvodca horúčky) a ovčie kiahne u ľudí sa mohli objaviť v dôsledku vývoja kiahní u kráv alebo myší. Medzi najnovšie príklady vývoja vírusov patrí syndróm získanej ľudskej imunodeficiencie (AIDS). Existujú dôkazy o genetických podobnostiach medzi vírusmi ľudskej imunodeficiencie a africkými zelenými opicami.
„Nové“ infekcie sú zvyčajne závažné, často smrteľné, ale počas vývoja patogénu sa môžu stať ľahšími. Dobrým príkladom je história vírusu myxomatózy. V roku 1950 bol tento vírus, endemický pre Južnú Ameriku a skôr neškodný pre miestne králiky, zavedený do Austrálie spolu s európskymi plemenami týchto zvierat. Choroba u austrálskych králikov, ktorá sa predtým s týmto vírusom nestretla, bola smrteľná v 99,5% prípadov. O niekoľko rokov neskôr sa úmrtnosť na toto ochorenie významne znížila, v niektorých oblastiach až na 50%, čo sa vysvetľuje nielen „zoslabením“ (oslabením) mutácií vírusového genómu, ale aj zvýšenou genetickou odolnosťou králikov voči tejto chorobe a v obidvoch prípadoch došlo k účinnému prirodzenému výberu za silný tlak prírodného výberu.
Reprodukciu vírusov v prírode podporujú rôzne druhy organizmov: baktérie, huby, prvoky, rastliny, zvieratá. Napríklad hmyz často trpí vírusmi, ktoré sa hromadia v ich bunkách vo forme veľkých kryštálov. Rastliny sú často napadnuté malými a jednoducho usporiadanými RNA vírusmi. Tieto vírusy nemajú ani špeciálne mechanizmy na vstup do bunky. Prenášajú ich hmyz (živiaci sa bunkovou šťavou), škrkavky a kontaktom, ktoré infikujú rastlinu, ak je mechanicky poškodená. Bakteriálne vírusy (bakteriofágy) majú najkomplexnejší mechanizmus dodania svojho genetického materiálu do citlivej bakteriálnej bunky. Najskôr sa fágový „chvost“, ktorý vyzerá ako tenká trubica, pripája k stene baktérie. Potom špeciálne enzýmy „chvosta“ rozpustia časť bakteriálnej steny a fágový genetický materiál (zvyčajne DNA) sa vstrekne do výsledného otvoru cez „chvost“, ako cez ihlu injekčnej striekačky.
Viac ako desať hlavných skupín vírusov je pre ľudí patogénnych. Medzi vírusmi obsahujúcimi DNA je to rodina poxvírusov (spôsobujúcich ovčie kiahne, vakcínie a iné ovčie kiahne), vírusy skupiny herpes (opary na perách, ovčie kiahne), adenovírusy (choroby) dýchacích ciest a oči), rodina papovavírusov (bradavice a iné výrastky na koži), hepadnavírusy (vírus hepatitídy B). Existuje oveľa viac vírusov obsahujúcich RNA, ktoré sú pre človeka patogénne. Pikornavírusy (z latinčiny pico - veľmi malé, anglické RNA - RNA) sú najmenšie vírusy cicavcov, podobné niektorým rastlinným vírusom; spôsobujú poliomyelitídu, hepatitídu A, akútnu prechladnutie... Mixovírusy a paramyxovírusy sú pôvodcami rôznych foriem chrípky, osýpok a príušníc. Arbovírusy (z angličtiny prenášané článkonožcami - „prenášané článkonožcami“) - najväčšia skupina vírusov (viac ako 300) - sú prenášané hmyzom a sú pôvodcami kliešťovej a japonskej encefalitídy, žltej zimnice, meningoencefalitídy koní, horúčky spôsobenej kliešťami v Colorade, škótskej encefalitídy a iných chorôb ... Reovírusy, ktoré sú skôr zriedkavými pôvodcami respiračných a črevných chorôb u ľudí, sa stali predmetom osobitného vedeckého záujmu vďaka skutočnosti, že ich genetický materiál je predstavovaný dvojvláknovou fragmentovanou RNA. Pozri tiež VENERÁLNE CHOROBY; OVČIE KIAHNE; HEPATITÍDA; CHRÍPKA; HLOUBKA HORÚČKA; MONONUKLEÓZA INFEKČNÁ; MEASLES; KRASNUHA; Meningitída; PRÍRODNÝ POISK; POLIO; DÝCHACIE VIRÁLNE CHOROBY; PRASIATKO; SYNDROM ZÍSKANEJ IMUNODEFICIENCIE (AIDS); ENCEFALITÍDA.
Pôvodcovia niektorých chorôb, vrátane veľmi závažných, nepatria do žiadnej z vyššie uvedených kategórií. Do zvláštnej skupiny pomalých vírusové infekcie nedávno sa uvažovalo napríklad o Creutzfeldt-Jakobovej chorobe a Kuru, degeneratívnych ochoreniach mozgu s veľmi dlhou inkubačnou dobou. Ukázalo sa však, že ich pôvodcom nie sú vírusy, ale najmenší infekční agens bielkovinovej povahy - prióny (pozri PRION).
Liečba a prevencia. Reprodukcia vírusov je úzko prepojená s mechanizmami syntézy bielkovín a nukleových kyselín bunky v infikovanom organizme. Preto je výroba liekov, ktoré selektívne potláčajú vírus, ale nepoškodzujú telo, mimoriadne náročnou úlohou. Napriek tomu sa ukázalo, že u najväčších vírusov herpesu a kiahní kóduje genómová DNA veľké množstvo enzýmov, ktoré sa líšia svojimi vlastnosťami od podobných bunkových enzýmov, a to slúžilo ako základ pre vývoj antivírusové lieky... Skutočne bolo vyvinutých niekoľko liekov, ktorých mechanizmus účinku je založený na potlačení syntézy vírusovej DNA. Niektoré zlúčeniny, ktoré sú príliš toxické na všeobecné použitie (intravenózne alebo ústami), sú vhodné na miestne použitie, napríklad na očné infekcie vírusom herpes.
Je známe, že ľudské telo produkuje špeciálne proteíny - interferóny. Potlačujú transláciu vírusových nukleových kyselín a tým brzdia reprodukciu vírusu. Vďaka genetickému inžinierstvu sa interferóny produkované baktériami stali dostupnými a sú testované v lekárskej praxi (pozri GENE ENGINEERING).
Medzi najúčinnejšie prvky prirodzenej obrany tela patria špecifické protilátky (špeciálne proteíny produkované imunitným systémom), ktoré interagujú s príslušným vírusom a tým účinne bránia rozvoju choroby; nemôžu však neutralizovať vírus, ktorý sa už dostal do bunky. Príkladom je herpetická infekcia: Herpetický vírus pretrváva v bunkách nervových uzlín (ganglií), kam sa protilátky nemôžu dostať. Vírus sa z času na čas aktivuje a spôsobí relapsy choroby.
Zvyčajne sa v tele vytvárajú špecifické protilátky v dôsledku penetrácie infekčného agens. Telu môže pomôcť umelé zvýšenie produkcie protilátok, vrátane budovania imunity vopred, očkovaním. Práve týmto spôsobom sa prostredníctvom masového očkovania na celom svete prakticky eliminovala choroba kiahní. Pozri tiež OČKOVANIE A IMUNIZÁCIA.
Moderné metódy očkovania a imunizácie sú rozdelené do troch hlavných skupín. Po prvé, ide o použitie oslabeného kmeňa vírusu, ktorý stimuluje produkciu protilátok v tele, ktoré sú účinné proti patogénnejšiemu kmeňu. Po druhé, zavedenie zabitého vírusu (napríklad inaktivovaného formalínom), ktorý tiež indukuje tvorbu protilátok. Treťou možnosťou je tzv. „Pasívna“ imunizácia, t.j. zavedenie hotových "cudzích" protilátok. Zviera, napríklad kôň, je imunizované, potom sú z jeho krvi izolované protilátky, purifikované a použité na podanie pacientovi na vytvorenie okamžitej, ale krátkodobej imunity. Niekedy sa protilátky používajú z krvi človeka, ktorý ochorel (napríklad osýpky, kliešťová encefalitída).
Hromadenie vírusov. Pri príprave vakcín je potrebné akumulovať vírus. Na tento účel sa často používajú vyvíjajúce sa kuracie embryá, ktoré sú infikované týmto vírusom. Po inkubácii infikovaných embryí po určitý čas sa zhromaždí vírus, ktorý sa v nich nahromadí v dôsledku reprodukcie, vyčistí sa (centrifugáciou alebo iným spôsobom) a v prípade potreby sa deaktivuje. Je veľmi dôležité z prípravkov vírusu odstrániť všetky balastné nečistoty, ktoré môžu počas očkovania spôsobiť vážne komplikácie. Rovnako dôležité je samozrejme zaistiť, aby v prípravkoch nezostal žiadny neinaktivovaný patogénny vírus. V posledných rokoch sa na akumuláciu vírusov široko používajú rôzne typy bunkových kultúr.
METÓDY ŠTÚDIA VÍRUSOV
Bakteriálne vírusy sa ako prvé stali predmetom podrobných štúdií ako najpohodlnejší model, ktorý má oproti iným vírusom množstvo výhod. Celý cyklus replikácie fágov, t.j. doba od infekcie bakteriálnej bunky do uvoľnenia rozmnožených vírusových častíc z nej nastáva do jednej hodiny. Ďalšie vírusy sa zvyčajne hromadia počas niekoľkých dní alebo dokonca dlhšie. Krátko pred druhou svetovou vojnou a krátko po jej skončení boli vyvinuté metódy na štúdium jednotlivých vírusových častíc. Doštičky s výživným agarom, na ktorých bola pestovaná monovrstva (súvislá vrstva) bakteriálnych buniek, sú infikované fágovými časticami pomocou svojich sériových zriedení. Počas množenia vírus zabíja „chránenú“ bunku a preniká do susedných buniek, ktoré tiež zomierajú po nahromadení potomkov fágov. Oblasť mŕtvych buniek je viditeľná voľným okom ako svetlá škvrna. Takéto škvrny sa nazývajú „negatívne kolónie“ alebo plaky. Vyvinutá metóda umožnila študovať potomstvo jednotlivých vírusových častíc, detegovať genetickú rekombináciu vírusov a určiť genetickú štruktúru a metódy replikácie fágov v detailoch, ktoré sa predtým zdali neuveriteľné.
Práca s bakteriofágmi prispela k rozšíreniu metodického arzenálu pri štúdiu živočíšnych vírusov. Predtým sa štúdie vírusov stavovcov uskutočňovali hlavne na laboratórnych zvieratách; také experimenty boli veľmi namáhavé, drahé a málo poučné. Následne sa objavili nové metódy založené na použití tkanivových kultúr; bakteriálne bunky použité vo fágových experimentoch boli nahradené bunkami stavovcov. Pre štúdium mechanizmov vývoja vírusových chorôb sú však veľmi dôležité experimenty na laboratórnych zvieratách, ktoré sa uskutočňujú aj v súčasnosti.
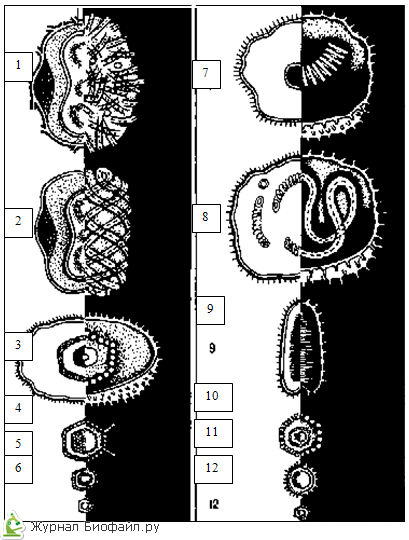
U ICOSAHEDRICKÉHO TYPU SYMETRIE, znázorneného na schéme štruktúry adenovírusu, kapsomérov alebo proteínových podjednotiek vírusu, tvoria izometrický proteínový obal pozostávajúci z 20 pravidelných trojuholníkov.
V PRÍPADE ŠPIRÁLOVEJ SYMETRIE, ktorá je znázornená na schéme štruktúry vírusu tabakovej mozaiky, tvoria kapsoméry alebo podjednotky vírusu okolo dutého rúrkovitého jadra špirálu.
KOMBINOVANÚ alebo zmiešanú symetriu vo vírusoch môžu predstavovať rôzne možnosti. Častica bakteriofága znázornená na diagrame má „hlavu“ pravidelného geometrického tvaru a „chvost“ so špirálovitou symetriou.
Kryštalizácia vírusov
V roku 1932 bol mladý americký biochemik Wendill Stanley požiadaný, aby študoval vírusy. Stanley začal stlačením fľaše džúsu z tony tabakových listov infikovaných vírusom tabakovej mozaiky. Začal vyšetrovať šťavu pomocou chemických metód, ktoré mal k dispozícii. Vystavil rôzne frakcie šťavy rôznym činidlám v nádeji, že získa čistý vírusový proteín (Stanley bol presvedčený, že vírus je proteín). Stanley kedysi dostával takmer čistú frakciu proteínu, ktorý sa svojím zložením líšil od proteínov rastlinných buniek. Vedec si uvedomil, že pred ním je to, o čo sa tak usilovne usiloval. Stanley izoloval mimoriadny proteín, rozpustil ho vo vode a roztok umiestnil do chladničky. Nasledujúce ráno namiesto toho v banke číra tekutina boli tam krásne hodvábne ihličkovité kryštály. Stanley získal lyžicu týchto kryštálov z tony listov. Potom Stanley naliala niekoľko kryštálov, rozpustila ich vo vode, navlhčila touto vodou gázu a potrela ňou listy zdravých rastlín. Rastlinná šťava prešla celým radom chemických vplyvov. Po takomto „masívnom spracovaní“ vírusy s najväčšou pravdepodobnosťou mali zomrieť.
Natierané listy sú choré. Takže zvláštne vlastnosti vírusu boli doplnené o ďalšiu - schopnosť kryštalizovať.
Kryštalizačný efekt bol taký ohromujúci, že Stanley sa na dlhú dobu vzdal myšlienky, že vírus je tvor. Pretože všetky enzýmy sú bielkoviny a veľa enzýmov sa zvyšuje aj s vývojom tela a môžu kryštalizovať, dospel Stanley k záveru, že vírusy sú čisté bielkoviny, skôr enzýmy.
Vedci sa čoskoro presvedčili, že je možné kryštalizovať nielen vírus tabakovej mozaiky, ale aj množstvo ďalších vírusov.
O päť rokov neskôr anglickí biochemici F. Bowden a N. Peary našli chybu v Stanleyho definícii. 94% obsahu vírusu tabakovej mozaiky pozostávalo z bielkovín a 6% tvorila nukleová kyselina. Vírus nebol vlastne proteín, ale nukleoproteín - kombinácia proteínu a nukleovej kyseliny.
Hneď ako boli biológom k dispozícii elektrónové mikroskopy, vedci zistili, že vírusové kryštály pozostávajú z niekoľkých stoviek miliárd častíc pevne stlačených k sebe. V jednom kryštáli vírusu poliomyelitídy je toľko častíc, že \u200b\u200bmôžu infikovať všetkých obyvateľov Zeme viackrát. Keď bolo možné skúmať jednotlivé vírusové častice v elektrónovom mikroskope, ukázalo sa, že majú rôzne tvary, ale vonkajší obal vírusov vždy pozostáva z proteínov, ktoré sa líšia v rôznych vírusoch, čo umožňuje ich rozpoznanie pomocou imunologických reakcií a vnútorný obsah predstavuje nukleová kyselina, ktorá je jednotka dedičnosti.
Zložky vírusovNajväčšie vírusy (vírusy kiahní) sa veľkosťou približujú malým baktériám, najmenšie (pôvodcovia encefalitídy, poliomyelitídy, slintačky a krívačky) - veľkým molekulám bielkovín. Inými slovami, vírusy majú svojich vlastných obrov a trpaslíkov. (viď obr. 1) Na meranie vírusov sa používa konvencia nazývaná nanometer (nm). Jedna nm je milióntina milimetra. Veľkosti rôznych vírusov sa pohybujú od 20 do 300 nm.
Vírusy sa teda skladajú z niekoľkých zložiek:
jadro - genetický materiál (DNA alebo RNA). Genetický aparát vírusu nesie informácie o niekoľkých druhoch proteínov, ktoré sú potrebné na tvorbu nového vírusu.
bielkovinový obal, ktorý sa nazýva kapsida. Obálka je často vyrobená z identických opakujúcich sa podjednotiek - kapsomérov. Capsomeres tvoria štruktúry s vysokým stupňom symetrie.
Dodatočná lipoproteínová membrána. Je tvorený z plazmatickej membrány hostiteľskej bunky. Nachádza sa iba v relatívne veľkých vírusoch (chrípka, opar). Tento vonkajší obal je fragmentom jadrovej alebo cytoplazmatickej membrány hostiteľskej bunky, z ktorej vírus vstupuje do extracelulárneho prostredia. Niekedy vo vonkajších obaloch komplexných vírusov sú okrem bielkovín obsiahnuté aj sacharidy, napríklad v pôvodcoch chrípky a oparu.
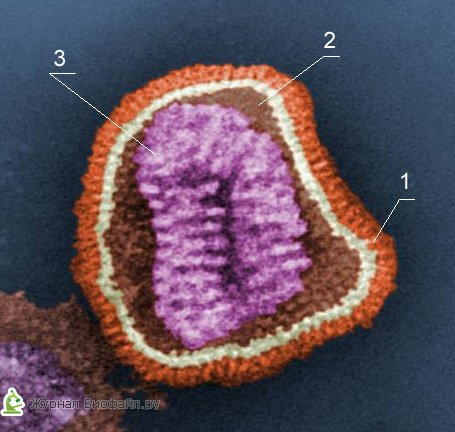
1. Dodatočná škrupina
2. Capsomer (bielkovinový obal)
3. Jadro (DNA alebo RNA)
Každá zložka viriónov má špecifické funkcie: bielkovinový obal ich chráni pred nepriaznivými účinkami, nukleová kyselina je zodpovedná za dedičné a infekčné vlastnosti a hrá vedúcu úlohu pri variabilite vírusov a na ich reprodukcii sa podieľajú enzýmy.
Štrukturálne zložitejšie vírusy obsahujú okrem bielkovín a nukleových kyselín aj sacharidy a lipidy. Každá skupina vírusov má svoju vlastnú skupinu bielkovín, tukov, sacharidov a nukleových kyselín. Niektoré vírusy obsahujú vo svojom zložení enzýmy.
Na rozdiel od bežných živých buniek vírusy nespotrebúvajú jedlo ani neprodukujú energiu. Nie sú schopné reprodukcie bez účasti živej bunky. Vírus sa začína množiť až potom, čo vstúpi do určitého typu bunky. Napríklad vírus poliomyelitídy môže žiť iba v nervových bunkách ľudí alebo vysoko organizovaných zvierat, ako sú napr opice. Bakteriálne vírusy majú trochu inú štruktúru.
Interakcia vírusu s bunkouVírusy mimo bunky sú kryštály, ale keď vstúpia do bunky, „ožijú“. K ich reprodukcii dochádza zvláštnym, neporovnateľným spôsobom. Najskôr do bunky vstúpia virióny a uvoľnia sa vírusové nukleové kyseliny. Potom sa „pozbierajú“ podrobnosti o budúcich viriónoch. Rozmnožovanie sa končí zhromaždením nových viriónov a ich uvoľnením do životného prostredia.
Stretnutie vírusov s bunkami sa začína jeho adsorpciou, to znamená pripojením k bunkovej stene. Potom sa začne zavedenie alebo prenikanie viriónu do bunky, čo sama vykonáva. Avšak penetrácii vírusu do cytoplazmy bunky spravidla predchádza jeho väzba so špeciálnym receptorovým proteínom umiestneným na povrchu bunky. Väzba na receptor sa uskutočňuje vďaka prítomnosti špeciálnych proteínov na povrchu vírusovej častice, ktoré „rozpoznávajú“ zodpovedajúci receptor na povrchu citlivej bunky. Na jednej bunke môžu byť adsorbované desiatky až stovky viriónov. Plocha povrchu bunky, ku ktorej sa vírus pripojil, sa ponorí do cytoplazmy a zmení sa na vakuolu. Vakuola, ktorej stena pozostáva z cytoplazmatickej membrány, môže splývať s inými vakuolami alebo s jadrom. Takto sa vírus dodáva do ktorejkoľvek časti bunky. Tento proces sa nazýva viropexis.
Infekčný proces začína, keď sa začnú množiť vírusy, ktoré sa dostali do bunky, t.j. dochádza k reduplikácii vírusového genómu a samomontáži kapsidy. Aby mohlo dôjsť k duplikácii, musí sa nukleová kyselina uvoľňovať z kapsidy. Po syntéze novej molekuly nukleovej kyseliny sa upraví a z vírusových proteínov syntetizovaných v cytoplazme bunky sa vytvorí kapsida. Hromadenie vírusových častíc vedie k ich výstupu z bunky. U niektorých vírusov k tomu dôjde „výbuchom“, v dôsledku čoho dôjde k narušeniu celistvosti bunky a jej odumretiu. Ostatné vírusy sa vylučujú podobným spôsobom ako pučanie. V tomto prípade si bunky tela môžu dlho udržiavať svoju životaschopnosť.
Ďalším spôsobom penetrácie do bunky v bakteriofágoch. Silné bunkové steny neumožňujú, aby sa receptorový proteín spolu s vírusom k nemu pripojeným ponoril do cytoplazmy, ako to býva pri infikovaní živočíšnych buniek. Preto bakteriofág vloží do bunky dutú tyč a pretláča cez ňu DNA (alebo RNA), ktorá je v jej hlave. Genóm bakteriofága vstupuje do cytoplazmy, zatiaľ čo kapsid zostáva vonku. V cytoplazme bakteriálnej bunky začína replikácia bakteriofágového genómu, syntéza jeho proteínov a tvorba kapsidy. Po určitom čase bakteriálna bunka zomrie a zrelé častice fága sa uvoľnia do životného prostredia.
Je úžasné, ako vírusy, ktoré sú desaťkrát až stokrát menšie ako bunky, šikovne a sebavedome riadia bunkovú ekonomiku. Pri množení vyčerpávajú bunkové zdroje a hlboko, často nezvratne, narúšajú metabolizmus, čo v konečnom dôsledku spôsobuje bunkovú smrť.
Forma rastlinných vírusov je zvyčajne tyčinkovitá a okrúhla. Rozmery vírusov v tvare tyčinky sú 300 - 480 x 15 nm a tie, ktoré majú zaoblený tvar, sú 25 - 30 nm. [...]
Ide o mikroorganizmy, ktoré nemajú bunkovú štruktúru. Veľkosti štruktúrnych jednotiek vírusov (viriónov) sa pohybujú od 10 do 300 nm. Medzi virióny patria molekuly ribonukleových (RNA) alebo deoxyribonukleových (DNA) kyselín obklopené proteínovým obalom. Vírusy majú rôzne tvary: kubický, sférický, tyčinkovitý atď. Reprodukcia vírusov sa uskutočňuje jednoduchým delením alebo zložitejším spôsobom iba v bunkách živého organizmu. Vírusy pôsobia špecificky, to znamená, že určité skupiny vírusov infikujú určité živé organizmy. [...]
Vírusy, ktoré sú menšie a menej zložité ako bunky, nemôžu žiť nezávisle. Sú to len veľmi zvláštne zabalené kúsky genetickej informácie, ktoré môžu žiť a množiť sa iba infikovaním bunky. V takom prípade môžu v jednej bunke vzniknúť tisíce vírusových častíc. Predpokladá sa, že vírusy nejako podrobia mechanizmus bunkového života a použijú ho na svoje vlastné účely. Evolučný pôvod vírusov nie je celkom jasný. Môžu sa považovať za vysoko generované bunky alebo ich fragmenty. Gény vírusov sú podobné génom iných foriem a môžu byť tiež mutované. [...]
Tento vírus obsahuje asi 20% RNA a jeho častice sú mnohosteny. Priemer častíc na elektrónových mikrofotografiách získaných negatívnym kontrastom je od 26 do 30 nm, čo závisí od charakteristík prípravku. Podrobná štruktúra tohto vírusu nie je jasná, je však predmetom značného záujmu, pretože niektoré izoláty obsahujú satelitný vírus spojený s týmto vírusom, ktorý je opísaný nižšie, ako aj v [...]
Ďalšia forma závislosti je charakteristická pre satelitný vírus vírusu nekrózy tabaku. Je to najmenší známy vírus. Jeho RNA obsahuje množstvo informácií dostatočné na kódovanie jeho vlastného kapsidového proteínu a prípadne špecifickej RIK polymázy. Pre ďalšie významné, ale stále neznáme funkcie, op závisí od prítomnosti nepríbuzného vírusu nekrózy tabaku. [...]
Virión vírusu je vláknitý, s veľkosťou 600-700 X 12 mikrónov, inaktivovaný pri 60-67 ° C, odoláva zmrazeniu. Dopravca nie je známy. [...]
Častice vírusu lucerny mozaiky (ALV) sa líšia od ostatných rastlinných vírusov svojím baciloformným tvarom. Štruktúra týchto vírusov má niektoré vlastnosti, ktoré sú charakteristické pre tyčinkovité aj izometrické vírusy. Z vírusového prípravku VML sa izolovalo päť zložiek (b0, la, bb, M a B). Zistilo sa, že najmenej štyri z nich sú nevyhnutné na vznik infekcie (kap. [...]
Veľkosť a tvar mikróbov. Veľkosť baktérií sa pohybuje od desatín do niekoľkých mikrónov. V priemere sa priemer tela väčšiny baktérií pohybuje v rozmedzí 0,5 - 1 mikrón a priemerná dĺžka pre tyčinkovité baktérie je 1 - 5 mikrónov. Rozlíšenie moderných bakteriologických mikroskopov je 0,2 mikrónu. Preto, aby ste videli ultramikroby (vírusy, bakteriofágy), musíte použiť elektrónový mikroskop, ktorý dokáže zväčšiť objem rádovo milióny a má rozlíšenie 0,4 mm. . [...]
Jednotlivé vírusy jedného alebo druhého druhu sú formáciami rôznych tvarov (okrúhle, tyčkovité alebo iných tvarov), vo vnútri ktorých je nukleová kyselina (DNA alebo RNA) uzavretá v proteínovom obale (kapside). [...]
AT všeobecný pohľad vírusy sú submikroskopické formácie pozostávajúce z bielkovín a nukleových kyselín organizované vo forme vírusových častíc, ktoré sa často nazývajú vírusové telieska, virióny, virospóry alebo nukleokapsidy. [...]
Na niektorých elektrónových mikrofotografiách pozorovali diskovité častice takmer rovnakého priemeru ako intaktný vírus. V týchto časticiach bol viditeľný centrálny kanál, ktorého priemer sa menil a bol obklopený 10 radiálne umiestnenými podjednotkami. [...]
Filtrovateľné formy baktérií sa líšia od filtrovateľných vírusov tým, že sa môžu vyvíjať aj na umelých živných médiách. [...]
Baktérie obsahujú 1 - 4% tukov, 8 - 14% bielkovín a 80 - 85% vody. Stopové množstvá obsahujú fosfor, draslík, vápnik, horčík, železo a ďalšie prvky. Vírusy nemajú bunkovú štruktúru a majú veľkosť 10 - 100 nm. [...]
Pôvodcom choroby je vírus žltej mozaiky Beap (vírus Phaseolus 2 Smith). Vírus sa inaktivuje pri teplote 70 ° C. Ovplyvňuje všetky strukoviny, nemôže sa prenášať semenami. [...]
Pôvodcami sú vírus ara-bis mozaika a malinový ringspol vírus. Oba vírusy patria do rovnakej skupiny, majú izometrické častice s priemerom asi: Yu im. Prenášajú sa kontaktnými mechanickými prostriedkami, pôdnymi nematódami a štepením. Na listoch sú svetlozelené alebo žltkasté škvrny, rôznych veľkostí a tvarov s nevýraznými okrajmi. Listy sú malé, zdeformované, rastliny sú depresívne. V prípade vážneho poškodenia rastliny náchylných odrôd do roka zomrú. [...]
NOLÓGIA pre extrakciu baktérií, vírusov a chemických znečisťujúcich látok z vody ", ktorá spočíva v tom, že mikroorganizmy prechádzajúce cez celulózový sorbent" sa vďaka elektrostatickej interakcii "lepia" na štruktúru sorbentu. " Výsledkom je, že „voda je 100% dezinfikovaná pred vírusmi, 100% takmer od všetkých baktérií a 95–100% od baktérií E. coli. Nečistoty sa z vody odstraňujú komplexným spôsobom: k tomu dochádza v dôsledku mechanického zadržiavania častíc v pórovitej štruktúre filtračného materiálu v dôsledku molekulárnej sorpcie, elektrostatickej interakcie a výmeny iónov. ““ Ja osobne v tejto technológii nevidím nič PRINCÍPNE NOVÉ, ale v Zdroji poskytujúcom život je jeden originálny okamih. Citujem: „Tvar hornej časti filtra v podobe kostolnej kupoly má priaznivý energetický a psychologický vplyv na ľudí, ktorí pijú vyčistenú vodu.“ Potom nasleduje tabuľka, ktorá porovnáva výraz „Životodarný“ so všetkými druhmi „aquaforov“ a „instapurov“ (sic), pomocou ktorých si samozrejme utiera nos. [...]
Bakteriofágy a filtrovateľné vírusy nemajú obvyklú bunkovú štruktúru, preto organizovaná bunka nie je poslednou jednotkou života. To potvrdzujú fakty o prechode viditeľných foriem baktérií na „neviditeľné“, nebunkové formy, nazývané filtrovateľné formy viditeľných baktérií. [...]
Lucerna mozaika. Pôvodcom je vírus mozaiky lucerny (vírus mozaiky Alfalfa, vírus Medicago 2 Smith). Prenáša sa kontaktno-mechanickými prostriedkami, voškami, semenami. Príznaky: najskôr sa na listoch objavia malé zaoblené žltkasté škvrny, potom podlhovasté alebo nepravidelné škvrny medzi bočnými žilami, svetložlté alebo belavé čiary pozdĺž žíl. Listy sú malé a zdeformované. V lete sú príznaky často maskované. Latentná infekcia nie je nezvyčajná. Vírus má široké spektrum hostiteľov: infikuje divoké a kultivované rastliny mnohých čeľadí: móly, Solanaceae, Asteraceae, tekvice atď. [...]
Okrem organizmov s bunkovou štruktúrou existujú aj nebunkové formy života - vírusy a bakteriofágy. Mimochodom, vírusy objavil v roku 1892 ruský biológ D.I. Ivanov a ich meno v preklade znamená „jed“, čo vo všeobecnosti v každodennom živote mnohých ľudí odráža ich vplyv na zdravie. [...]
Neexistuje jasná hranica medzi živými a neživými látkami, čo potvrdzuje existencia vírusov. Posledné menované majú vlastnosti živé aj neživé. Všeobecne prijímaná definícia pre ne ešte nebola formulovaná. Spravidla sa verí, že vírusy sú najmenej organizované formy života, ktoré nemajú svoj vlastný metabolizmus a môžu existovať iba vo vnútri buniek iných organizmov. Nerozmnožujú sa mimo buniek. Schopnosť vírusov reprodukovať sa, aj keď je v kontakte s inými bunkami, je súčasne znakom živých vecí. [...]
Pôda obsahuje rôzne mikroorganizmy: baktérie, aktinomycety alebo žiarivé huby, huby, vírusy atď. Väčšina z nich spracováva podstielku (vrstvu humusu), zlepšuje štruktúru pôdy a premieňa organické zlúčeniny na stráviteľné formy. So zvyšovaním kyslosti pôdy a tvorbou rozpustných foriem toxických kovov klesá aktivita mikroorganizmov, najmä pri spracovaní lesného odpadu. [...]
Inaktivačný účinok produktov elektrolýzy a chlóru je do značnej miery ovplyvnený množstvom a formou zvyškového chlóru (voľného alebo viazaného). Štúdia dynamiky inaktivácie produktmi elektrolýzy a chlóru modelového vírusu poliomyelitídy, fágov Escherichia coli a Escherichia coli, ukázala, že v prítomnosti zvyškového chlóru iba vo viazanom stave, po 30 minútach kontaktu, Escherichia coli úplne zomrela, a vírus fága - iba o 80, respektíve 60%. So stopami voľného zvyškového chlóru boli do 20 minút kontaktu E. coli a fág inaktivované o viac ako 99% a vírus bol deaktivovaný iba o 90%. Keď je obsah voľného zvyškového chlóru vo vode 0,1-0,3 mg / l, po 10 minútach kontaktu bol úplne dezinfikovaný vo vzťahu k E. coli a fágom a po 30 minútach bolo zistené iba zanedbateľné množstvo aktívnych vírusov. Rozdiel medzi stupňom inaktivácie študovaných mikroorganizmov bol štatisticky významný vo všetkých prípadoch. Za testovaných podmienok dezinfekcie produktmi elektrolýzy a chlórom vody obsahujúcej mikroorganizmy v rovnakých koncentráciách bola Escherichia coli menej odolná ako fág a fág bol menej odolný ako vírus. Preto môžu E. coli a fágy slúžiť ako spoľahlivé hygienické ukazovatele účinnej dezinfekcie vody produktmi elektrolýzy a chlórom proti enterovírusom. V zásade sa to týka tých prípadov, keď v dôsledku nepriaznivých epidemických hygienických podmienok môže koncentrácia enterovírusov vo vode nádrží významne stúpať a dosahovať hladinu E. coli (E. L. Lovtsevich, L. A. Sergunina, 1968). [...]
Po vynáleze antibiotík teda hlavným nepriateľom človeka neboli najjednoduchšie huby a jednobunkové bunky, ale vírusy. Existujú prvé príznaky, ktoré retrovírusy prichádzajú nahradiť vírusy - predvírusové, starodávnejšie formy života, budujúce svoju organizáciu nie na základe molekuly DNA, ale na základe RNA. Jedným z najslávnejších predstaviteľov tejto formy života je AIDS retrovírus. [...]
Mikroorganizmy, ktoré sú pod mikroskopom neviditeľné, sa nazývajú ultramikróby. Z tejto skupiny ultramikroskopických foriem sú v ľudskej praxi najdôležitejšie bakteriofágy - filtrovateľné vírusy a neviditeľné formy baktérií. Ultramikroby bolo možné pozorovať iba v elektrónovom mikroskope, čo dáva nárast až 45 000-krát. Vírusy (obr. 85) sú častice zložené z proteínov a nukleovej kyseliny (DNA alebo RNA). Nemajú obvyklú bunkovú štruktúru. Medzi nebunkovú formu života patria aj bakteriofágy (obr. 86), čo sú podlhovasté útvary so zahusteným koncom. [...]
Infekčný proces je komplex reakcií v makroorganizme, ktoré vznikajú v reakcii na zavedenie a množenie mikróbov, vírusov atď. Nie vždy je sprevádzaný prítomnosťou príznakov ochorenia. Napríklad pri mikrobearálnej alebo asymptomatickej infekcii neexistujú žiadne klinické príznaky, hoci jeho patogén je prítomný v tele a ovplyvňuje jeho rôzne systémy, čo spôsobuje ich imunologickú reštrukturalizáciu. Ak je infekčný proces sprevádzaný prejavom klinických príznakov, potom sa táto forma infekcie nazýva infekčné ochorenie. Infekčné ochorenie je teda takzvanou zjavnou formou infekcie. [...]
Tieto vírusy sa vyznačujú podobnosťou morfologických charakteristík, reakciami na elektromagnetické žiarenie, reprodukciou atď. Ich hlavnými zložkami sú: C, H, N, P, O, uhľohydráty a lipidy. Je známe, že Prak-Pries a všetky onkovírusy sú tepelne nestabilné a ničia sa pri teplotách od 50 do 70 ° C, v závislosti od typu onkovírusu. [...]
Táto skupina zahŕňa koloidné (minerálne a organominerálne) častice pôd a usadenín, ako aj nedisociované a nerozpustné formy humínových látok, ktoré dodávajú vode farbu. Posledne menované sú vyplavované do prírodných vodných plôch z lesných, močaristých a rašelinových pôd a tiež sa tvoria v samotných vodných útvaroch v dôsledku životne dôležitej činnosti vodných rastlín a rias. Táto skupina môže tiež zahŕňať vírusy a iné organizmy, ktorých veľkosť sa blíži ku koloidným časticiam. Pretože medzi nimi existujú aj patogénne (patogénne) organizmy, je ich odstránenie z vody veľmi zodpovednou udalosťou. [...]
Druhá skupina nečistôt kombinuje hydrofilné a hydrofóbne minerálne a organominerálne koloidné častice pôd a usadenín, nedisociované a nerozpustné formy humínových látok s vysokou molekulovou hmotnosťou a detergenty. Kinetická stabilita hydrofóbnych nečistôt je charakterizovaná pomerom síl gravitačného poľa a Brownovho pohybu; ich agregovaná stabilita je spôsobená elektrostatickým stavom rozhrania a tvorbou difúznych vrstiev alebo vytváraním stabilizačných vrstiev na povrchu častíc. Do tejto skupiny patria aj vírusy a iné mikroorganizmy, ktoré sú svojou veľkosťou podobné koloidným časticiam. [...]
Na izoláciu a kvantifikáciu rastlinných vírusov je možné použiť Brackovu metódu centrifugácie s hustotným gradientom. Ako sa ukázalo, táto metóda má veľa možností a v súčasnosti sa široko používa v oblasti virológie a molekulárnej biológie. Pri vykonávaní štúdií centrifugáciou v gradiente hustoty je centrifugačná trubica čiastočne naplnená roztokom, ktorého hustota klesá v smere od dna k menisku. Sacharóza sa najčastejšie používa na vytvorenie gradientu pri frakcionácii rastlinných vírusov. Pred začiatkom centrifugácie môžu byť vírusové častice distribuované po celom objeme roztoku alebo nanesené na vrchol gradientu. Bracke navrhol tri rôzne spôsoby centrifugácie s hustotným gradientom. Pri izopycpickej (rovnovážnej) centrifugácii proces pokračuje, kým všetky častice v gradiente nedosiahnu úroveň, keď sa hustota média rovná ich vlastnej hustote. K frakcionácii častíc teda v tomto prípade dochádza v súlade s rozdielmi v ich hustote. Roztoky sacharózy nie sú dostatočne husté na izopynikovú separáciu mnohých vírusov. Pri vysokorýchlostnej zónovej centrifugácii sa vírus najskôr aplikuje na predtým vytvorený gradient. Častice každého typu sa sedimentujú súčasne gradientom vo forme zóny alebo pásu, ktorého rýchlosť závisí od ich veľkosti, tvaru a hustoty. V tomto prípade je centrifugácia ukončená, keď častice pokračujú v sedimentácii. Rovnovážna zonálna centrifugácia je podobná ako pri vysokorýchlostnej zonálnej centrifugácii, avšak v tomto prípade centrifugácia pokračuje, kým sa nedosiahne izopynikový stav. Úlohou gradientu hustoty pri vysokorýchlostnej centrifugácii je inhibovať konvekciu a fixovať rôzne druhy molekúl v špecifických zónach. Teória odstreďovania s gradientom hustoty je zložitá a nie je dobre pochopená. V praxi však ide o jednoduchú a elegantnú metódu, ktorá sa široko používa pri práci s rastlinnými vírusmi. [...]
Hlavnou črtou AHC CEC lokalizovanej v matrici (ako aj onkovírusov v bunke) je prítomnosť rozhrania medzi dvoma médiami s rôznou vodivosťou. Na obr. 2.11 ukazuje údaje z elektrónovej mikroskopie ukazujúce adenovírus, vírus Epstein-Barr (EBV) a ЦЭЧ v ShchGK. Obr. 2.11 je zrejmé, že všetky formácie rovnakej mierky majú tvar guľovitého tvaru, pozostávajúci z jadra a plášťa, v chemické zloženie každá škrupina obsahuje elektricky aktívne ióny, s maticami zreteľne vyznačené hranice pre vírusy a CEC. [...]
Na listoch sa objavuje skoro na jar ako žltá škvrna. Do polovice leta tento príznak zmizne, ale postihnuté listy sa niekedy pokrčia. Plody sú malé, často nepravidelné a s tuberkulózami pozdĺž švu. Ich dozrievanie sa oneskoruje. Pôvodca choroby - vírus broskyňovej mozaiky sa prenáša očkovaním a pučaním. Verí sa, že vírus nesie slivka. [...]
Napriek mnohým objavom je na obrázku biogenézy stále veľa prázdnych miest. Iba hlavné míľniky možno považovať za nespochybniteľné. Teraz teda nie je pochýb o tom, že vznik biosféry bol výnimočnou jedinou udalosťou. Drobný vírus a obrovská morská príšera, jednobunkové riasy a stromová papraď, ktoré zmizli pred miliónmi rokov, sú všetko iba konáre a listy na rovnakom fylogenetickom strome. Formy života vždy a všade odhaľujú takpovediac „pokrvný vzťah“ a všetky jej deti sú geneticky spojené. Odo dňa, keď sa na Zemi objavilo prvé stvorenie, život pochádza iba zo života. [...]
Bunka je základnou štrukturálnou a funkčnou jednotkou všetkých živých organizmov, elementárnym živým systémom. Môže existovať ako samostatný organizmus (baktérie, prvoky, niektoré riasy a huby) a ako súčasť tkanív mnohobunkových organizmov. Iba vírusy sú nebunkové formy života. [...]
Podľa navrhovanej schémy v prvej etape procesu nastáva tvorba komplexu enzýmu a substrátu EI reštrikčnej endonukleázy EcoR I a dvojvláknovej plazmidovej DNA. Kľúčovým bodom schémy je tvorba komplexu E-II reštrikčnej endonukleázy EcoR I s kruhovou formou DNA, ktorá obsahuje jednovláknový zlom, získaný v dôsledku hydrolýzy fosfodiesterovej väzby v jednom z reťazcov DNA. Ďalej v závislosti na podmienkach (povaha substrátu, teplota atď.) ) buď k štiepeniu druhého reťazca DNA v rovnakom komplexe E - II môže dôjsť vytvorením komplexu enzýmu E-III s lineárnou formou DNA, alebo k disociácii komplexu E-P vytvorením voľného enzýmu a kruhovej DNA obsahujúcej jednovláknový zlom, čo vedie k akumulácia formy II v roztoku. Táto schéma umožnila vysvetliť rozdiely v mechanizmoch hydrolýzy DNA vírusu SV 40 na jednej strane a DNA ColE I a bakteriofágu G4 na druhej strane. V prípade DNA vírusu SV 40 dochádza k disociácii komplexu enzýmu a substrátu E-11, čo vedie k akumulácii kruhovej formy DNA v roztoku. Navrhlo sa, že rozdiely v mechanizme hydrolýzy týchto molekúl DNA (vírus SV 40; DNA ColE I a bakteriofág G4) sú výsledkom interakcie reštrikčného enzýmu EcoR I s rôznymi nukleotidovými sekvenciami lemujúcimi rozpoznávacie miesto reštrikčného enzýmu EcoR I. Tento predpoklad však neumožňuje vysvetliť rozdiely. v mechanizme hydrolýzy cirkulárnej DNA ColE I v závislosti od teploty (pozri vyššie). [...]
Toto ochorenie je známe v mnohých krajinách sveta. V ZSSR sa nachádzal na Ukrajine, v Moldavsku, Estónsku a Gruzínsku a podlieha vnútornej karanténe. Ovplyvnené sú slivka, čerešňová slivka, mirabelky, marhule a broskyňa. Pôvodcom choroby je slivka kiahní (\u003d vírus Prunus 7 Smith). Forma vírusu je vláknová, veľkosť 760X20 pomenovaná po [...]
Ďalší vývoj výskumu štúdia mechanizmu hydrolýzy plazmidovej DNA reštrikčnými endonukleázami prijatý v prácach Halford et al. ... Reakčný mechanizmus je teda podobný ako mechanizmus navrhovaný na hydrolýzu DNA vírusu EÚ 40 reštrikčným enzýmom EcoI I. [...]
Okrem vyššie uvedených „všeobecných“ funkcií, prítomnosti homeostázy tela, existuje ešte jedna veľmi dôležitá vlastnosť: živá hmota akoby vytvárala ďalší biotop, a to možnosť osídlenia tela inými živými bytosťami na trvalé alebo dočasné bývanie. Je to nový biotický biotop, ktorý vytvoril život. Mnoho odborníkov považuje vírusy za tvory, ktoré obývajú toto prostredie. Takže I.A. Shilov (2000) sa domnieva, že mimoriadna jednoduchosť ich dizajnu je sekundárnym javom, ešte pravdepodobnejšie je to novoobjavená forma živých bytostí, ktorá úplne ovládla intracelulárne prostredie v organizmoch iných úrovní. Druhým potvrdením tejto práce je, že vírusy majú vysoký stupeň zložitosti a rozmanitosti genetického systému. Zjednodušenie štruktúry, ktoré bolo možné vďaka povinnému bezpodmienečnému spojeniu vírusov s hostiteľským organizmom, poskytujúcim stabilné životné podmienky, ovplyvnilo dokonca aj základné vlastnosti obsiahnuté v drvivej väčšine foriem života: vírusy nemajú podráždenosť a chýba im vlastný aparát na syntézu bielkovín. Vírusy nie sú schopné samostatnej existencie a ich spojenie s bunkou nie je len priestorové, ale aj pevné funkčné spojenie, s ktorým predstavuje bunka a vírus akúsi jednotu. [...]
Krátkodobé alkalické ošetrenie HPLC pri 30 ° C a vysokej iónovej sile in situ spôsobuje praskliny, ktoré vedú k tvorbe fragmentov RNA, ktoré majú pomerne jednotnú veľkosť, v ktorých s2 [...]
Počet ľudí postihnutých maláriou, hepatitídou, HIV a mnohými ďalšími chorobami je obrovský. Mnoho lekárov je presvedčených, že by sa nemalo hovoriť o „víťazstve“, ale iba o dočasnom úspechu v boji proti týmto chorobám. História boja infekčné choroby je veľmi krátka a nepredvídateľnosť zmien v prostredí (najmä v mestách) môže tieto zisky vyvrátiť. Z tohto dôvodu sa „návrat“ infekčných agensov zaznamenáva medzi vírusmi. Mnoho vírusov sa „odtrhne“ od prírodnej bázy a prechádza do novej etapy, ktorá môže žiť v ľudskom prostredí - stávajú sa pôvodcami chrípky, vírusových nádorov a iných chorôb. Možno je to forma HIV. [...]
Zmena hmotnostného priemeru molekulovej hmotnosti a polomeru rotácie sa posudzovala na základe údajov o rozptyle svetla. Pre HPMC RNA pomocou obidvoch degradačných metód zistili, že polomer rotácie sa zvýšil pred nástupom intenzívnej degradácie molekuly, zatiaľ čo polomer rotácie a hmotnostná priemerná molekulová hmotnosť TMV RNA sa znížili od samého začiatku tohto procesu. Strazielli a kol. Toto zistenie vysvetlil tým, že naznačuje, že RNA z HPMC existuje vo forme uzavretej slučky. Tieto výsledky je však možné interpretovať iným spôsobom. Napríklad Hazelkorn ukázal, že TMV RNA a HPMT RNA ko-sedimentujú za podmienok pH a iónovej sily podobných tým, ktoré používajú Strazielli a kol. Naopak, kruhové a lichgeous formy DNA fága cpX174 sú ľahko rozlíšiteľné podľa ich sedimentačných vlastností [1 515]. Kuiper na základe údajov o sedimentácii za rôznych podmienok naznačil, že izolovaná RNA vírusu mozaiky uhoriek (kmeň Y) môže existovať v dvoch formách: otvorený reťazec a kruhová štruktúra. Tieto údaje, rovnako ako v prípade opísanom vyššie, je však možné vysvetliť rôznymi spôsobmi. [...]
Bakteriálna DNA je vysoko polymérna zlúčenina pozostávajúca z veľkého množstva nukleotidov - polynukleotidov s molekulovou hmotnosťou asi 4 milióny.Molekula DNA je reťazec nukleotidov, kde ich umiestnenie má špecifickú sekvenciu. V poradí usporiadania dusíkatých báz je zakódovaná genetická informácia každého druhu. Porušenie tejto sekvencie je možné pri prírodných mutáciách alebo pod vplyvom mutagénnych faktorov. V tomto prípade mikroorganizmus nadobúda alebo stráca akýkoľvek majetok. Zdedil zmeny vlastností, to znamená, že sa objaví nová forma mikroorganizmu. Vo všetkých mikroorganizmoch - prokaryotoch a eukaryotoch - sú nosičmi genetickej informácie nukleové kyseliny - DNA a RNA. Iba niekoľko vírusov je výnimkou: nemajú DNA a dedičné informácie sa zaznamenávajú alebo odrážajú iba v RNA.