Вірусні захворювання виникли в далекій давнині, однак вірусологія як наука почала розвиватися в кінці XIX століття.
У 1892 р російський вчений-ботанік Д. І. Іванівський, вивчаючи мозаїчну хворобу листя тютюну, встановив, що захворювання це викликається дрібними мікроорганізмами, які проходять через дрібнопористі бактеріальні фільтри. Ці мікроорганізми отримали назву фільтруються вірусів (від лат. Virus - отрута). Надалі було показано, що є й інші мікроорганізми, що проходять через бактеріальні фільтри, тому що фільтруються віруси стали називати просто вірусами.
В цілому, форма віріона і наявність або відсутність конверта мало говорять про те, які захворювання можуть викликати віруси або які види вони можуть заразити, але все ж є корисним засобом для початку вірусної класифікації. Малюнок 5 Віруси можуть бути складними за формою або відносно простими.
Яке з наступних тверджень про структуру вірусу вірно? Ядро вірусу містить геном або загальне генетичне зміст вірусу. Вірусні геноми, як правило, малі в порівнянні з бактеріями або еукаріот, що містять тільки ті гени, які кодують білки, які вірус не може отримати з клітини-хазяїна. Цей генетичний матеріал може бути одноланцюговим або двухцепочечную. Він також може бути лінійним або круговим. Хоча більшість вірусів містять один сегмент нуклеїнової кислоти, у інших є геноми, які складаються з декількох сегментів.
Великий внесок у вивчення вірусів внесли радянські вірусологи: М. А. Морозов, Н. Ф. Гамалія, Л. А. Зільбер, М. П. Чумаков, А. А. Смородинцев, В. М. Жданов та ін.
Віруси - це некліткова форма існування живої матерії. Вони дуже малі. За образним висловом В. М. Жданова "величину їх по відношенню до величини середніх бактерій можна порівняти з величиною миші по відношенню до слону". Побачити віруси стало можливим тільки після винаходу електронного мікроскопа.
Віруси можна розглядати як облігатні внутрішньоклітинні паразити. Вірус повинен прикріплюватися до живій клітині, приймати всередину, виробляти його білки і копіювати його геном, а також шукати спосіб уникнути клітини, щоб вірус міг заразити інші клітини і, в кінцевому рахунку, інших людей. Віруси можуть заражати тільки певні види хостів і тільки певні клітини всередині цього хоста. Молекулярна основа цієї специфічності полягає в тому, що певну поверхневу молекулу, відому як вірусний рецептор, необхідно знайти на поверхні клітини-господаря для прикріплення вірусу.
В даний час для вивчення вірусів використовують багато методів: хімічні, фізичні, молекулярно-біологічні, імунобіологічні і генетичні.
Всі віруси підрозділяються на вражають людину, тварин, комах, бактерії і рослини.
У вірусів спостерігається велика різноманітність форм і біологічних властивостей, проте всі вони мають загальні риси будови. Зрілі частинки вірусів називають вирионами.
Крім того, метаболічні відмінності, які спостерігаються в різних типах клітин, засновані на диференціальної експресії генів, є ймовірним фактором, в якому клітини можуть використовувати вірус для реплікації. Клітка повинна становити речовини, необхідні вірусу, такі як ферменти, в яких сам вірусний геном не містить генів, або вірус не зможе реплицироваться з використанням цієї клітини.
Вірус повинен «захопити» осередок для реплікації. Цикл вірусної реплікації може призводити до різких біохімічним і структурним змінам в клітці-хазяїні, що може викликати пошкодження клітин. Ці зміни, звані цитопатичної ефект, можуть змінювати функції клітин або навіть руйнувати клітку. Деякі інфіковані клітини, такі як інфіковані простудних вірусом, вмирають шляхом лізису або апоптозу, одночасно вивільняючи все віріони потомства. симптоми вірусних захворювань є результатом імунної відповіді на вірус, який намагається контролювати і усувати вірус з організму, а також від пошкодження клітин, викликаного вірусом.
На відміну від інших мікроорганізмів, що містять одночасно ДНК і РНК, віріон містить тільки одну з нуклеїнових кислот - або ДНК, або РНК.
Нуклеїнова кислота вірусів може бути однонитчатим і двунитчатой. Майже всі віруси, що містять РНК, мають в своєму геномі однонитчатим РНК, а містять ДНК - двунитчатую ДНК. Відповідно до двома типами генетичного матеріалу віруси поділяють на РНК і ДНК-містять. До ДНК-відносяться 5 сімейств, РНК-- 10 сімейств.
Під час процесу брунькування клітина не піддається лізису і не відразу побивається. Більшість продуктивних вірусних інфекцій слідують аналогічних кроків в циклі реплікації вірусу: вкладення, проникнення, непокриті, реплікація, збірка і випуск. Вірус прикріплюється до певного рецепторному сайту на мембрані клітини-господаря через кріпильні білки в капсиді або білках, вбудованих в його оболонку. Вкладення специфічно, і зазвичай вірус буде прикріплюватися тільки до клітин одного або декількох видів і тільки певним типам клітин в межах цих видів з відповідними рецепторами.
* (Тут наведені дані, що стосуються тільки деяких з патогенних для людини вірусів.)
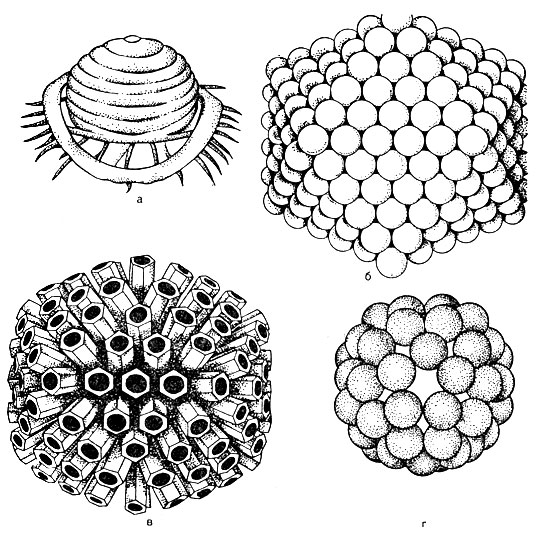
структура віріона. У центрі віріона знаходиться нуклеїнова кислота, яка оточена капсидом (від грец. Kanca - ящик). Капсид складається з білкових субодиниць, званих капсомерами. Зрілий вірус за хімічною структурою є нуклеокапсидом. Кількість капсомер і спосіб їх укладання (рис. 52) строго постійні для кожного виду вірусу. Наприклад, вірус поліомієліту містить 32 капсомеров, а аденовірус - 252 капсомеров. Капсомери можуть бути покладені в вигляді багатогранника з рівномірними симетричними гранями - кубоідальная форма (наприклад, аденовірус). Укладання у вигляді спіралей (сферична) характерна для вірусів грипу. Може бути тип симетрії, при якому нуклеїнова кислота має вид пружини, навколо якої покладені капсомери, в цьому випадку вірус має паличкоподібну форму - вірус, що викликає хворобу листя тютюну.
На відміну від тварин вірусів, нуклеїнова кислота бактеріофагів вводиться в клітину-хазяїн голим, залишаючи капсид поза клітиною. Віруси рослин і тварин можуть проникати в клітини через ендоцитоз, в якому клітинна мембрана оточує і поглинає весь вірус. Деякі оповиті віруси потрапляють в клітину, коли вірусна оболонка зливається безпосередньо з клітинної мембраною. Після всередині клітини вірусний капсид розкладається і вивільняється вірусна нуклеїнова кислота, яка потім стає доступною для реплікації і транскрипції.
Механізм реплікації залежить від вірусного генома. Звичайно, є винятки з цього шаблону. Якщо клітина-господар не забезпечує ферменти, необхідні для реплікації вірусу, вірусні гени надають інформацію для безпосереднього синтезу відсутніх білків.
Складний тип симетрії має фаг: головка - кубоідальной, а відросток - паличкоподібні форми (сперматозоідная форма) (див. Рис. 21, 22).
Таким чином, в залежності від способу укладання віруси поділяють на кубоідальную, сферичну, паличкоподібну і сперматозоідную форми.
Останнім етапом реплікації вірусу є вивільнення нових віріонів в організм господаря, де вони здатні інфікувати сусідні клітини і повторювати цикл реплікації. Деякі віруси вивільняються, коли клітина-господар помирає, а інші віруси можуть залишати заражені клітини, просочуючись через мембрану, не вбиваючи безпосередньо клітку.
Малюнок 6 При зараженні вірусом грипу глікопротеїни приєднуються до епітеліальної клітці господаря. В результаті вірус поглинається. Малюнок 7 Віруси є причиною безлічі захворювань у людей, починаючи від легких хвороб і закінчуючи серйозними захворюваннями.
Деякі віруси, що володіють більш складною структурою, мають оболонку, яка називається пеплос. Вона утворюється при виході вірусу з клітини господаря. Вірусний капсид при цьому обволікається внутрішньою поверхнею мембрани цитоплазми клітини господаря і утворюється один або кілька шарів оболонки суперкапсид. Таку оболонку мають тільки деякі віруси, наприклад віруси сказу, герпесу, енцефаліту. Ця оболонка містить фосфоліпіди, що руйнуються під впливом ефіру. Таким чином, впливаючи ефіром, можна відрізнити вірус, що має пеплос, від вірусу з "голим капсидом".
Вакцина може бути приготовлена \u200b\u200bз використанням ослаблених живих вірусів, убитих вірусів або молекулярних субодиниць вірусу. Загалом, живі віруси призводять до підвищення імунітету, але мають можливість викликати захворювання на деякій низькій частоті. Убита вірусна вакцина і суб'едінічние віруси здатні викликати хворобу, але в цілому призводять до менш ефективного або довготривалого імунітету.
Ослаблені живі вірусні вакцини розроблені в лабораторії, щоб викликати мало симптомів у реципієнтів, одночасно надаючи їм імунітет проти майбутніх інфекцій. Поліомієліт був однією з хвороб, яка стала важливою віхою в використанні вакцин. Успіх вакцини проти поліомієліту проклав шлях до рутинного розподілу дитячих вакцин проти кору, епідемічного паротиту, краснухи, вітрянки та інших захворювань. Живі вакцини зазвичай виробляють за рахунок ослаблення вірусу «дикого типу» шляхом вирощування його в лабораторії в тканинах або при температурах, відмінних від того, до чого вірус звик в приймаючій.
У деяких вірусів з зовнішнього ліпідного шару оболонки виступають капсомери у вигляді шипів (ці шипи тупі). Такі віруси називаються пепломерамі (наприклад, вірус грипу, див. Рис. 52).
Нуклеїнова кислота вірусу є носієм спадкових властивостей, а капсид і зовнішня оболонка несуть захисні функції, як би оберігаючи нуклеїнових кислот. Крім того, вони сприяють проникненню вірусу в клітину.
Наприклад, вірус можна вирощувати в клітинах в пробірці, ембріонах птахів або живих тварин. Адаптація до цих нових клітин або температурі викликає мутації в геномах вірусу, що дозволяє їм краще рости в лабораторії, одночасно перешкоджаючи їх здатності викликати захворювання при повторному введенні в стан, виявлене у господаря. Ці аттенуіровані віруси, таким чином, все ж викликають інфекцію, але вони не ростуть дуже добре, що дозволяє імунних реакцій розвиватися в часі, щоб запобігти серйозне захворювання.
Небезпека використання живих вакцин, які, як правило, більш ефективні, ніж убиті вакцини, - це низький, але зі значним ризиком того, що ці віруси повернуться до своєї хвороботворної формі шляхом зворотних мутацій. Зворотні мутації виникають, коли вакцина піддається мутаціям в господаря, так що вона зчитується господареві і може знову викликати захворювання, яке потім може бути поширене на інших людей в епідемії.
розміри вірусів. Вимірюються віруси в нанометрах. Величина їх коливається в широкому діапазоні від 15-20 до 350-400 нм.
Методи вимірювання вірусів: 1) фільтрування через бактеріальні фільтри з відомою величиною пір; 2) ультрацентрифугирование - великі віруси осідають швидше; 3) фотографування вірусів в електронному мікроскопі.
При грипі мутація в генах для поверхневих молекул допомагає вірусу уникнути захисного імунітету, який міг бути отриманий в попередній сезон грипу, що робить необхідним, щоб люди отримували вакцинацію щороку. Інші віруси, такі як ті, які викликають хвороби дитинства, кору, епідемічного паротиту та краснухи, мутують так мало, що одна і та ж вакцина використовується рік за роком.
Вакцини та противірусні препарати для лікування
У деяких випадках вакцини можуть використовуватися для лікування активної вірусної інфекції. У разі сказу смертельне неврологічне захворювання, що передається в слині вірусів, заражених сказом, прогресування захворювання з моменту укусу тварин до моменту його надходження в центральну нервову систему може становити два тижні або довше. Це досить часу для вакцинації людини, який підозрюється, що його вкусив скажена тварина, і підсиленої імунної відповіді від вакцинації достатньо, щоб запобігти проникненню вірусу в нервову тканину.
Хімічний склад вірусів. Кількість і зміст ДНК і РНК вірусів неоднакові. У ДНК молекулярна маса коливається від 1 · 10 6 до 1,6 × 10 8, а у РНК - від 2 · 10 6 до 9,0 · 10 6.
Білки у віріонів виявлені в незначній кількості, вони складаються з 16-20 амінокислот. Крім капсидних білків, є ще внутрішні білки, пов'язані з нуклеїнової кислотою. Білки обумовлюють антигенні властивості вірусів, а також в силу щільного укладання поліпептидних ланцюгів захищають вірус від дії ферментів клітини господаря.
Таким чином, смертельні неврологічні наслідки хвороби запобігають, і індивідуум повинен тільки оговтатися від зараженого укусу. Цей підхід також використовується для лікування Ебола, одного з найшвидших і смертоносних вірусів, що вражають людей, хоча зазвичай заражає обмежені популяції. Ебола також є головною причиною смерті в горил. Використовуючи недавно розроблені вакцини, які підсилюють імунну відповідь, є надія, що імунна система постраждалих людей буде краще контролювати вірус, що потенційно знижує рівень смертності.
Ліпіди і вуглеводи виявлені у зовнішній оболонці складних віріонів. Джерелом ліпідів і вуглеводів є оболонка клітини господаря. Полісахариди, які входять до складу деяких вірусів, обумовлюють здатність їх викликати аглютинацію еритроцитів.
ферменти вірусів. Віруси не мають власного метаболізму, тому вони не потребують ферментах обміну речовин. Однак у деяких вірусів виявлено наявність ферментів, що сприяють проникненню їх в клітку господаря. Наприклад, у вірусу грипу А виявлено нейрамінідаза, відщеплюється нейрамінової кислоту, що міститься в оболонках тварин клітин (еритроцитів і ін.). У фагів - лізоцим, який руйнує клітинну оболонку, фосфатаза та ін.
Іншим способом лікування вірусних інфекцій є використання противірусних препаратів. Ці препарати часто мають обмежену здатність вилікувати вірусне захворювання, але були використані для контролю і зменшення симптомів для широкого спектра вірусних захворювань. Для більшості вірусів ці препарати інгібують вірус, блокуючи дії одного або декількох його білків. Важливо, щоб цільові білки кодувалися вірусними генами і що ці молекули не присутні в здорової клітини-господаря.
Таким чином, вірусний зростання відзначено зниження без пошкодження господаря. існує безліч антивірусних препаратів, Доступних для лікування інфекцій, деякі специфічні для конкретного вірусу і інші, які можуть впливати на кілька вірусів. Противірусні препарати розроблені для лікування генітального герпесу і грипу. Для генітального герпесу такі препарати, як ацикловір, можуть зменшувати кількість і тривалість епізодів активного вірусного захворювання, при якому у пацієнтів розвиваються вірусні ураження в клітинах шкіри.
Виявлення вірусних антигенів. Вірусні антигени в інфікованих клітинах господаря можна виявити за допомогою методу імунофлюоресценції. Препарати, що містять клітини, інфіковані вірусами, Обробляють специфічними імунними люминесцирующими сироватками. При перегляді в люмінесцентному мікроскопі в місцях скупчення вірусних частинок спостерігається характерне світіння. Вид вірусу визначають за відповідністю специфічної люминесцирующей сироватки, що викликала світіння.
Оскільки вірус залишається прихованим в нервової тканини організму для життя, цей препарат не є ліками, але може зробити симптоми захворювання більш керованими. Для грипу такі препарати, як Таміфлю, можуть зменшити тривалість симптомів «грипу» на один або два дні, але препарат не повністю попереджає симптоми. інші противірусні препарати, Такі як рибавірин, були використані для лікування різних вірусних інфекцій.
Коли будь-який з цих препаратів використовується індивідуально, висока швидкість мутації вірусу дозволяє вірусу швидко розвивати стійкість до препарату. Віруси - це безклітинні суті, які зазвичай можна побачити тільки за допомогою електронного мікроскопа. Віруси різноманітні, заражають археї, бактерії, гриби, рослини і тварин. Віруси складаються з ядра нуклеїнової кислоти, оточеного білковим капсидом з або без зовнішньої ліпідної оболонки.
Впровадження вірусу в клітину, взаємодія його з кліткою господаря і репродукція (Розмноження) складаються з ряду послідовних стадій.
Стадія 1. Починається з процесу адсорбції за рахунок рецепторів віріона і клітини. У складних віріонів рецептори розташовуються на поверхні оболонки в вигляді шиловидних виростів (вірус грипу), у простих віріонів - на поверхні капсида.
Стадія 2. Проникнення вірусу в клітину господаря протікає по-різному у різних вірусів. Наприклад, деякі фаги протикають оболонку своїм відростком і впорскують нуклеїнових кислот в клітку господаря (див. Розділ 8). Інші віруси потрапляють в клітину шляхом втягування вірусної частинки за допомогою вакуолі, т. Е. На місці впровадження в оболонці клітини утворюється поглиблення, потім краї її сходяться і вірус виявляється в клітці. Таке втягування називається віропексіса.
Стадія 3. "Роздягання вірусу" (дезінтеграція). Для свого відтворення вірусна нуклеїнова кислота звільняється від захищають її білкових покривів (оболонки і капсида). Процес роздягання може початися під час адсорбції, а може статися тоді, коли вірус знаходиться вже всередині клітини.
Стадія 4. На цій стадії відбувається реплікація (відтворення) нуклеїнових кислот і синтез вірусних білків. Ця стадія відбувається за участю ДНК або РНК клітини господаря.
Стадія 5. Складання вириона. Цей процес забезпечується самосборке білкових частинок навколо вірусної нуклеїнової кислоти. Синтез білка може початися безпосередньо після синтезу вірусної нуклеїнової кислоти або після інтервалу в кілька хвилин або кілька годин. У одних вірусів самосборка відбувається в цитоплазмі. У інших в ядрі клітини господаря. Освіта зовнішньої оболонки (пеплоса) завжди відбувається в цитоплазмі.
Стадія 6. Вихід віріона з клітки господаря відбувається шляхом просочування вірусу через оболонку клітини або через отвір, що утворився в клітці господаря (в цьому випадку клітина господаря гине).
Типи взаємодії вірусу і клітини. Перший тип - продуктивна інфекція - характеризується утворенням нових віріонів в клітці господаря.
Другий тип - абортивна інфекція полягає в тому, що обривається реплікація нуклеїнової кислоти.
Третій тип - характеризується встраиванием вірусної нуклеїнової кислоти в ДНК клітини господаря; виникає форма співіснування вірусу і клітини господаря (вірогенія). У цьому випадку забезпечується синхронність реплікації вірусної і клітинної ДНК. У фагів це називається лізогенія.
мікроскопічне дослідження. При окремих вірусних інфекціях в цитоплазмі або ядрах клітин організму господаря спостерігаються специфічні внутрішньоклітинні тільця - включення, що мають діагностичне значення (тільця Бабеша - Негрі при сказі, тільця Гварніері при віспі і ін.). Розміри вірусних частинок і тілець-включень вдається штучно збільшити спеціальними методами обробки препаратів з протравов і імпрегнацією (наприклад, метод сріблення по Морозову) і спостерігати за иммерсионной мікроскопії. Більш дрібні віріони, що лежать за межами видимості оптичного мікроскопа, виявляються тільки при електронній мікроскопії. Існують різні точки зору щодо внутрішньоклітинних включень. Одні автори вважають, що вони являють собою скупчення вірусів. Інші вважають, що вони виникають в результаті реакції клітини на впровадження вірусів.
генетика вірусів. Модифікація (неуспадковане зміни) у вірусів обумовлюється особливостями клітини господаря, в якій відбувається репродукція вірусу. Модифіковані віруси набувають здатність заражати клітини, аналогічні тим, в яких вони модифікувалися. У різних вірусів модифікація по-різному проявляється. Наприклад, у фагів змінюється форма "негативних плям" (фагових колоній).
Мутація - у вірусів виникає під впливом тих же мутагенів, які викликають мутацію у бактерій (фізичні і хімічні чинники). Виникає мутація під час реплікації нуклеїнових кислот. Мутації зачіпають різні властивості вірусів, наприклад чутливість до температури і ін.
Генетична рекомбінація у вірусів може виникнути в результаті одночасного зараження клітини господаря двома вірусами, при цьому може відбутися обмін окремими генами між двома вірусами і утворюються рекомбінанти, що містять гени двох батьків.
Генетична реактивация генів іноді відбувається при схрещуванні инактивированного вірусу з повноцінним, що призводить до порятунку инактивированного вірусу.
Спонтанна і спрямована генетика вірусів має велике значення в розвитку інфекційного процесу.
Стійкість до факторів навколишнього середовища. Більшість вірусів інактивується при дії високих температур. Проте є винятки, наприклад вірус гепатиту термостійкий.
До низьких температур віруси не чутливі, ультрафіолетові сонячні промені надають інактивує дію на віруси. Розсіяне сонячне світло діє на них менш активно. Віруси стійкі до гліцерину, що дає можливість тривалий час зберігати їх в гліцерині. Вони стійкі до антибіотиків (при культивуванні вірусів досліджуваний матеріал обробляють антибіотиками для придушення бактеріальної флори).
Кислоти, луги, дезінфікуючі речовини інактивують віруси. Однак деякі віруси, інактивовані формаліном, зберігають імуногенні властивості, що дозволяє використовувати формалін для отримання вакцин (вакцина проти сказу).
сприйнятливість тварин. Коло сприйнятливих тварин для деяких вірусів дуже широкий, наприклад до вірусів сказу чутливі багато тварин. Деякі віруси вражають тільки один вид тварини, наприклад вірус чуми собак вражає тільки собак. Є віруси, до яких тварини не чутливі - наприклад, вірус кору і т. Д.
Органотропність вірусів. Віруси мають здатність вражати певні органи, тканини і системи. Наприклад, вірус сказу вражає нервову систему. Вірус віспи має дермотропні і т. Д.
Виділення вірусів в навколишнє середовище. З хворого організму віруси можуть виділятися з калом, наприклад вірус поліомієліту та інші ентеровіруси. Вірус сказу виділяється зі слиною, вірус грипу - з виділенням слизової носоглотки і т. Д.
Основні шляхи передачі вірусів. Повітряно-краплинний (грип, віспа), харчової (поліомієліт, гепатит А), контактно-побутовий (сказ), трансмісивний (енцефаліт).
противірусний імунітет. Організм людини володіє природженою стійкістю до деяких вірусів. Наприклад, людина не чутливий до вірусу чуми собак. Тварини не чутливі до вірусу кору. У цих випадках противірусний імунітет заснований на відсутності клітин, здатних підтримувати репродукцію вірусів.
Противірусний імунітет обумовлюється як клітинними, так і гуморальними факторами захисту, неспецифічними і специфічними. Неспецифічні фактори. Потужним інгібітором репродукції вірусів є білкова речовина - інтерферон. В здоровому організмі він міститься в незначній кількості, а віруси сприяють продукції інтерферону і кількість його значно збільшується. Він неспецифичен, так як блокує репродукцію різних вірусів. Однак він володіє тканинної специфічністю, т. Е. Клітини різних тканин утворюють неоднаковий інтерферон. Вважають, що механізм дії його полягає в тому, що він перешкоджає синтезу білка в клітині господаря і цим припиняє репродукцію вірусу.
До специфічних факторів противірусного імунітету відносяться вируснейтрализующие антитіла, гемагглютинирующие і преципитирующие.
Методи культивування вірусів. Віруси розмножуються тільки в життєздатних клітинах. Їх культивують: в курячих ембріонах (рис. 53), культурах тканини людини і різних тварин, в організмі чутливих тварин, сприйнятливих членистоногих.
У перший період розвитку вірусології основним методом вивчення вірусів було штучне зараження тварин, але цей метод складний, і крім цього тварини до багатьох вірусів виявилися несприйнятливі.
Велике значення в розвитку вірусології мало запровадження методів культивування вірусів в курячих ембріонах і в культурі клітин тканин людини і тварин.
Зараження курячих ембріонів. Для репродукції вірусів використовують курячі ембріони 7-12-денного віку, інкубовані в термостаті при 37 ° С. Необхідною умовою для правильного розвитку зародка є дотримання певної вологості повітря, яку можна створити, помістивши в термостат посудину з водою.
Придатність курячого ембріона для зараження визначається за наявністю рухів ембріона і розвиненої мережі кровоносних судин на хоріон-аллантоісной оболонці при просвічуванні за допомогою овоскопа.
Культивування вірусів в курячих ембріонах проводиться в різних місцях ембріона, який заражають (див. Рис. 53):
1) на хоріон-аллантоісную оболонку,
2) в аллантоісную порожнину;
3) в амніотичну порожнину;
4) в жовтковий мішок.
Зараження курячих ембріонів проводять в боксі з використанням стерильних інструментів. Перед зараженням курячі ембріони дворазово протирають ватним тампоном, змоченим спиртом.
Зараження на хоріон-аллантоісную оболонку. Після дезінфекції яйця обережно зрізають шматочок шкаралупи з тупого кінця, знімають подскорлупную оболонку - при цьому виявляється хоріон-аллантоісная оболонка. Інфекційний матеріал в кількості 0,1-0,2 мл за допомогою шприца або пастерівської піпетки наносять на хоріон-аллантоісную оболонку. Після зараження отвір закривають ковпачком і просвіт між ним і курячим ембріоном заливають парафіном.
На іншій стороні яйця простим олівцем пишуть назву інфекційного матеріалу і дату зараження.
Зараження в амніотичну порожнину. Яйце овоскопіруют і на бічній стороні вибирають ділянку, де хоріон-аллантоіс позбавлений великих кровоносних судин. Ця ділянка відзначають олівцем. Яйця укладають на підставку в горизонтальному положенні, дезінфікують і спеціальним стерильним списом проколюють отвір в шкаралупі на глибину 213 мм, через яку вводять на цю ж відстань голку з інфекційним матеріалом безпосередньо в амніотичну порожнину. Для того щоб вводиться рідина не витікала назад, попередньо роблять прокол над повітряним мішком, після чого обидва отвори заливають парафіном.
Зараження в аллантоісную порожнину. Зараження проводять в затемненому боксі. Відзначають повітряний простір, шкаралупу над повітряним простором дезінфікують і через отвір в шкаралупі вводять у напрямку до ембріону голку шприца з матеріалом. Якщо голка потрапила в аллантоісную порожнину, то спостерігається зміщення тіні ембріона. Після зараження отвір заливають парафіном.
Зараження в жовтковий мішок. Шкаралупу дезінфікують. Яйце поміщають на підставку тупим кінцем вправо так, щоб жовтковий мішок був повернутий догори. Над повітряною камерою в центрі проколюють отвір. Через отвір в шкаралупі в горизонтальному напрямку на глибину 2-3 мм вводять голку шприца, яка потрапляє в жовтковий мішок. Матеріал вводять в обсязі 0,2-0,3 мл. Після введення матеріалу отвір парафінують.
Температурний режим і тривалість інкубації залежать від біологічних властивостей введеного вірусу.
Інфіковані яйця щодня перевіряють - овоскопіруют для перевірки життєздатності ембріона. Якщо ембріони гинуть в першу добу, то причиною цього зазвичай буває травма при зараженні. Такі яйця виводять з досвіду.
При необхідності окремо досліджувати кожну складову частину ембріона матеріал збирають у певному порядку: відсмоктують аллантоісную рідина, потім амніотичну рідину, розрізають хоріон-аллантоісную оболонку, відокремлюють амниотическую оболонку, ембріон, жовтковий мішок і тільки після цього витягують хоріон-аллантоісную оболонку, відокремивши її від внутрішньої поверхні шкаралупи. Наявність вірусу в зараженому ембріоні визначають по характерних змін хоріон-аллантоісной оболонки зараженого курячого ембріона.
Віруси, що не володіють гемагглютинирующей активністю, виявляють за допомогою РСК.
Для виявлення вірусу в аллантоісной або амниотических рідинах заражених ембріонів ставлять РДА (гемагглютинация викликається аллантоісной або амніотичної рідинами або суспензією, приготовленої з хоріон-аллантоісной оболонки).
Культивування вірусів в культурі клітин. Для накопичення вірусів в чувстсітельних клітинних культурах використовуються тканини людини і різних тварин. Найбільше практичне застосування отримали одношарові культури первинно-трипсінізірована і перещеплюваних ліній клітин.
Одношарові культури клітин вирощують в скляних плоских судинах-матрацах. Клітинна суспензія в рідкому поживному середовищі при температурі 37 ° С дозволяє отримати "in vitro" шар клітин з певною гістологічної структурою. Присутність вірусів в культурах тканин виявляють по зміні (дегенерації) клітин. Тип вірусів визначають шляхом нейтралізації дії вірусів при додаванні до віруссодержащего матеріалу відповідних типоспецифічних сироваток.
Ці методи дозволяють швидше враховувати результати дослідження і є більш економічними. У тих випадках, коли віруси не викликають цитопатичної дії (дегенерації) і не розвиваються в курячих ембріонах, користуються методами зараження тварин (див. Розділ 11).
Для культивування вірусів використовують перещеплюваних клітини, які частіше отримують з клітин злоякісних пухлин.
Одношарові культури отримують з ембріонів людини, курки, тварин.
Перевага одношарових культур клітин - простота методики і легкість обліку.
Здатність клітин до розмноження поза організмом пов'язана зі ступенем диференціації тканини. Менш диференційовані тканини мають більшу здатність до проліферації (сполучна, епітеліальна тканина).
Сутність методів при приготуванні первинних культур тканини полягає в руйнуванні міжклітинної тканини і роз'єднанні клітин для подальшого отримання моношару.
Роз'єднання клітин проводиться шляхом впливу на тканину протеолітичних ферментів, найчастіше трипсину. Розчин трипсину сприяє роз'єднанню клітин при збереженні у них здатності до розмноження. Для вирощування культури клітин необхідна живильне середовище. Склад середовища складний, він включає цілий ряд інгредієнтів: амінокислоти, глюкозу, вітаміни, мінеральні солі, коферменти і т. Д. Отримання культури тканини проводять в строго асептичних умовах. У середу додаються антибіотики (500 ОД пеніциліну і 250 ОД стрептоміцину в 1 мл) для пригнічення росту бактеріальної флори.
Підготовлену тканину заливають 0,25% розчином підігрітого трипсину і інкубують в термостаті при 37 ° С. Під час інкубації тканину періодично помішують шляхом обертання колби. Трипсінізірована клітини центрифугують при 800-1000 об / хв протягом 5 хв.
Тріпсінізаціі і центрифугування проводять дуже обережно, щоб не травмувати клітини. Після центрифугування надосадову рідину видаляють, а осад клітин поміщають в невеликий обсяг живильного середовища. Для отримання однорідної маси суспензія клітин фільтрують через один шар марлі в воронці (стерильною). Суспензія клітин перевіряють на стерильність шляхом посіву по 0,1 мл, в 2 пробірки з цукровим бульйоном.
Успіх культивування клітин залежить від посівної Дози, тому після тріпсінізаціі проводять підрахунок клітин в камері Горяєва. Після підрахунку суспензія клітин розводять живильним середовищем з такого розрахунку, щоб в 1 мл містилося 500000-1000000 клітин і розливають по пробірках і матраців. Пробірки з культурою тканини инкубируют в термостаті в похилому положенні.
Посіяні культури щодня переглядають під малим збільшенням мікроскопа для визначення характеру їх зростання. Нормальні проліферуючі клітини світлі і ростуть одношаровим пластом. Якщо клітини темні, зернисті і не проліферують, що може бути результатом забруднення (погана обробка посуду або забруднення інгредієнтів), то такі культури вилучають з досвіду.
Зміна живильного середовища через 2-3 дня після посіву покращує інтенсивність проліферації.
Нормальні, добре проліферуючі клітини заражають досліджуваним матеріалом.
Перещеплюваних культури переважно отримують з злоякісних пухлин. Штам Hela - культура клітин раку шийки матки жінки на ім'я Helena (отриманий в 1950 р); штам Нер-2 виділено від хворого на рак гортані. Зростання цих клітин підтримується в лабораторіях шляхом послідовних пасажів. Особливість їх полягає в тому, що вони розмножуються протягом тривалого терміну. В даний час ці клітини пройшли вже тисячі генерацій. В процесі пасажів вони втрачають деякі морфологічні і біохімічні властивості - піддаються мутації. Однак залишаються цілком придатними для культивування в них вірусів. Культурою цих клітин користуються лабораторії всього світу.
Розмноження вірусу в культурі клітин відбувається в різні терміни залежно від властивостей вірусу і виду клітин.
Про наявність вірусу судять по цитопатичної дії. У мікроскопі спостерігається дегенерація клітин. Час цитопатичної дії і його характер залежать від дози і властивостей вірусу.
У деяких вірусів цитопатическое дія виявляється через кілька днів (вірус віспи), у інших - через 1-2 тижні (вірус гепатиту та ін.).
В даний час відомі вже сотні вірусів, що вражають людину. Боротьба з вірусними інфекціями здійснюється різними методами. Найбільш ефективна імунізація. Таким способом ліквідована віспа, скорочена захворюваність на поліомієліт. Важливе значення в боротьбі з вірусними інфекціями мають громадська профілактика - знищення бродячих собак (боротьба зі сказом), особиста профілактика і т. Д.
Однак ці заходи не можуть забезпечити ліквідацію всіх вірусних захворювань. Вчені наполегливо шукають шляхи, за допомогою яких можна було б вразити вірус, не пошкодивши клітку, в якій він знаходиться.
Тому закономірно, що в програмі КПРС вірусологія названа однією з провідних галузей природничо-наукових знань, яка повинна отримати переважне розвиток в найближчі роки.
Основні методи дослідження вірусів. 1. Реакція гемаглютинації, реакція затримки гемаглютинації, реакція непрямої гемаглютинації. Реакція зв'язування комплементу.
2. Реакція нейтралізації вірусів в культурі тканин.
3. Метод імунофлюоресценції.
4. Гістологічний метод - виявлення включень (тілець Бабеша - Негрі - при сказі; тілець Пашена - при віспі і ін.).
5. Біологічний метод.
Вірусна інфекція клітини являє собою комплекс про процесів, що виникають при взаємодії клітини з вірусним ге номом. Позаклітинний вирион інертний, і ця інертність зберігаючи ється до тих пір, поки вірусний геном не почне функціонувати внутрішньоклітинно, як самостійна генетична одиниця. Кар дінальним ознакою, покладеним в основу наведеної нижче класифікації, є взаємини геномів вірусу і клітини. Якщо вірусний геном реплікується незалежно від Реплі кации клітинного генома - це автономний тип вірусної инфек ції. Якщо вірусний геном включається до складу клітинного генома і реплицируется разом з ним - це інтегративний тип. Кожен з цих двох основних типів вірусної інфекції може бути під розділений на кілька форм за освітою вірусу (продуктивна і абортивна інфекція), по результату інфекційного процесу для клітини (літична і нелітіческая інфекція), по динаміці взаємо модействие вірусу і клітини (гостра і хронічна інфекція ).
Автономний тип вірусної інфекції клітини характерний для більшості відомих вірусів тварин і людини. При цьому типі інфекції геном вірусу реплицируется незалежно від Реплі кации клітинного генома.
Продуктивна автономна вірусна інфекція характеризується утворенням повноцінного вірусу. Це найбільш вивчена фор ма взаємодії вірусів і клітин. Основні механізми репро продукції вірусів, про яку говорилося в главі 3, досліджені і описані саме для даної форми інфекції.
Автономна абортивна вірусна інфекція виникає, якщо вза імодействіе вірусу з клітиною не закінчується утворенням повно цінних віріонів. Зазвичай це має місце при зараженні клітин дефектним вірусом; при проникненні вірусу в резистентні до нього клітини; при інфікуванні чутливих клітин повно цінних вірусом в неперміссівних умовах.
Літична автономна вірусна інфекція призводить клітку до загибелі. Вірус, що викликає загибель клітини, називається цітопато генним, а пошкодження клітини позначається терміном «цітопа, тогенний ефект». Характер цітопатологіческіх змін при вірусних інфекціях дуже різноманітний. Це може бути деге
Нерація клітин (вакуолізація цитоплазми, пикноз ядер, хромо Зімніть аберації), утворення специфічних вірусних внут ріклеточних включень, виникнення гігантських клітин типу полікаріоцітов або симпластов. Лизис клітини - заключна стадія дії цитопатогенного вірусів. Для прояви цито патогенності вірусу не потрібно відтворення генома і реа лізація всієї генетичної інформації.
Нелітіческая автономна вірусна інфекція характерна для вірусів, що містять ліпопротеїднихоболонку зовнішню оболонку і по кидають клітини брунькуванням. Ретро, \u200b\u200bортоміксо, параміксо, аренавіруси здатні викликати продуктивну нелітіческую ін фекции клітин. Однак в найбільш типових випадках нелітічес кий характер властивий абортивною інфекцій, коли відсутність про обхідних або не виявляються генетичні властивості вірусів, необ ходимо для здійснення цитопатогенного функцій.
За динамікою інфекційного процесу вірусні інфекції поділяють на гострі і хронічні. при гострої інфекції в зараженій клітині відбувається утворення віріонів, після чо го інфекційний процес припиняється. При хронічній ін фекции заражена клітина продукує віріони або вірусні компоненти до своєї природної загибелі (чи не від цієї вірус ної інфекції), при цьому дочірні клітини залишаються инфициро ванними. Продуктивна і абортивна литическая інфекція - завжди носить гострий характер. Продуктивна нелітіческая ін фекции - завжди хронічна. Абортивна нелітіческая инфек ція, як правило, має хронічний перебіг, але може проти кати і в гострій формі.
Інтегративний тип вірусної інфекції характеризується фізичним ного об'єднанням (інтеграцією) вірусного і клітинного гено мов. При цьому вірусний геном реплікується і функціонує як складова частина клітинного генома. Такий тип вірусної инфек ції обов'язковий для ретровірусів і можливий в разі папова, аде але і герпесвірусів.
Інтегративні інфекції можуть бути класифіковані за тими ж параметрами, що і автономні, - за продуктивністю, ді динаміці і результату; інтеграційні взаємовідносини вірусу і клітини також можуть бути продуктивними і абортивними. Продук, тивная інтеграційна інфекція типова для ретровірусів. Абор, тивная інтеграційна інфекція, що викликається ретровирусами, віз ника при тих же умовах, які описані вище для аборту них автономних інфекцій. Так, дефектні ретровіруси, позбавлений
Ні частини генома (деякі штами вірусу саркоми Рауса, ви руси саркоми мишей, кішок, мавп, віруси лейкозу мишей Френда і Раушера і т.п.) здатні самі по собі викликати тільки абортивні інтегративну інфекцію, яка може превра щаться в продуктивну при наявності вірусу помічника. Примі ром абортивної інтегративної інфекції, спричиненої повно цінних ретровирусом в неперміссівних клітинах, є зара ження пташиним вірусом саркоми Рауса клітин ссавців. При цьому в клітинах протікає абортивний процес інтеграції, що характеризується зниженим утворенням вірусної РНК, накопиченням неповного асортименту вірусних білків, порушен ням їх процесингу, відсутністю освіти віріонів.
Зазвичай інтегративні інфекції є нелітіческімі, як в разі ретровірусів типів В, С, D, онкогенних папова, аде але, герпесвирусов, але існують і литические інфекції, викли ваемие Цитопатогенні ретровирусом ретікулоендотеліоза птахів, або «пеня» вірусом.
Вірогенная інтеграційна інфекція - це абортивна ін фекции, яку можна перевести в продуктивну форму. Клет ки з такою формою інфекції можуть спонтанно або під впливом спеціальних (фізичних, хімічних, біологічних) воздей ствий почати продукцію інфекційного вірусу. Це явище названо вірогеніей, а клітини позначаються як вірогенние, за аналогією з Лізогенія і Лізогенія клітинами в системі фаг - бактерія.
Ще по темі Вірусна інфекція клітин:
- Вірусні інфекції та інфекції імовірно вірусної етіології
- Бактеріальні та вірусні повітряно-крапельні інфекції: грип, парагрип, аденовірусна інфекція, респіраторно-сінцітіаль-ва інфекція. Бактеріальна бронхопневмонія, лобарная пневмонія.